
Cortical Correlates of Increased Postural Task Difficulty in Young Adults: A Combined Pupillometry and EEG Study

 ,
, 
Abstract
:Highlights
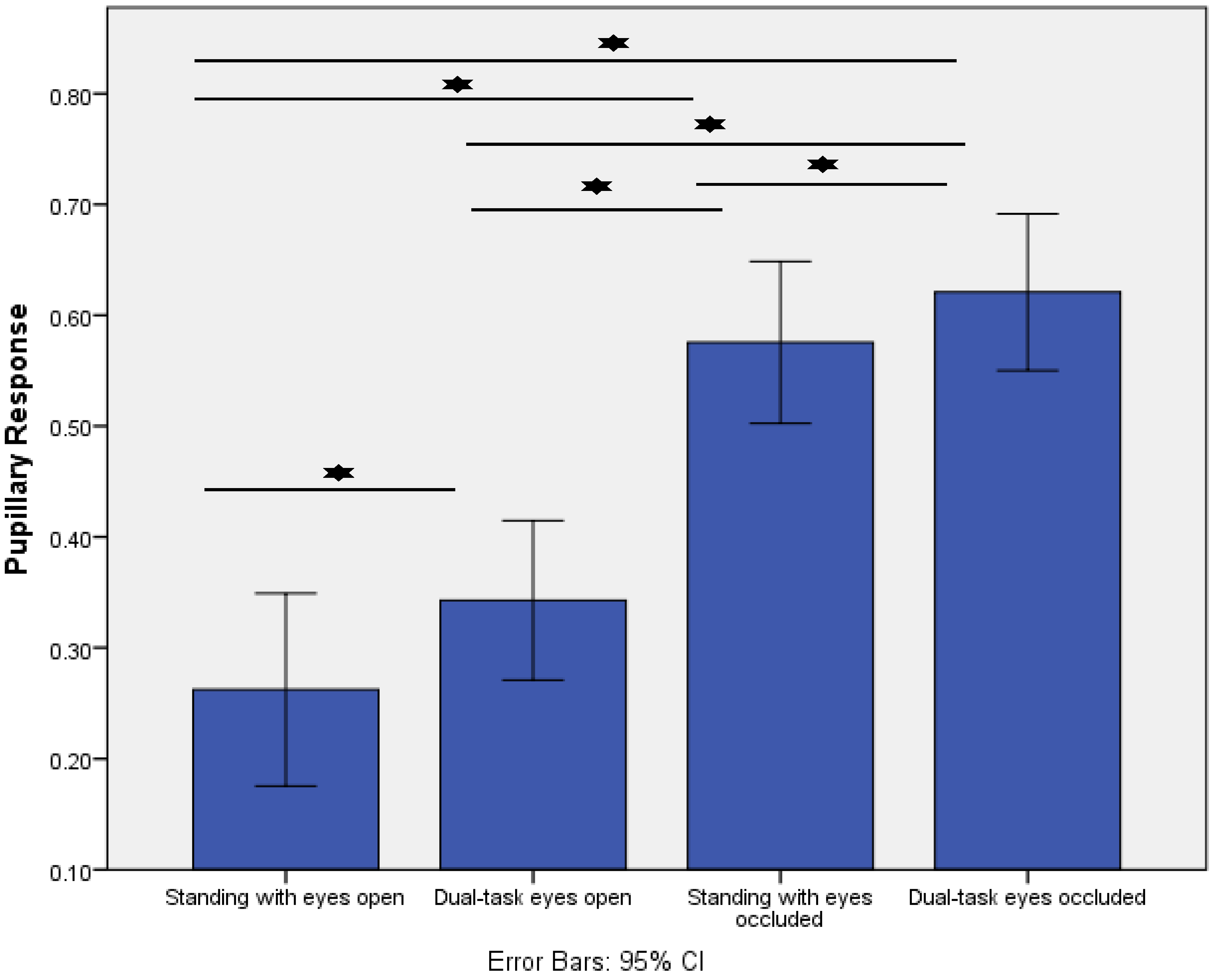
