
An Account of Models of Molecular Circuits for Associative Learning with Reinforcement Effect and Forced Dissociation


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Associative Learning | Decision Making | Oscillator |
|---|---|---|
| Fernando et al., 2009 [53] | Nene et al., 2012 [30] | Stricker et al., 2008 [36] |
| Nesbeth et al., 2016 [18] | Filicheva et al., 2016 [31] | Tigges et al., 2009 [37] |
| Macia et al., 2017 [34] | Abrego and Zaikin, 2017 [32] | Borg et al., 2022 [35] |
2. Models
2.1. Fernando’s Model
2.2. A Model with Forced Dissociation
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benner, S.A.; Sismour, A.M. Synthetic biology. Nat. Rev. Genet. 2005, 6, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.E.; Bashor, C.J.; Collins, J.J. A brief history of synthetic biology. Nat. Rev. Microbiol. 2014, 12, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.S.; Collins, J.J. Synthetic biology: Applications come of age. Nat. Rev. Genet. 2010, 11, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Ruder, W.C.; Lu, T.; Collins, J.J. Synthetic biology moving into the clinic. Science 2011, 333, 1248–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueso, Y.F.; Lehouritis, P.; Tangney, M. In situ biomolecule production by bacteria; a synthetic biology approach to medicine. J. Control. Release 2018, 275, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.R.; Jusiak, B.; Lu, T.K. Engineering advanced cancer therapies with synthetic biology. Nat. Rev. Cancer 2019, 19, 187–195. [Google Scholar] [CrossRef]
- Goold, H.D.; Wright, P.; Hailstones, D. Emerging opportunities for synthetic biology in agriculture. Genes 2018, 9, 341. [Google Scholar] [CrossRef] [Green Version]
- Wurtzel, E.T.; Vickers, C.E.; Hanson, A.D.; Millar, A.H.; Cooper, M.; Voss-Fels, K.P.; Nikel, P.I.; Erb, T.J. Revolutionizing agriculture with synthetic biology. Nat. Plants 2019, 5, 1207–1210. [Google Scholar] [CrossRef]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome engineering: Synthetic biology of plant-associated microbiomes in sustainable agriculture. Trends Biotechnol. 2021, 39, 244–261. [Google Scholar] [CrossRef]
- Bueso, Y.F.; Tangney, M. Synthetic biology in the driving seat of the bioeconomy. Trends Biotechnol. 2017, 35, 373–378. [Google Scholar] [CrossRef]
- Anderson, J.C.; Clarke, E.J.; Arkin, A.P.; Voigt, C.A. Environmentally controlled invasion of cancer cells by engineered bacteria. J. Mol. Biol. 2006, 355, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qi, L.S. A CRISPR–dCas toolbox for genetic engineering and synthetic biology. J. Mol. Biol. 2019, 431, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Lee, J.; Na, D. Recent advances in genetic engineering tools based on synthetic biology. J. Microbiol. 2020, 58, 1–10. [Google Scholar] [CrossRef]
