Acoustic Sensor-Based Soundscape Analysis and Acoustic Assessment of Bird Species Richness in Shennongjia National Park, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Data Collection
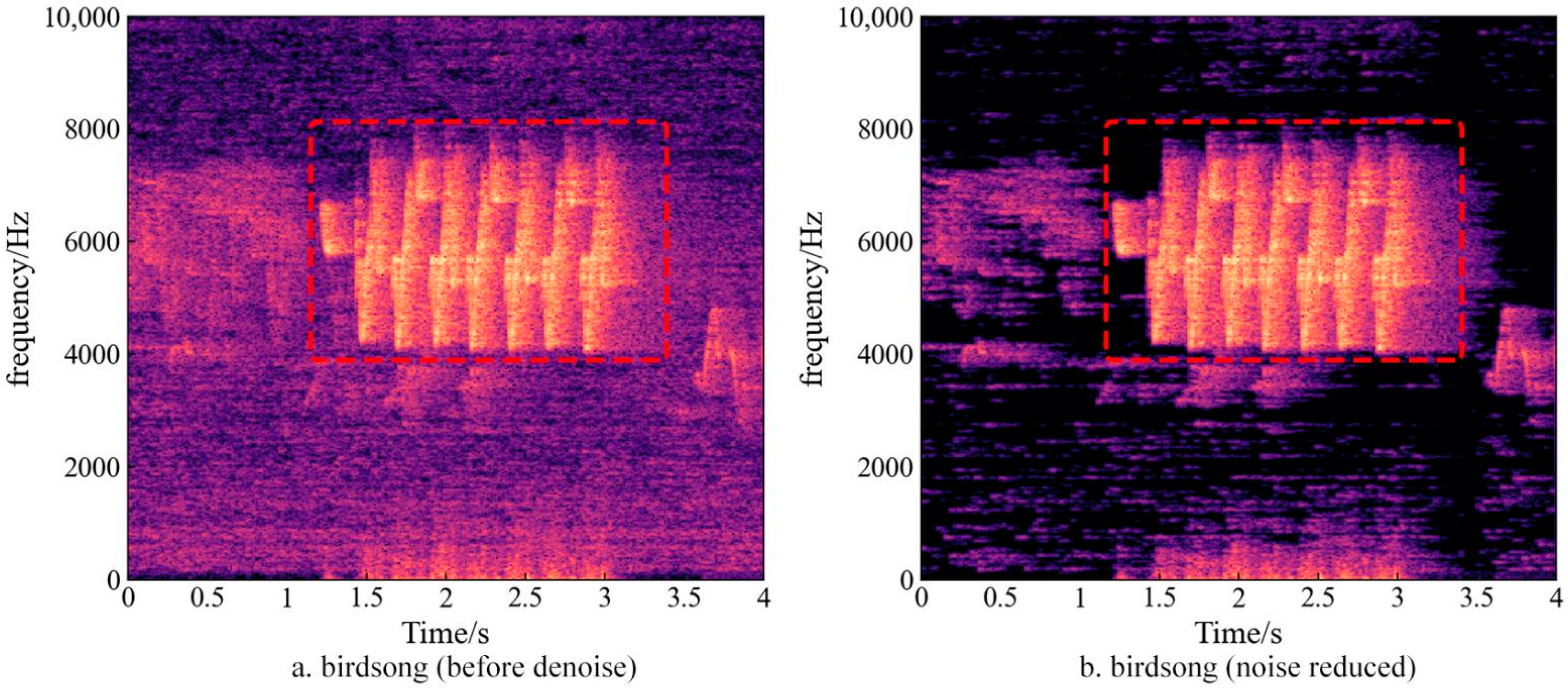
2.3. Acoustic Data Pre-Processing
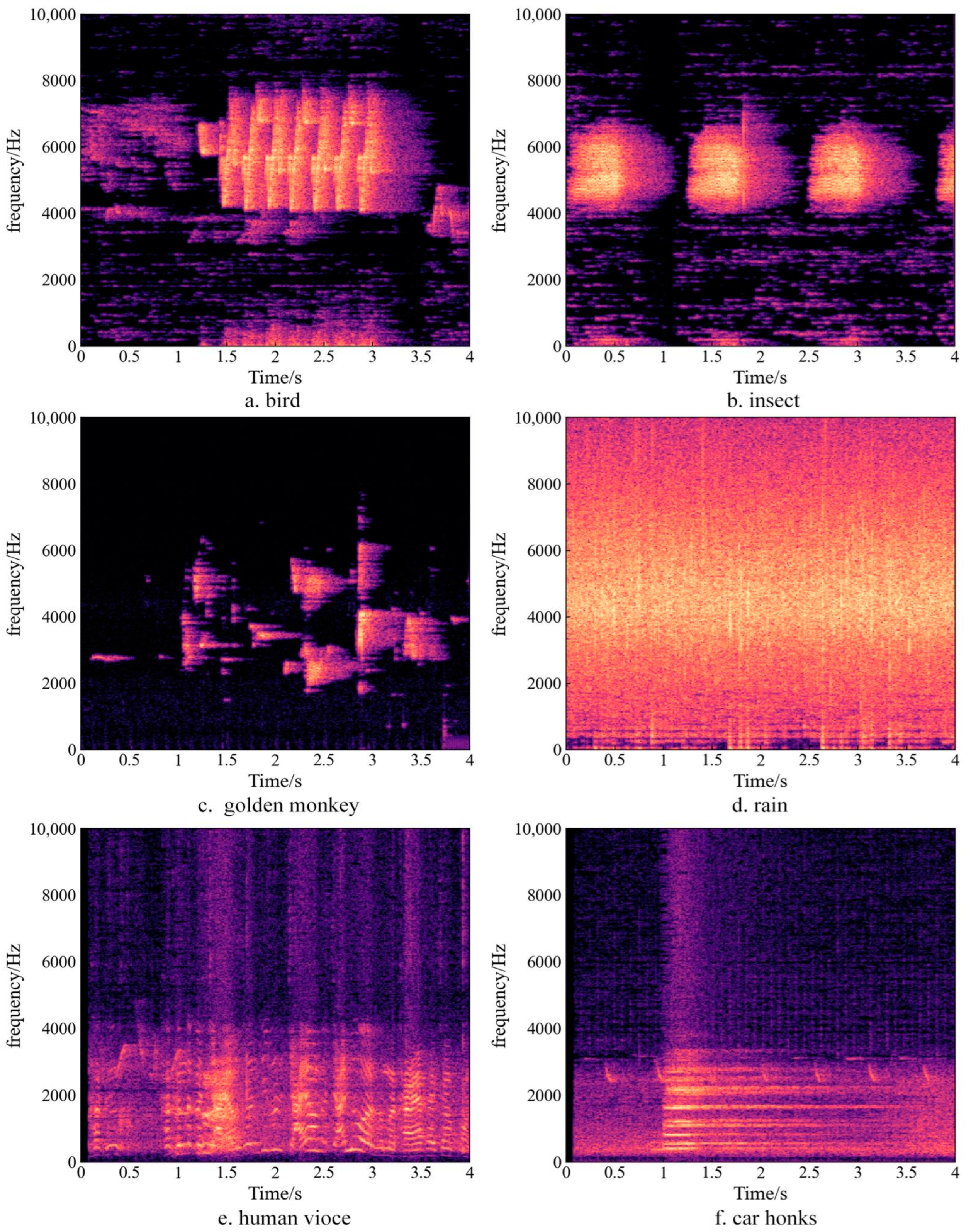
2.4. Soundscape Analysis Method
2.5. Acoustic Index Calculation
2.6. Correlation Analysis between Acoustic Index and Bird Species Richness
3. Results and Analysis
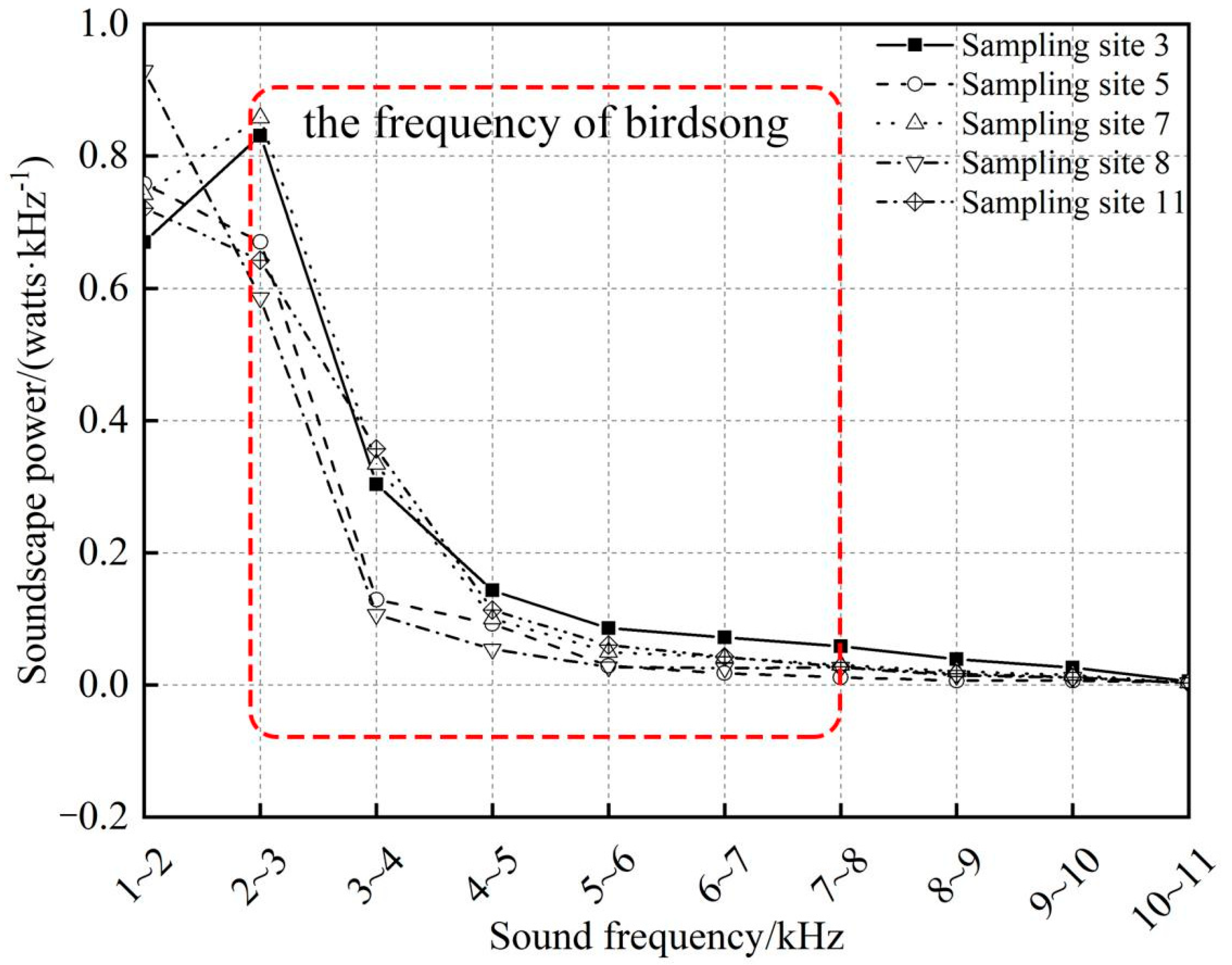
3.1. Frequency Distribution of Sound from Different Sources in the Soundscape
3.2. Correlation between Acoustic Index and Bird Species Richness
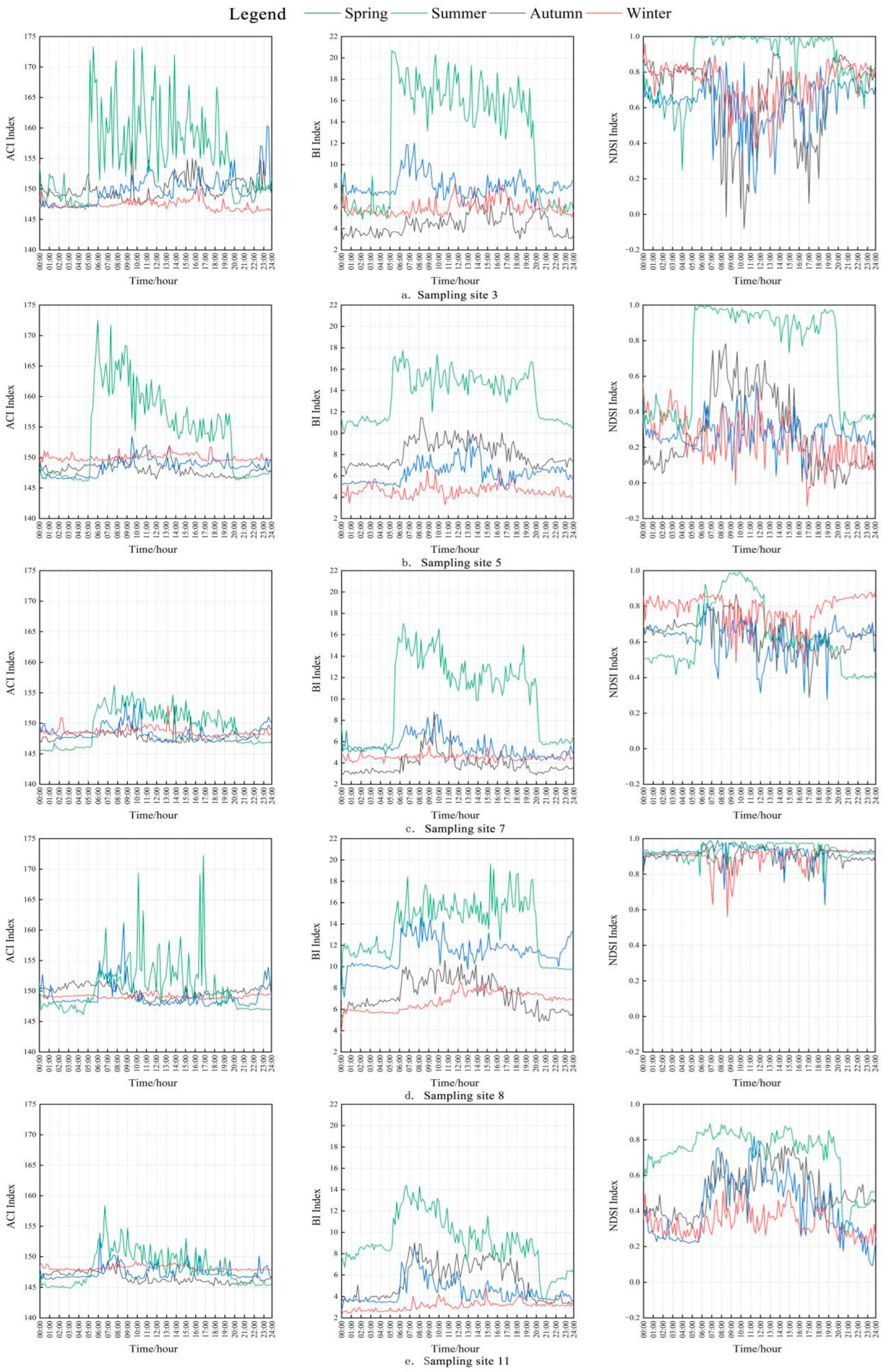
3.3. Diurnal Dynamic Characteristics of Acoustic Indices in Different Seasons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibb, R.; Browning, E.; Glover-Kapfer, P.; Jones, K.E. Emerging opportunities and challenges for passive acoustics in ecological assessment and monitoring. Methods Ecol. Evol. 2019, 10, 169–185. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.W.; Nie, Y.G.; Miao, H.X.; Lu, H.; Hu, Y.B. Advancements of the researches on biodiversity loss mechanisms. Chin. Sci. Bull. 2014, 59, 430–437. [Google Scholar] [CrossRef]
- Marques, T.A.; Thomas, L.; Martin, S.W.; Mellinger, D.K.; Ward, J.A.; Moretti, D.J.; Harris, D.; Tyack, P.L. Estimating animal population density using passive acoustics. Biol. Rev. 2013, 88, 287–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burivalova, Z.; Game, E.T.; Butler, R.A. The sound of a tropical forest. Science 2019, 363, 28–29. [Google Scholar] [CrossRef]