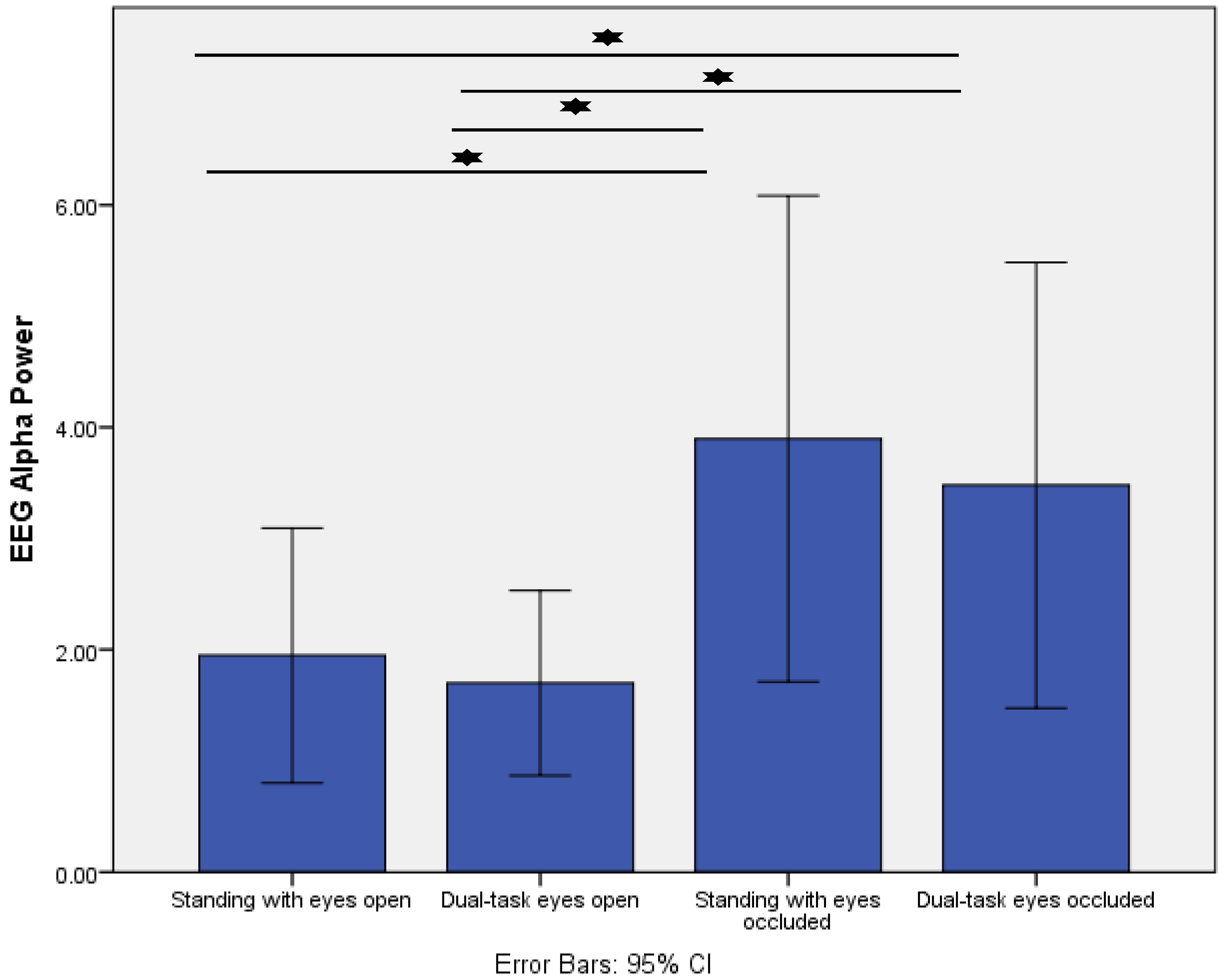
- Increased postural task difficulty was associated with greater responses for pupillary response, posterior alpha power, and fronto-central theta/beta power ratio.