- Jensen, M.K.; Keasling, J.D. Recent applications of synthetic biology tools for yeast metabolic engineering. FEMS Yeast Res. 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.A.; Ellington, A.D. Synthetic DNA synthesis and assembly: Putting the synthetic in synthetic biology. Cold Spring Harb. Perspect. Biol. 2017, 9, a023812. [Google Scholar] [CrossRef] [Green Version]
- Borg, Y.; Grigonyte, A.M.; Boeing, P.; Wolfenden, B.; Smith, P.; Beaufoy, W.; Rose, S.; Ratisai, T.; Zaikin, A.; Nesbeth, D.N. Open source approaches to establishing Roseobacter clade bacteria as synthetic biology chassis for biogeoengineering. PeerJ 2016, 4, e2031. [Google Scholar] [CrossRef] [Green Version]
- Nesbeth, D.N.; Zaikin, A.; Saka, Y.; Romano, M.C.; Giuraniuc, C.V.; Kanakov, O.; Laptyeva, T. Synthetic biology routes to bio-artificial intelligence. Essays Biochem. 2016, 60, 381–391. [Google Scholar]
- Bianchini, F. Artificial intelligence and synthetic biology: A tri-temporal contribution. Biosystems 2016, 148, 32–39. [Google Scholar] [CrossRef]
- Abrego, L.; Zaikin, A. Integrated information as a measure of cognitive processes in coupled genetic repressilators. Entropy 2019, 21, 382. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lee, J.; Land, M.A.; Lai, S.; Igoshin, O.A.; St-Pierre, F. A synthetic circuit for buffering gene dosage variation between individual mammalian cells. Nat. Commun. 2021, 12, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; White, K.S.; Winfree, E. Construction of an in vitro bistable circuit from synthetic transcriptional switches. Mol. Syst. Biol. 2006, 2, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacchus, W.; Aubel, D.; Fussenegger, M. Biomedically relevant circuit-design strategies in mammalian synthetic biology. Mol. Syst. Biol. 2013, 9, 691. [Google Scholar] [CrossRef] [PubMed]
- Brophy, J.A.; Voigt, C.A. Principles of genetic circuit design. Nat. Methods 2014, 11, 508–520. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Kaern, M.; Araki, M.; Chung, K.; Gardner, T.S.; Cantor, C.R.; Collins, J.J. Programmable cells: Interfacing natural and engineered gene networks. Proc. Natl. Acad. Sci. USA 2004, 101, 8414–8419. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Fussenegger, M. Designing cell function: Assembly of synthetic gene circuits for cell biology applications. Nat. Rev. Mol. Cell Biol. 2018, 19, 507–525. [Google Scholar] [CrossRef]