- Merchant, N.D.; Fristrup, K.M.; Johnson, M.P.; Tyack, P.L.; Witt, M.J.; Blondel, P.; Parks, S.E. Measuring acoustic habitats. Methods Ecol. Evol. 2015, 6, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Cato, D.; McCauley, R.; Rogers, T.; Noad, M. Passive acoustics for monitoring marine animals-progress and challenges. In Proceedings of the ACOUSTICS, Christchurch, New Zealand, 20–22 November 2006; pp. 453–460. [Google Scholar]
- Wimmer, J.; Towsey, M.; Planitz, B.; Williamson, I.; Roe, P. Analysing environmental acoustic data through collaboration and automation. Future Gener. Comput. Syst. 2013, 29, 560–568. [Google Scholar] [CrossRef] [Green Version]
- Sousa-Lima, R.S.; Norris, T.F.; Oswald, J.N.; Fernandes, D.P. A review and inventory of fixed autonomous recorders for passive acoustic monitoring of marine mammals. Aquat. Mamm. 2013, 39, 23–53. [Google Scholar] [CrossRef]
- Digby, A.; Towsey, M.; Bell, B.D.; Teal, P.D. A practical comparison of manual and autonomous methods for acoustic monitoring. Methods Ecol. Evol. 2013, 4, 675–683. [Google Scholar] [CrossRef]
- Puyana-Romero, V.; Ciaburro, G.; Brambilla, G.; Garzón, C.; Maffei, L. Representation of the soundscape quality in urban areas through colours. Noise Mapp. 2019, 6, 8–21. [Google Scholar] [CrossRef]
- Pijanowski, B.C.; Farina, A.; Gage, S.H.; Dumyahn, S.L.; Krause, B.L. What is soundscape ecology? An introduction and overview of an emerging new science. Landsc. Ecol. 2011, 26, 1213–1232. [Google Scholar] [CrossRef]
- Pijanowski, B.C.; Villanueva-Rivera, L.J.; Dumyahn, S.L.; Farina, A.; Krause, B.L.; Napoletano, B.M.; Gage, S.H.; Pieretti, N. Soundscape ecology: The science of sound in the landscape. BioScience 2011, 61, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Kasten, E.P.; Gage, S.H.; Fox, J.; Joo, W. The remote environmental assessment laboratory’s acoustic library: An archive for studying soundscape ecology. Ecol. Inform. 2012, 12, 50–67. [Google Scholar] [CrossRef]