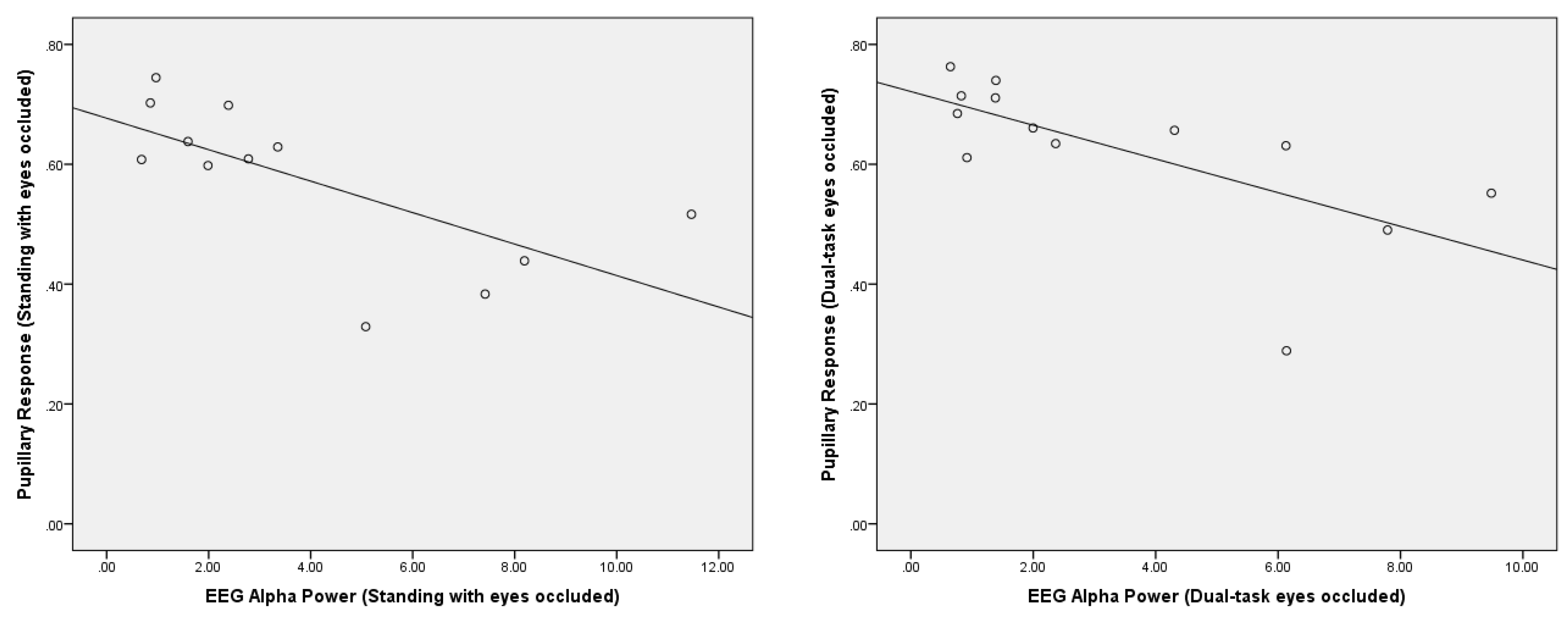
- Greater pupillary response correlated with lower posterior EEG alpha power during eyes occluded standing with and without dual-task.
- Lastly, a greater pupillary response was associated with lower CoP displacement in the anterior-posterior direction during dual-task eyes occluded.
Abstract
1. Introduction
2. Methods
2.1. Participants
2.2. Protocol
- Standing with eyes open (control condition): Participants stood on a force plate and maintained an upright standing posture for 320 s.