- Xia, P.F.; Ling, H.; Foo, J.L.; Chang, M.W. Synthetic genetic circuits for programmable biological functionalities. Biotechnol. Adv. 2019, 37, 107393. [Google Scholar] [CrossRef]
- Tolle, F.; Stücheli, P.; Fussenegger, M. Genetic circuitry for personalized human cell therapy. Curr. Opin. Biotechnol. 2019, 59, 31–38. [Google Scholar] [CrossRef]
- Prochazka, L.; Benenson, Y.; Zandstra, P.W. Synthetic gene circuits and cellular decision-making in human pluripotent stem cells. Curr. Opin. Syst. Biol. 2017, 5, 93–103. [Google Scholar] [CrossRef]
- Nene, N.R.; Garca-Ojalvo, J.; Zaikin, A. Speed-dependent cellular decision making in nonequilibrium genetic circuits. PLoS ONE 2012, 7, e32779. [Google Scholar] [CrossRef] [Green Version]
- Filicheva, S.; Zaikin, A.; Kanakov, O. Dynamical decision making in a genetic perceptron. Phys. D Nonlinear Phenom. 2016, 318, 112–115. [Google Scholar] [CrossRef]
- Abrego, L.; Zaikin, A. Decision making in an intracellular genetic classifier. Math. Model. Nat. Phenom. 2017, 12, 30–42. [Google Scholar] [CrossRef]
- Kanakov, O.; Kotelnikov, R.; Alsaedi, A.; Tsimring, L.; Huerta, R.; Zaikin, A.; Ivanchenko, M. Multi-input distributed classifiers for synthetic genetic circuits. PLoS ONE 2015, 10, e0125144. [Google Scholar] [CrossRef] [Green Version]
- Macia, J.; Vidiella, B.; Solé, R.V. Synthetic associative learning in engineered multicellular consortia. J. R. Soc. Interface 2017, 14, 20170158. [Google Scholar] [CrossRef] [PubMed]
- Borg, Y.; Alsford, S.; Pavlika, V.; Zaikin, A.; Nesbeth, D.N. Synthetic biology tools for engineering Goodwin oscillation in Trypanosoma brucei brucei. Heliyon 2022, 8, e08891. [Google Scholar] [CrossRef]
- Stricker, J.; Cookson, S.; Bennett, M.R.; Mather, W.H.; Tsimring, L.S.; Hasty, J. A fast, robust and tunable synthetic gene oscillator. Nature 2008, 456, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Tigges, M.; Marquez-Lago, T.T.; Stelling, J.; Fussenegger, M. A tunable synthetic mammalian oscillator. Nature 2009, 457, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Ullner, E.; Koseska, A.; Zaikin, A.; Volkov, E.; Kurths, J.; Garcia-Ojalvo, J. Dynamics of Multicellular Synthetic Gene Networks. In Handbook on Biological Networks; World Scientific: Singapore, 2009. [Google Scholar]
- Ullner, E.; Ares, S.; Morelli, L.G.; Oates, A.C.; Jülicher, F.; Nicola, E.; Heussen, R.; Whitmore, D.; Blyuss, K.; Fryett, M.; et al. Noise and oscillations in biological systems: Multidisciplinary approach between experimental biology, theoretical modelling and synthetic biology. Int. J. Mod. Phys. B 2012, 26, 1246009. [Google Scholar] [CrossRef] [Green Version]