- Farina, A. Soundscape Ecology: Principles, Patterns, Methods and Applications; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Barber, J.R.; Crooks, K.R.; Fristrup, K.M. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol. Evol. 2010, 25, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Towsey, M.; Wimmer, J.; Williamson, I.; Roe, P. The use of acoustic indices to determine avian species richness in audio-recordings of the environment. Ecol. Inform. 2014, 21, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Pieretti, N.; Farina, A.; Morri, D. A new methodology to infer the singing activity of an avian community: The Acoustic Complexity Index (ACI). Ecol. Indic. 2011, 11, 868–873. [Google Scholar] [CrossRef]
- Boelman, N.T.; Asner, G.P.; Hart, P.J.; Martin, R.E. Multi-trophic invasion resistance in Hawaii: Bioacoustics, field surveys, and airborne remote sensing. Ecol. Appl. 2007, 17, 2137–2144. [Google Scholar] [CrossRef]
- Sueur, J.; Pavoine, S.; Hamerlynck, O.; Duvail, S. Rapid acoustic survey for biodiversity appraisal. PLoS ONE 2008, 3, e4065. [Google Scholar] [CrossRef] [Green Version]
- Villanueva-Rivera, L.J.; Pijanowski, B.C.; Doucette, J.; Pekin, B. A primer of acoustic analysis for landscape ecologists. Landsc. Ecol. 2011, 26, 1233–1246. [Google Scholar] [CrossRef]
- Depraetere, M.; Pavoine, S.; Jiguet, F.; Gasc, A.; Duvail, S.; Sueur, J. Monitoring animal diversity using acoustic indices: Implementation in a temperate woodland. Ecol. Indic. 2012, 13, 46–54. [Google Scholar] [CrossRef]