- Dual-task eyes open: Participants stood on the force plate for 320 s while concurrently completing an auditory 2-back test by using a thumb-press button.
- Standing with eyes occluded: Participants stood on a force plate for 320 s with a sleep mask covering the eye tracker. We asked our participants to keep their eyes open to record the pupillary response.
- Dual-task eyes occluded: Participants stood on a force plate for 320 s while completing the 2-back test with their eyes occluded.
2.3. Outcome Variables
2.3.1. Pupillary Response
2.3.2. EEG
2.3.3. COP
3. Statistical Analysis
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobs, J.V.; Horak, F.B. Cortical control of postural responses. J. Neural Transm. 2007, 114, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, B.E.; McIlroy, W.E. Cognitive demands and cortical control of human balance-recovery reactions. J. Neural Transm. 2007, 114, 1279–1296. [Google Scholar] [CrossRef] [Green Version]
- Kahya, M.; Wood, T.A.; Sosnoff, J.; Devos, H. Increased postural demand is associated with greater cognitive workload in healthy young adults: A pupillometry study. Front. Hum. Neurosci. 2018, 12, 288. [Google Scholar] [CrossRef] [Green Version]
- Kahya, M.; Lyons, K.E.; Pahwa, R.; Akinwuntan, A.E.; He, J.; Devos, H. Reliability and validity of pupillary response during dual-task balance in Parkinson Disease. Arch. Phys. Med. Rehabil. 2020, 102, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Kahya, M.; Lyons, K.E.; Pahwa, R.; Akinwuntan, A.E.; He, J.; Devos, H. Pupillary Response to Postural Demand in Parkinson’s Disease. Front. Bioeng. Biotechnol. 2021, 9, 237. [Google Scholar] [CrossRef]
- Bopp, K.L.; Verhaeghen, P. Aging and n-back performance: A meta-analysis. J. Gerontol. Ser. B 2020, 75, 229–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychol. Bull. 1982, 91, 276. [Google Scholar] [CrossRef]
- Sara, S.J. The locus coeruleus and noradrenergic modulation of cognition. Nat. Rev. Neurosci. 2009, 10, 211–223. [Google Scholar] [CrossRef]
- Beatty, J.; Lucero-Wagoner, B. The pupillary system. Handb. Psychophysiol. 2000, 2, 142–162. [Google Scholar]
- Costa, V.D.; Rudebeck, P.H. More than meets the eye: The relationship between pupil size and locus coeruleus activity. Neuron 2016, 89, 8–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiew, K.S.; Braver, T.S. Temporal dynamics of motivation-cognitive control interactions revealed by high-resolution pupillometry. Front. Psychol. 2013, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Varghese, J.P.; Staines, W.R.; McIlroy, W.E. Activity in functional cortical networks temporally associated with postural instability. Neuroscience 2019, 401, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Gramann, K.; Gwin, J.T.; Ferris, D.; Oie, K.; Jung, T.-P.; Lin, C.-T.; Liao, L.-D.; Makeig, S. Cognition in action: Imaging brain/body dynamics in mobile humans. Rev. Neurosci. 2011, 22, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Urigüen, J.A.; Garcia-Zapirain, B. EEG artifact removal—state-of-the-art and guidelines. J. Neural Eng. 2015, 12, 031001. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S.; Bell, A.J.; Jung, T.-P.; Sejnowski, T.J. Independent component analysis of electroencephalographic data. Adv. Neural Inf. Process. Syst. 1996, 145–151. [Google Scholar] [CrossRef]