- Ullner, E.; Zaikin, A.; Volkov, E.I.; García-Ojalvo, J. Multistability and clustering in a population of synthetic genetic oscillators via phase-repulsive cell-to-cell communication. Phys. Rev. Lett. 2007, 99, 148103. [Google Scholar] [CrossRef] [Green Version]
- Anesiadis, N.; Kobayashi, H.; Cluett, W.R.; Mahadevan, R. Analysis and design of a genetic circuit for dynamic metabolic engineering. ACS Synth. Biol. 2013, 2, 442–452. [Google Scholar] [CrossRef]
- Shanks, D.R. The Psychology of Associative Learning; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Wasserman, E.A.; Miller, R.R. What’s elementary about associative learning? Annu. Rev. Psychol. 1997, 48, 573–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, J.M.; Bouton, M.E. Theories of associative learning in animals. Annu. Rev. Psychol. 2001, 52, 111–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, T.E.; Hunt, L.T.; Woolrich, M.W.; Rushworth, M.F. Associative learning of social value. Nature 2008, 456, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; De Houwer, J.; Lovibond, P.F. The propositional nature of human associative learning. Behav. Brain Sci. 2009, 32, 183–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, P.I. Conditioned reflexes: An investigation of the physiological activity of the cerebral cortex. Ann. Neurosci. 2010, 17, 136. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, M.; Soni, R.; Patelczyk, T.; Ignatov, M.; Bartsch, T.; Meuffels, P.; Kohlstedt, H. An electronic version of Pavlov’s dog. Adv. Funct. Mater. 2012, 22, 2744–2749. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Poulos, A.M. The neuroscience of mammalian associative learning. Annu. Rev. Psychol. 2005, 56, 207–234. [Google Scholar] [CrossRef] [Green Version]
- Uwano, T.; Nishijo, H.; Ono, T.; Tamura, R. Neuronal responsiveness to various sensory stimuli, and associative learning in the rat amygdala. Neuroscience 1995, 68, 339–361. [Google Scholar] [CrossRef]
- Faber, T.; Joerges, J.; Menzel, R. Associative learning modifies neural representations of odors in the insect brain. Nat. Neurosci. 1999, 2, 74–78. [Google Scholar] [CrossRef]
- Johansen, J.P.; Diaz-Mataix, L.; Hamanaka, H.; Ozawa, T.; Ycu, E.; Koivumaa, J.; Kumar, A.; Hou, M.; Deisseroth, K.; Boyden, E.S.; et al. Hebbian and neuromodulatory mechanisms interact to trigger associative memory formation. Proc. Natl. Acad. Sci. USA 2014, 111, E5584–E5592. [Google Scholar] [CrossRef] [Green Version]
