
Detection of Antibodies against Hepatitis A Virus (HAV) by a Surface Plasmon Resonance (SPR) Biosensor: A New Diagnosis Tool Based on the Major HAV Capsid Protein VP1 (SPR-HAVP1)


 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Serum Samples from Patients
2.3. IgM Purification
2.4. HAV Samples in Cell Culture
2.5. Prediction of VP1 Protein B-Lymphocyte Epitopes
2.6. SDS Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.7. Immunoenzymatic Assays
2.7.1. Western Blotting
2.7.2. Dot Blot
2.8. Establishment of the SPR-HAVP1 Assays
2.8.1. Immobilization of VP1
2.8.2. Standardization of Anti-HAV IgM Binding
2.9. Detection Assay of Anti-HAV Antibody in Serum Samples
2.10. Data Analysis
3. Results
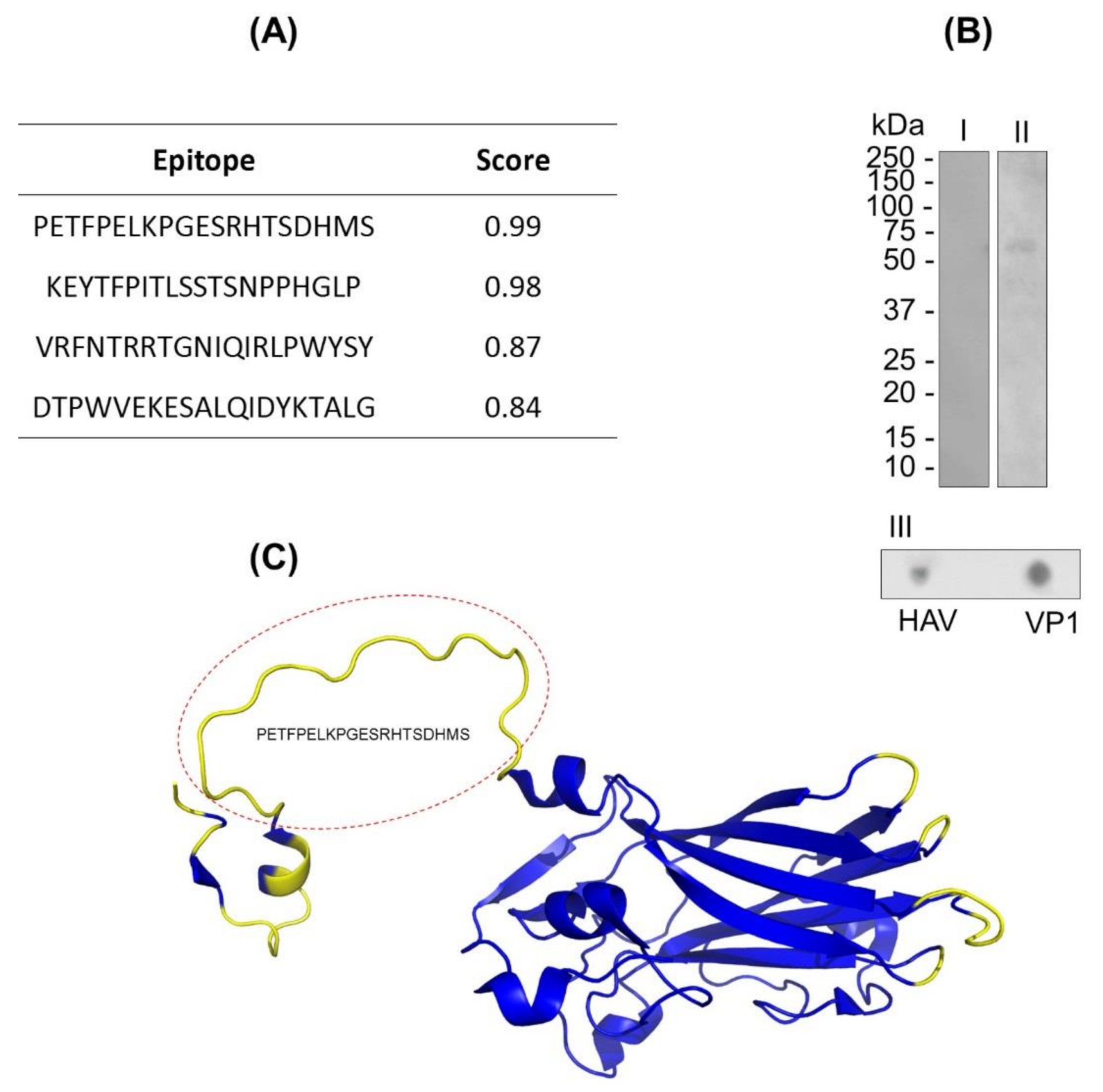
3.1. In Silico Analysis of the B-Lymphocyte Epitopes Predictions of VP1
3.2. In Vitro Analysis of Continuous and Discontinuous Epitopes in VP1
3.3. SPR Assays to Detect Anti-VP1 Immunoglobulin
3.3.1. Sensor Chip Stability and Regeneration
3.3.2. Interaction of IgM anti-HAV Antibodies with VP1
3.3.3. Assessing the anti-HAV Antibodies in Human Serum Samples by SPR-HAVP1 Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stapleton, J.T.; Lemon, S.M. Neutralization escape mutants define a dominant immunogenic neutralization site on hepatitis A virus. J. Virol. 1987, 61, 491–498. [Google Scholar] [CrossRef]
- Ping, L.H.; Jansen, R.W.; Stapleton, J.T.; Cohen, J.I.; Lemon, S.M. Identification of an immunodominant antigenic site involving the capsid protein VP3 of hepatitis A virus. Proc. Natl. Acad. Sci. USA 1988, 85, 8281–8285. [Google Scholar] [CrossRef]
- Gómara, M.; Ercilla, G.; Alsina, M.; Haro, I. Assessment of synthetic peptides for hepatitis A diagnosis using biosensor technology. J. Immunol. Methods 2000, 246, 13–24. [Google Scholar] [CrossRef]
- Linder, K.A.; Malani, P.N. Hepatitis A. JAMA 2017, 318, 2393. [Google Scholar] [CrossRef]
- World Health Organization. Position Paper on Hepatitis A Vaccines—June 2012. Relev. Epidemiol. Hebd. 2012, 87, 261–276. [Google Scholar]
- World Health Organization. Hepatitis A Outbreaks Mostly Affecting Men Who Have Sex With Men—European Region and the Americas. Available online: https://www.who.int/news/item/07-06-2017-hepatitis-a-outbreaks-mostly-affecting-men-who-have-sex-with-men-european-region-and-the-americas (accessed on 25 November 2020).
- Gozlan, Y.; Bar-Or, I.; Rakovsky, A.; Savion, M.; Amitai, Z.; Sheffer, R.; Ceder, N.; Anis, E.; Grotto, I.; Mendelson, E.; et al. Ongoing hepatitis A among men who have sex with men (MSM) linked to outbreaks in Europe in Tel Aviv area, Israel, December 2016—June 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Jeong, H.S.; Cho, B.-K.; Ji, M.-J.; Kim, J.-H.; Lee, A.-N.; Lee, K.-R.; Cheon, D.-S. Evaluation of an immunochromatographic assay for the detection of anti-hepatitis A virus IgM. Virol. J. 2010, 7, 164. [Google Scholar] [CrossRef] [PubMed]