- Fuller, S.; Axel, A.C.; Tucker, D.; Gage, S.H. Connecting soundscape to landscape: Which acoustic index best describes landscape configuration? Ecol. Indic. 2015, 58, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Linke, S.; Gifford, T.; Desjonquères, C.; Tonolla, D.; Aubin, T.; Barclay, L.; Karaconstantis, C.; Kennard, M.J.; Rybak, F.; Sueur, J. Freshwater ecoacoustics as a tool for continuous ecosystem monitoring. Front. Ecol. Environ. 2018, 16, 231–238. [Google Scholar] [CrossRef]
- Metcalf, O.C.; Barlow, J.; Devenish, C.; Marsden, S.; Berenguer, E.; Lees, A.C. Acoustic indices perform better when applied at ecologically meaningful time and frequency scales. Methods Ecol. Evol. 2021, 12, 421–431. [Google Scholar] [CrossRef]
- Pavan, G.; Favaretto, A.; Bovelacci, B.; Scaravelli, D.; Macchio, S.; Glotin, H. Bioacustica E Ecoacustica Applicate Al Monitoraggio E Alla Gestione Ambientale Bioacoustics and Ecoacoustics Applied to Environmental Monitoring and Management. Riv. Ital. Acust. 2015, 39, 69–75. [Google Scholar]
- Sueur, J.; Farina, A. Ecoacoustics: The ecological investigation and interpretation of environmental sound. Biosemiotics 2015, 8, 493–502. [Google Scholar] [CrossRef]
- Royle, J.A. Modelling sound attenuation in heterogeneous environments for improved bioacoustic sampling of wildlife populations. Methods Ecol. Evol. 2018, 9, 1939–1947. [Google Scholar] [CrossRef]
- Bradfer-Lawrence, T.; Gardner, N.; Bunnefeld, L.; Bunnefeld, N.; Willis, S.G.; Dent, D.H. Guidelines for the use of acoustic indices in environmental research. Methods Ecol. Evol. 2019, 10, 1796–1807. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Luo, Y.; Mammides, C.; Cao, K.-F.; Zhu, S.; Goodale, E. The relationship between acoustic indices, elevation, and vegetation, in a forest plot network of southern China. Ecol. Indic. 2021, 129, 107942. [Google Scholar] [CrossRef]