- Fern, C.T.; Liekens, A.M.; Bingle, L.E.; Beck, C.; Lenser, T.; Stekel, D.J.; Rowe, J.E. Molecular circuits for associative learning in single-celled organisms. J. R. Soc. Interface 2009, 6, 463–469. [Google Scholar]
- Bray, D. Molecular networks: The top-down view. Science 2003, 301, 1864–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, E.B.; Becker, I.; Shapira, Y.; Levine, H. Bacterial linguistic communication and social intelligence. TRENDS Microbiol. 2004, 12, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.K.; Saez-Rodriguez, J.; Sorger, P.K.; Lauffenburger, D.A. Logic-based models for the analysis of cell signaling networks. Biochemistry 2010, 49, 3216–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, T.S.; Cantor, C.R.; Collins, J.J. Construction of a genetic toggle switch in Escherichia coli. Nature 2000, 403, 339–342. [Google Scholar] [CrossRef]
- Siuti, P.; Yazbek, J.; Lu, T.K. Synthetic circuits integrating logic and memory in living cells. Nat. Biotechnol. 2013, 31, 448–452. [Google Scholar] [CrossRef]
- Padirac, A.; Fujii, T.; Rondelez, Y. Bottom-up construction of in vitro switchable memories. Proc. Natl. Acad. Sci. USA 2012, 109, E3212–E3220. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Kitney, R.I.; Joly, N.; Buck, M. Engineering modular and orthogonal genetic logic gates for robust digital-like synthetic biology. Nat. Commun. 2011, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.V. The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. J. Physiol. 1910, 40, 4–7. [Google Scholar]
- O’Brien, E.L.; Van Itallie, E.; Bennett, M.R. Modeling synthetic gene oscillators. Math. Biosci. 2012, 236, 1–5. [Google Scholar] [CrossRef]
- Zorzan, I.; López, A.R.; Malyshava, A.; Ellis, T.; Barberis, M. Synthetic designs regulating cellular transitions: Fine-tuning of switches and oscillators. Curr. Opin. Syst. Biol. 2021, 25, 11–26. [Google Scholar] [CrossRef]
- Fitzgerald, M.; Gibbs, C.; Shimpi, A.A.; Deans, T.L. Adoption of the Q transcriptional system for regulating gene expression in stem cells. ACS Synth. Biol. 2017, 6, 2014–2020. [Google Scholar] [CrossRef] [PubMed]
- Linsker, R. Local synaptic learning rules suffice to maximize mutual information in a linear network. Neural Comput. 1992, 4, 691–702. [Google Scholar] [CrossRef]
- Higham, N.J. Accuracy and Stability of Numerical Algorithms; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 2002. [Google Scholar]
- Burden, R.L.; Faires, J.D.; Burden, A.M. Numerical Analysis; Cengage Learning: Boston, MA, USA, 2015. [Google Scholar]
- Dorm, J.R.; Prince, P.J. A family of embedded Runge-Kutta formulae. J. Comput. Appl. Math. 1980, 6, 19–26. [Google Scholar]