- Light, G.A.; Williams, L.E.; Minow, F.; Sprock, J.; Rissling, A.; Sharp, R.; Swerdlow, N.R.; Braff, D.L. Electroencephalography (EEG) and event-related potentials (ERPs) with human participants. Curr. Protoc. Neurosci. 2010, 52, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Hülsdünker, T.; Mierau, A.; Strüder, H.K. Higher balance task demands are associated with an increase in individual alpha peak frequency. Front. Hum. Neurosci. 2016, 9, 695. [Google Scholar] [CrossRef] [Green Version]
- Del Percio, C.; Brancucci, A.; Bergami, F.; Marzano, N.; Fiore, A.; Di Ciolo, E.; Aschieri, P.; Lino, A.; Vecchio, F.; Iacoboni, M.; et al. Cortical alpha rhythms are correlated with body sway during quiet open-eyes standing in athletes: A high-resolution EEG study. Neuroimage 2007, 36, 822–829. [Google Scholar] [CrossRef]
- Ray, W.J.; Cole, H.W. EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar] [CrossRef]
- Klimesch, W. Alpha-band oscillations, attention, and controlled access to stored information. Trends Cogn. Sci. 2012, 16, 606–617. [Google Scholar] [CrossRef] [Green Version]
- Compton, R.J.; Gearinger, D.; Wild, H.; Rette, D.; Heaton, E.C.; Histon, S.; Thiel, P.; Jaskir, M. Simultaneous EEG and pupillary evidence for post-error arousal during a speeded performance task. Eur. J. Neurosci. 2020, 53, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Ala, T.S.; Graversen, C.; Wendt, D.; Alickovic, E.; Whitmer, W.M.; Lunner, T. An exploratory Study of EEG Alpha Oscillation and Pupil Dilation in Hearing-Aid Users During Effortful listening to Continuous Speech. PLoS ONE 2020, 15, e0235782. [Google Scholar]
- LoTemplio, S.; Silcox, J.; Federmeier, K.D.; Payne, B.R. Inter-and intra-individual coupling between pupillary, electrophysiological, and behavioral responses in a visual oddball task. Psychophysiology 2020, 58, e13758. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; Karamacoska, D.; Johnstone, S.J. The EEG theta/beta ratio: A marker of arousal or cognitive processing capacity? Appl. Psychophysiol. Biofeedback 2019, 44, 123–129. [Google Scholar] [CrossRef] [PubMed]
- van Dinteren, R.; Arns, M.; Jongsma, M.L.; Kessels, R.P. P300 development across the lifespan: A systematic review and meta-analysis. PLoS ONE 2014, 9, e87347. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.P. Identifying cognitive state from eye metrics. Aviat. Space Environ. Med. 2007, 78, B165–B175. [Google Scholar]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Hinkle, D.E.; Wiersma, W.; Jurs, S.G. Applied Statistics for the Behavioral Sciences; Houghton Mifflin: Boston, MS, USA, 1988. [Google Scholar]
- Babiloni, C.; Del Percio, C.; Arendt-Nielsen, L.; Soricelli, A.; Romani, G.L.; Rossini, P.M.; Capotosto, P. Cortical EEG alpha rhythms reflect task-specific somatosensory and motor interactions in humans. Clin. Neurophysiol. 2014, 125, 1936–1945. [Google Scholar] [CrossRef]
- Edwards, A.E.; Guven, O.; Furman, M.D.; Arshad, Q.; Bronstein, A.M. Electroencephalographic correlates of continuous postural tasks of increasing difficulty. Neuroscience 2018, 395, 35–48. [Google Scholar] [CrossRef]
- Başar, E. A review of alpha activity in integrative brain function: Fundamental physiology, sensory coding, cognition and pathology. Int. J. Psychophysiol. 2012, 86, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Gentili, R.J.; Jaquess, K.J.; Shuggi, I.M.; Shaw, E.P.; Oh, H.; Lo, L.-C.; Tan, Y.Y.; Domingues, C.A.; Blanco, J.A.; Rietschel, J.C. Combined assessment of attentional reserve and cognitive-motor effort under various levels of challenge with a dry EEG system. Psychophysiology 2018, 55, e13059. [Google Scholar] [CrossRef] [PubMed]