- Lemon, S.M.; Ott, J.J.; Van Damme, P.; Shouval, D. Type A viral hepatitis: A summary and update on the molecular virology, epidemiology, pathogenesis and prevention. J. Hepatol. 2018, 68, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Wang, H.; Fu, Q.; Peng, J.; Wang, Y.; Du, J.; Zhou, Y.; Zhan, L. Gold nanorod-based localized surface plasmon resonance biosensor for sensitive detection of hepatitis B virus in buffer, blood serum and plasma. Biosens. Bioelectron. 2010, 26, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Tam, Y.J.; Zeenathul, N.A.; Rezaei, M.A.; Mustafa, N.H.; Azmi, M.L.M.; Bahaman, A.R.; Lo, S.C.; Tan, J.S.; Hani, H.; Rasedee, A. Wide dynamic range of surface-plasmon-resonance-based assay for hepatitis B surface antigen antibody optimal detection in comparison with ELISA. Biotechnol. Appl. Biochem. 2017, 64, 735–744. [Google Scholar] [CrossRef]
- Riedel, T.; Hageneder, S.; Surman, F.; Pop-Georgievski, O.; Noehammer, C.; Hofner, M.; Brynda, E.; Rodriguez-Emmenegger, C.; Dostálek, J. Plasmonic Hepatitis B Biosensor for the Analysis of Clinical Saliva. Anal. Chem. 2017, 89, 2972–2977. [Google Scholar] [CrossRef]
- Das, C.M.; Guo, Y.; Kang, L.; Ho, H.; Yong, K. Investigation of Plasmonic Detection of Human Respiratory Virus. Adv. Theory Simulations 2020, 3, 2000074. [Google Scholar] [CrossRef]
- Singh, P. Surface Plasmon Resonance: A Boon for Viral Diagnostics. Ref. Modul. Life Sci. 2017. [Google Scholar] [CrossRef]
- Lee, J.; Takemura, K.; Park, E.Y. Plasmonic Nanomaterial-Based Optical Biosensing Platforms for Virus Detection. Sensors 2017, 17, 2332. [Google Scholar] [CrossRef]
- Mauriz, E. Recent Progress in Plasmonic Biosensing Schemes for Virus Detection. Sensors 2020, 20, 4745. [Google Scholar] [CrossRef] [PubMed]
- Villar, L.; Amado, L.; Gaspar, A. In Situ enzyme immunoassay for titration of a Brazilian hepatitis A virus strain (HAF-203). Braz. J. Med Biol. Res. 2004, 37, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, A.M.; Vitral, C.L.; Yoshida, C.F.; Schatzmayr, H.G. Primary isolation of a Brazilian strain of hepatitis A virus (HAF-203) and growth in a primate cell line (FRhK-4). Braz. J. Med Biol. Res. 1992, 25, 697–705. [Google Scholar]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Santos-De-Souza, R.; Souza-Silva, F.; De Albuquerque-Melo, B.C.; Ribeiro-Guimarães, M.L.; Côrtes, L.M.D.C.; Pereira, B.A.S.; Silva-Almeida, M.; Cysne-Finkelstein, L.; Junior, F.O.R.D.O.; Pereira, M.C.D.S.; et al. Insights into the tracking of the cysteine proteinase B COOH-terminal polypeptide of Leishmania (Leishmania) amazonensis by surface plasmon resonance. Parasitol. Res. 2019, 118, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Junior, H.C.D.S.; Da Silva, E.D.; Rodrigues, L.L.L.-X.D.S.; Medeiros, M.A. Recombinant VP1 protein as a potential marker for the diagnosis of acute hepatitis A virus infection. J. Virol. Methods 2017, 245, 1–4. [Google Scholar] [CrossRef]