- Ye, H.; Baba, M.D.E.; Peng, R.W.; Fussenegger, M. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science 2011, 332, 1565–1568. [Google Scholar] [CrossRef]
- Kalos, M.; June, C.H. Adoptive T cell transfer for cancer immunotherapy in the era of synthetic biology. Immunity 2013, 39, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Krivonosov, M.; Nazarenko, T.; Bacalini, M.G.; Zaikin, A.; Ivanchenko, M.; Franceschi, C. Network markers of DNA methylation in neurodegenerative diseases. In Proceedings of the 2020 4th Scientific School on Dynamics of Complex Networks and their Application in Intellectual Robotics (DCNAIR), Innopolis, Russia, 7–9 September 2020; pp. 138–139. [Google Scholar]
- Krivonosov, M.; Nazarenko, T.; Bacalini, M.G.; Franceschi, C.; Zaikin, A.; Ivanchenko, M. DNA methylation changes with age as a complex system: A parenclitic network approach to a family-based cohort of patients with Down Syndrome. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Rössger, K.; Charpin-El Hamri, G.; Fussenegger, M. Reward-based hypertension control by a synthetic brain–dopamine interface. Proc. Natl. Acad. Sci. USA 2013, 110, 18150–18155. [Google Scholar] [CrossRef] [Green Version]


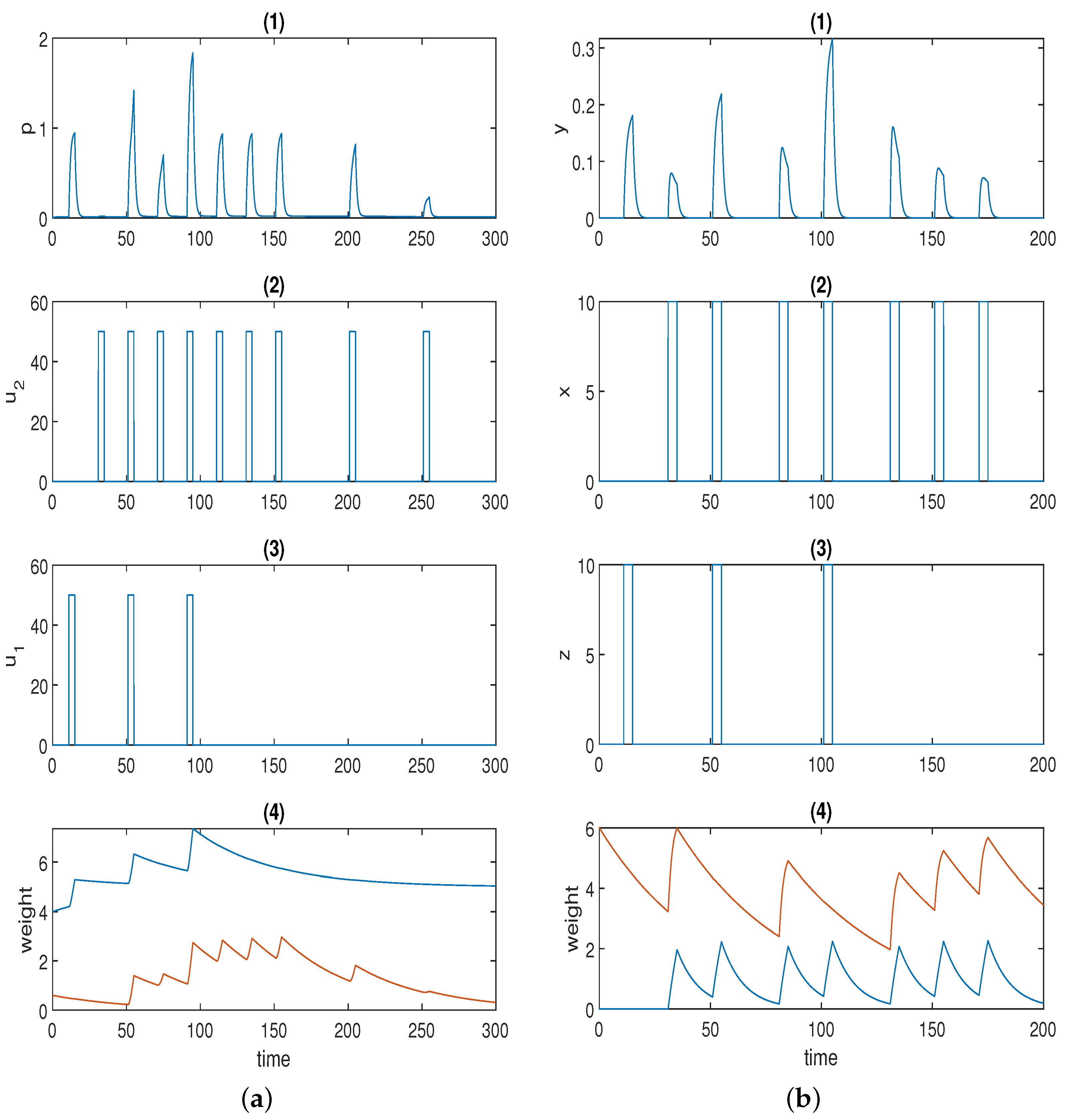
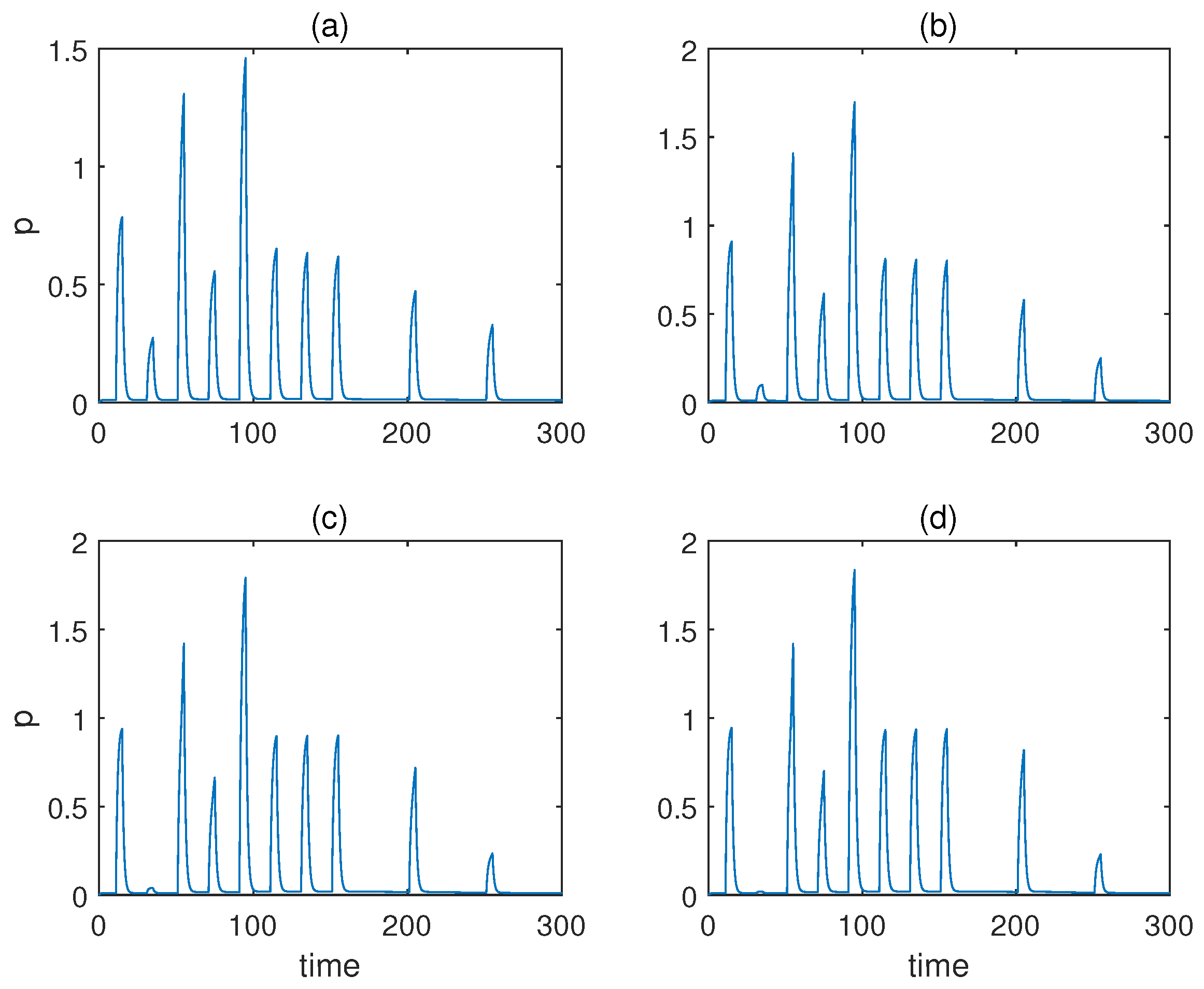





| Parameter | Value |
|---|---|
| 1 | |
| 0.8 | |
| 0.02 | |
| 0.1 | |
| S | 10 |
| Parameter | Value |
|---|---|
| 2 | |
| 4 | |
| 4 | |
| 0.6 | |
| 1.5 | |
| 0.1 | |
| 0.02 |
| Parameter | Value |
|---|---|
| 4 | |
| 1 | |
| 0.6 | |
| 1 | |
| 0.1 | |
| 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Fattah, A.; Timashev, P.; Zaikin, A. An Account of Models of Molecular Circuits for Associative Learning with Reinforcement Effect and Forced Dissociation. Sensors 2022, 22, 5907. https://doi.org/10.3390/s22155907
Li Z, Fattah A, Timashev P, Zaikin A. An Account of Models of Molecular Circuits for Associative Learning with Reinforcement Effect and Forced Dissociation. Sensors. 2022; 22(15):5907. https://doi.org/10.3390/s22155907
Chicago/Turabian StyleLi, Zonglun, Alya Fattah, Peter Timashev, and Alexey Zaikin. 2022. "An Account of Models of Molecular Circuits for Associative Learning with Reinforcement Effect and Forced Dissociation" Sensors 22, no. 15: 5907. https://doi.org/10.3390/s22155907
APA StyleLi, Z., Fattah, A., Timashev, P., & Zaikin, A. (2022). An Account of Models of Molecular Circuits for Associative Learning with Reinforcement Effect and Forced Dissociation. Sensors, 22(15), 5907. https://doi.org/10.3390/s22155907