- Opaev, A.; Gogoleva, S.; Palko, I.; Rozhnov, V. Annual acoustic dynamics are associated with seasonality in a monsoon tropical forest in South Vietnam. Ecol. Indic. 2021, 122, 107269. [Google Scholar] [CrossRef]
- Llusia, D.; Márquez, R.; Beltrán, J.F.; Benítez, M.; Do Amaral, J.P. Calling behaviour under climate change: Geographical and seasonal variation of calling temperatures in ectotherms. Glob. Chang. Biol. 2013, 19, 2655–2674. [Google Scholar] [CrossRef]
- Krause, B.; Farina, A. Using ecoacoustic methods to survey the impacts of climate change on biodiversity. Biol. Conserv. 2016, 195, 245–254. [Google Scholar] [CrossRef]
- Machado, R.B.; Aguiar, L.; Jones, G. Do acoustic indices reflect the characteristics of bird communities in the savannas of Central Brazil? Landsc. Urban Plan. 2017, 162, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Sekercioglu, C.H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Alquezar, R.D.; Machado, R.B. Comparisons between autonomous acoustic recordings and avian point counts in open woodland savanna. Wilson J. Ornithol. 2015, 127, 712–723. [Google Scholar] [CrossRef]
- Wu, Y.J.; Yang, Q.S.; Xia, L.; Feng, Z.J.; Zhou, H.M. Species diversity and distribution pattern of non-volant small mammals along the elevational gradient on eastern slope of Gongga Mountain. Acta Ecol. Sin. 2012, 32, 4318–4328. [Google Scholar]
- Bardeli, R.; Wolff, D.; Kurth, F.; Koch, M.; Tauchert, K.-H.; Frommolt, K.-H. Detecting bird sounds in a complex acoustic environment and application to bioacoustic monitoring. Pattern Recognit. Lett. 2010, 31, 1524–1534. [Google Scholar] [CrossRef]
- Buxton, R.T.; Brown, E.; Sharman, L.; Gabriele, C.M.; McKenna, M.F. Using bioacoustics to examine shifts in songbird phenology. Ecol. Evol. 2016, 6, 4697–4710. [Google Scholar] [CrossRef] [Green Version]