- Shaw, E.P.; Rietschel, J.C.; Hendershot, B.D.; Pruziner, A.L.; Miller, M.W.; Hatfield, B.D.; Gentili, R.J. Measurement of attentional reserve and mental effort for cognitive workload assessment under various task demands during dual-task walking. Biol. Psychol. 2018, 134, 39–51. [Google Scholar] [CrossRef]
- Peterson, S.M.; Ferris, D.P. Differentiation in theta and beta electrocortical activity between visual and physical perturbations to walking and standing balance. Eneuro 2018, 5, ENEURO.0207-18.2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenny, R.P.W.; Eaves, D.L.; Martin, D.; Behmer, L.P.; Dixon, J. The Effects of Textured Insoles on Cortical Activity and Quiet Bipedal Standing With and Without Vision: An EEG Study. J. Mot. Behav. 2020, 52, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.C.; O’Keeffe, C.; O’Rourke, E.; Feerick, N.; Reilly, R.B. Cortical Theta Activity and Postural Control in Non-Visual and High Cognitive Load Tasks: Impact for Clinical Studies. In Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Berlin, Germany, 23–27 July 2019. [Google Scholar]
- Hülsdünker, T.; Mierau, A.; Neeb, C.; Kleinöder, H.; Strüder, H. Cortical processes associated with continuous balance control as revealed by EEG spectral power. Neurosci. Lett. 2015, 592, 1–5. [Google Scholar] [CrossRef]
- Slobounov, S.M.; Teel, E.; Newell, K.M. Modulation of cortical activity in response to visually induced postural perturbation: Combined VR and EEG study. Neurosci. Lett. 2013, 547, 6–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.; Solis-Escalante, T.; Scherer, R.; Neuper, C.; Müller-Putz, G. It’s how you get there: Walking down a virtual alley activates premotor and parietal areas. Front. Hum. Neurosci. 2014, 8, 93. [Google Scholar] [CrossRef]
- Sipp, A.R.; Gwin, J.T.; Makeig, S.; Ferris, D. Loss of balance during balance beam walking elicits a multifocal theta band electrocortical response. J. Neurophysiol. 2013, 110, 2050–2060. [Google Scholar] [CrossRef] [Green Version]
- Haley, S.M.; Fragala-Pinkham, M.A. Interpreting change scores of tests and measures used in physical therapy. Phys. Ther. 2006, 86, 735–743. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Healthy Young Adults (n = 15) |
|---|---|
| Age (years) | 25.6 ± 2.9 |
| Education (years) | 18.6 ± 1.6 |
| Sex (female/male) | 5/10 |
| Montreal Cognitive Assessment, out of 30 | 28.8 ± 1.5 |
| Handedness (right/left) | 15/0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahya, M.; Liao, K.; Gustafson, K.M.; Akinwuntan, A.E.; Manor, B.; Devos, H. Cortical Correlates of Increased Postural Task Difficulty in Young Adults: A Combined Pupillometry and EEG Study. Sensors 2022, 22, 5594. https://doi.org/10.3390/s22155594
Kahya M, Liao K, Gustafson KM, Akinwuntan AE, Manor B, Devos H. Cortical Correlates of Increased Postural Task Difficulty in Young Adults: A Combined Pupillometry and EEG Study. Sensors. 2022; 22(15):5594. https://doi.org/10.3390/s22155594
Chicago/Turabian StyleKahya, Melike, Ke Liao, Kathleen M. Gustafson, Abiodun E. Akinwuntan, Brad Manor, and Hannes Devos. 2022. "Cortical Correlates of Increased Postural Task Difficulty in Young Adults: A Combined Pupillometry and EEG Study" Sensors 22, no. 15: 5594. https://doi.org/10.3390/s22155594
APA StyleKahya, M., Liao, K., Gustafson, K. M., Akinwuntan, A. E., Manor, B., & Devos, H. (2022). Cortical Correlates of Increased Postural Task Difficulty in Young Adults: A Combined Pupillometry and EEG Study. Sensors, 22(15), 5594. https://doi.org/10.3390/s22155594