- Provost, P.J.; Hilleman, M.R. Propagation of Human Hepatitis A Virus in Cell Culture in Vitro. Exp. Biol. Med. 1979, 160, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, C.M.; Fields, H.A.; Schable, C.A.; Meinke, W.J.; Maynard, J.E. Adsorption, purification, and growth characteristics of hepatitis A virus strain HAS-15 propagated in fetal rhesus monkey kidney cells. J. Clin. Microbiol. 1986, 23, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Ping, L.H.; Lemon, S.M. Antigenic structure of human hepatitis A virus defined by analysis of escape mutants selected against murine monoclonal antibodies. J. Virol. 1992, 66, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.L.; Silva, M.; De Lima, M.A.; Yoshida, C.F.; Gaspar, A.M.C.; Galler, R. Genetic variability of hepatitis A virus strain HAF-203 isolated in Brazil and expression of the VP1 gene in Escherichia coli. Memórias Inst. Oswaldo Cruz 2006, 101, 759–766. [Google Scholar] [CrossRef][Green Version]
- El-Manzalawy, Y.; Dobbs, D.; Honavar, V.G. In Silico Prediction of Linear B-Cell Epitopes on Proteins. Methods Mol. Biol. 2016, 1484, 255–264. [Google Scholar] [CrossRef]
- Lofas, S.; Johnsson, B.; Tegendal, K.; Rönnberg, I. Dextran modified gold surfaces for surface plasmon resonance sensors: Immunoreactivity of immobilized antibodies and antibody-surface interaction studies. Colloids Surf. B Biointerfaces 1993, 1, 83–89. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, G.M.C.d.; Alves, C.R.; Pinto, M.A.; Amado Leon, L.A.; Souza-Silva, F. Detection of Antibodies against Hepatitis A Virus (HAV) by a Surface Plasmon Resonance (SPR) Biosensor: A New Diagnosis Tool Based on the Major HAV Capsid Protein VP1 (SPR-HAVP1). Sensors 2021, 21, 3167. https://doi.org/10.3390/s21093167
Santos GMCd, Alves CR, Pinto MA, Amado Leon LA, Souza-Silva F. Detection of Antibodies against Hepatitis A Virus (HAV) by a Surface Plasmon Resonance (SPR) Biosensor: A New Diagnosis Tool Based on the Major HAV Capsid Protein VP1 (SPR-HAVP1). Sensors. 2021; 21(9):3167. https://doi.org/10.3390/s21093167
Chicago/Turabian StyleSantos, Gabriel Menezes Costa dos, Carlos Roberto Alves, Marcelo Alves Pinto, Luciane Almeida Amado Leon, and Franklin Souza-Silva. 2021. "Detection of Antibodies against Hepatitis A Virus (HAV) by a Surface Plasmon Resonance (SPR) Biosensor: A New Diagnosis Tool Based on the Major HAV Capsid Protein VP1 (SPR-HAVP1)" Sensors 21, no. 9: 3167. https://doi.org/10.3390/s21093167
APA StyleSantos, G. M. C. d., Alves, C. R., Pinto, M. A., Amado Leon, L. A., & Souza-Silva, F. (2021). Detection of Antibodies against Hepatitis A Virus (HAV) by a Surface Plasmon Resonance (SPR) Biosensor: A New Diagnosis Tool Based on the Major HAV Capsid Protein VP1 (SPR-HAVP1). Sensors, 21(9), 3167. https://doi.org/10.3390/s21093167