- Mammides, C.; Goodale, E.; Dayananda, S.K.; Kang, L.; Chen, J. Do acoustic indices correlate with bird diversity? Insights from two biodiverse regions in Yunnan Province, south China. Ecol. Indic. 2017, 82, 470–477. [Google Scholar] [CrossRef]
- Zhang, B. Diversity of Bird Communities in Shennongjia National Nature Reserve. Ph.D. Thesis, Central China Normal University, Wuhan, China, 2014. [Google Scholar]
- Liao, M.Y. Comprehensive Survey Report of Natural Resources in Shennongjia Area; China Forestry Publishing House: Beijing, China, 2015. [Google Scholar]
- Yang, Q.; Tingting, L.I.; Wang, Z.; Lin, L.; Peng, Q.; Lin, B.; Zheng, C.; University, H. Integrated assessment on ecological sensitivity for Shennongjia National Park. J. Hubei Univ. (Nat. Sci.) 2017, 39, 455–461. [Google Scholar]
- Liu, H.Y. Study om the Plateau Wetland Birdsong Acquisition and Identifications System. Master’s Thesis, Southwest Forestry University, Kunming, China, 2016. [Google Scholar]
- Boll, S. Suppression of acoustic noise in speech using spectral subtraction. IEEE Trans. Acoust. Speech Signal Process. 1979, 27, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Napoletano, B.M. Measurement, Quantification and Interpretation of Acoustic Signals within an Ecological Context. Master’s Thesis, Michigan State University, Michigan, MI, USA, 2004. [Google Scholar]
- Schafer, P.M. The Soundscape: Our Sonic Environment and the Tuning of the World; Inner Traditions: Rochester, VT, USA, 1994. [Google Scholar]
- ISO/FDIS 12913-1; Acoustics—Soundscape—Part 1: Definition and Conceptual Framework. International Standards Organization (ISO): Geneva, Switzerland, 2014; Volume 2014.
- Romero, V.P.; Maffei, L.; Brambilla, G.; Ciaburro, G. Modelling the soundscape quality of urban waterfronts by artificial neural networks. Appl. Acoust. 2016, 111, 121–128. [Google Scholar] [CrossRef]
- Welch, P. The use of fast Fourier transform for the estimation of power spectra: A method based on time averaging over short, modified periodograms. IEEE Trans. Audio Electroacoust. 1967, 15, 70–73. [Google Scholar] [CrossRef] [Green Version]
- MathWorks. Signal Processing Toolbox: User’s Guide (R2021b). 2022. Available online: https://ww2.mathworks.cn/help/signal/ref/pwelch.html?s_tid=doc_ta (accessed on 11 May 2022).
- Gage, S.H.; Axel, A.C. Visualization of temporal change in soundscape power of a Michigan lake habitat over a 4-year period. Ecol. Inform. 2014, 21, 100–109. [Google Scholar] [CrossRef]
- Zhang, J.T. Quantitative Ecology, 3rd ed.; Science Press: Beijing, China, 2018. [Google Scholar]
- Farina, A.; Gage, S.H. Ecoacoustics: The Ecological Role of Sounds; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Zheng, G.M. Ornithology, 2nd ed.; Normal University Press: Beijing, China, 2012. [Google Scholar]
- Bruni, A.; Mennill, D.J.; Foote, J.R. Dawn chorus start time variation in a temperate bird community: Relationships with seasonality, weather, and ambient light. J. Ornithol. 2014, 155, 877–890. [Google Scholar] [CrossRef]
- Eldridge, A.; Guyot, P.; Moscoso, P.; Johnston, A.; Eyre-Walker, Y.; Peck, M. Sounding out ecoacoustic metrics: Avian species richness is predicted by acoustic indices in temperate but not tropical habitats. Ecol. Indic. 2018, 95, 939–952. [Google Scholar] [CrossRef] [Green Version]
- Benocci, R.; Roman, H.E.; Bisceglie, A.; Angelini, F.; Brambilla, G.; Zambon, G. Eco-Acoustic Assessment of an Urban Park by Statistical Analysis. Sustainability 2021, 13, 7857. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Parameter |
|---|---|
| Channel | Mono, stereo |
| Format | Wav, flac |
| Sonic range | 20 Hz–48 kHz |
| Sampling rates (Hz) | 8 k–96 k |
| Storage | 2 card slots, each up to 128 GB (SDHC) or 512 GB (SDXC); Maximum capacity up to 1.0 TB (two 512 GB SDXC cards); |
| Shell | Waterproof |
| Name | Calculation Method | Symbolic Interpretation | Usage | |
|---|---|---|---|---|
| BI [19] | (1) | : the mean spectrum across ; : the minimum dB value of this mean spectrum; : list of the frequencies of the spectrogram. | Used to compare frequency amplitude spectra to detect or infer differences in avian community composition among sites [19]. | |
| ACI [18] | : number of spectrums; : number of time steps in bin; : the number of subsets in a single frequency bin; : the intensity in a single frequency bin. | Used to determine changes in behavior and composition of a vocalizing community and consequently to better monitor animal dynamics in a quick way [18]. | ||
| ADI [21] | : the proportion of the signals in each bin above a threshold (−50 dBFS), ; : the number of spectrograms. | Representing sound diversity used to compare the soundscape differences between different times among different sites [21]. | ||
| AEI [21] | Used to compare the frequency differences between sounds from different land usages [21]. | |||
| H [20] | (5) (6) | : length of the signal in number of digitized points; : probability mass function of the amplitude envelope; : probability mass function of the mean spectrum calculated. | Derived from the Shannon information statistic, higher values of H indicates richer habitats [20]. | |
| NDSI [14] | (7) | : biophony; : anthrophony; NDSI < 1 the levels of anthrophonies is higher; NDSI < 1 the levels of biophonies is higher. | Used to estimate the level of anthropogenic disturbance on the soundscape [14]. | |
| Sampling Sites | Proportion of SP in Soundscape (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1~2 | 2~3 | 3~4 | 4~5 | 5~6 | 6~7 | 7~8 | 8~9 | 9~10 | 10~11 | |
| Site 3 | 30.0 | 37.2 | 13.6 | 6.4 | 3.8 | 3.8 | 2.6 | 1.8 | 1.8 | 0.3 |
| Site 5 | 43.9 | 38.9 | 7.5 | 5.4 | 1.7 | 1.0 | 0.7 | 0.4 | 0.4 | 0.2 |
| Site 7 | 33.8 | 39.1 | 15.2 | 4.6 | 2.3 | 1.9 | 1.4 | 0.9 | 0.7 | 0.2 |
| Site 8 | 52.1 | 32.8 | 6.0 | 3.0 | 1.5 | 1.4 | 1.5 | 0.8 | 0.6 | 0.2 |
| Site 11 | 36.1 | 32.2 | 17.9 | 5.7 | 3.0 | 2.1 | 1.4 | 0.8 | 0.6 | 0.1 |
| Sampling Sites | BI | ACI | ADI | AEI | H | NDSI |
|---|---|---|---|---|---|---|
| Site 3 | 0.226 | 0.544 ** | 0.118 | −0.010 | −0.383 | 0.485 * |
| Site 5 | 0.610 ** | 0.698 ** | 0.127 | −0.154 | −0.192 | 0.698 ** |
| Site 7 | 0.595 ** | 0.281 | −0.002 | 0.010 | −0.270 | 0.669 ** |
| Site 8 | 0.439 * | 0.381 | −0.232 | 0.260 | −0.447 * | 0.503 ** |
| Site 11 | 0.351 * | 0.736 ** | 0.414 * | −0.474 ** | 0.475 ** | 0.225 |
| Sampling Sites | BI | ACI | ADI | AEI | H | NDSI |
|---|---|---|---|---|---|---|
| Site 3 | 0.321 | 0.824 ** | −0.312 | 0.466 * | −0.372 | 0.509 * |
| Site 5 | 0.617 ** | 0.706 ** | −0.039 | 0.116 | −0.311 | 0.696 ** |
| Site 7 | 0.617 ** | 0.751 ** | −0.049 | 0.194 | −0.404 | 0.645 ** |
| Site 8 | 0.322 | 0.581 * | −0.131 | 0.121 | −0.427 * | 0.481 * |
| Site 11 | 0.661 ** | 0.806 ** | 0.355 * | −0.450 ** | 0.187 | 0.225 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.; Yu, X.; Yang, J.; Ouyang, X.; Fan, D. Acoustic Sensor-Based Soundscape Analysis and Acoustic Assessment of Bird Species Richness in Shennongjia National Park, China. Sensors 2022, 22, 4117. https://doi.org/10.3390/s22114117
Hou Y, Yu X, Yang J, Ouyang X, Fan D. Acoustic Sensor-Based Soundscape Analysis and Acoustic Assessment of Bird Species Richness in Shennongjia National Park, China. Sensors. 2022; 22(11):4117. https://doi.org/10.3390/s22114117
Chicago/Turabian StyleHou, Yanan, Xinwen Yu, Jingyuan Yang, Xuan Ouyang, and Dongpu Fan. 2022. "Acoustic Sensor-Based Soundscape Analysis and Acoustic Assessment of Bird Species Richness in Shennongjia National Park, China" Sensors 22, no. 11: 4117. https://doi.org/10.3390/s22114117
APA StyleHou, Y., Yu, X., Yang, J., Ouyang, X., & Fan, D. (2022). Acoustic Sensor-Based Soundscape Analysis and Acoustic Assessment of Bird Species Richness in Shennongjia National Park, China. Sensors, 22(11), 4117. https://doi.org/10.3390/s22114117