Rapid On-Site Phenotyping via Field Fluorimeter Detects Differences in Photosynthetic Performance in a Hybrid—Parent Barley Germplasm Set

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Samplings
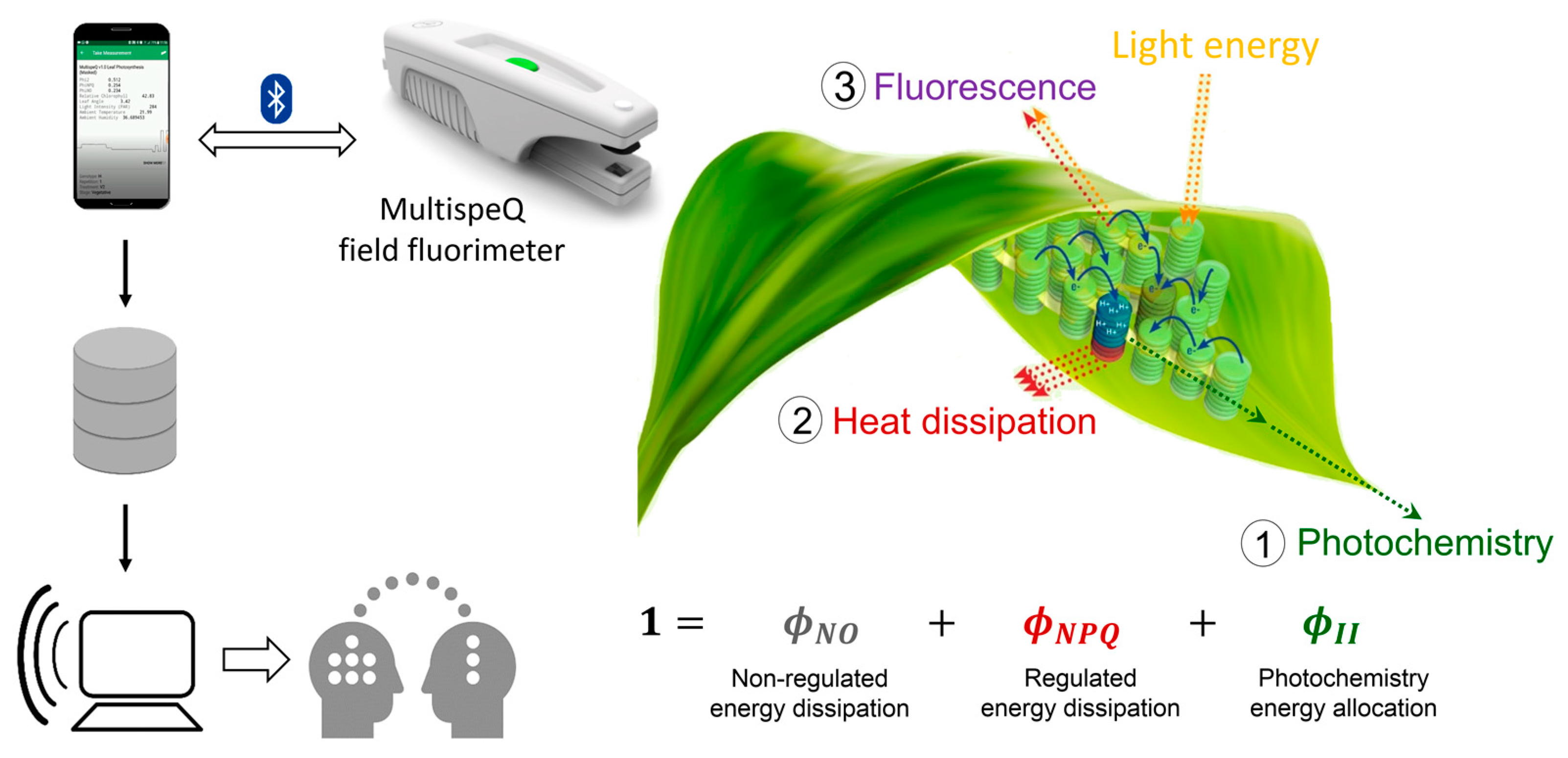
2.3. Chlorophyll Fluorescence, Absorbance and Environmental Variables Measurements
2.4. Photosynthetic and Physiological Parameter Calculations
2.5. Statistical Analysis
3. Results
3.1. Chlorophyll Fluorescence-Derived Parameters
3.2. Absorbance-Based Parameters
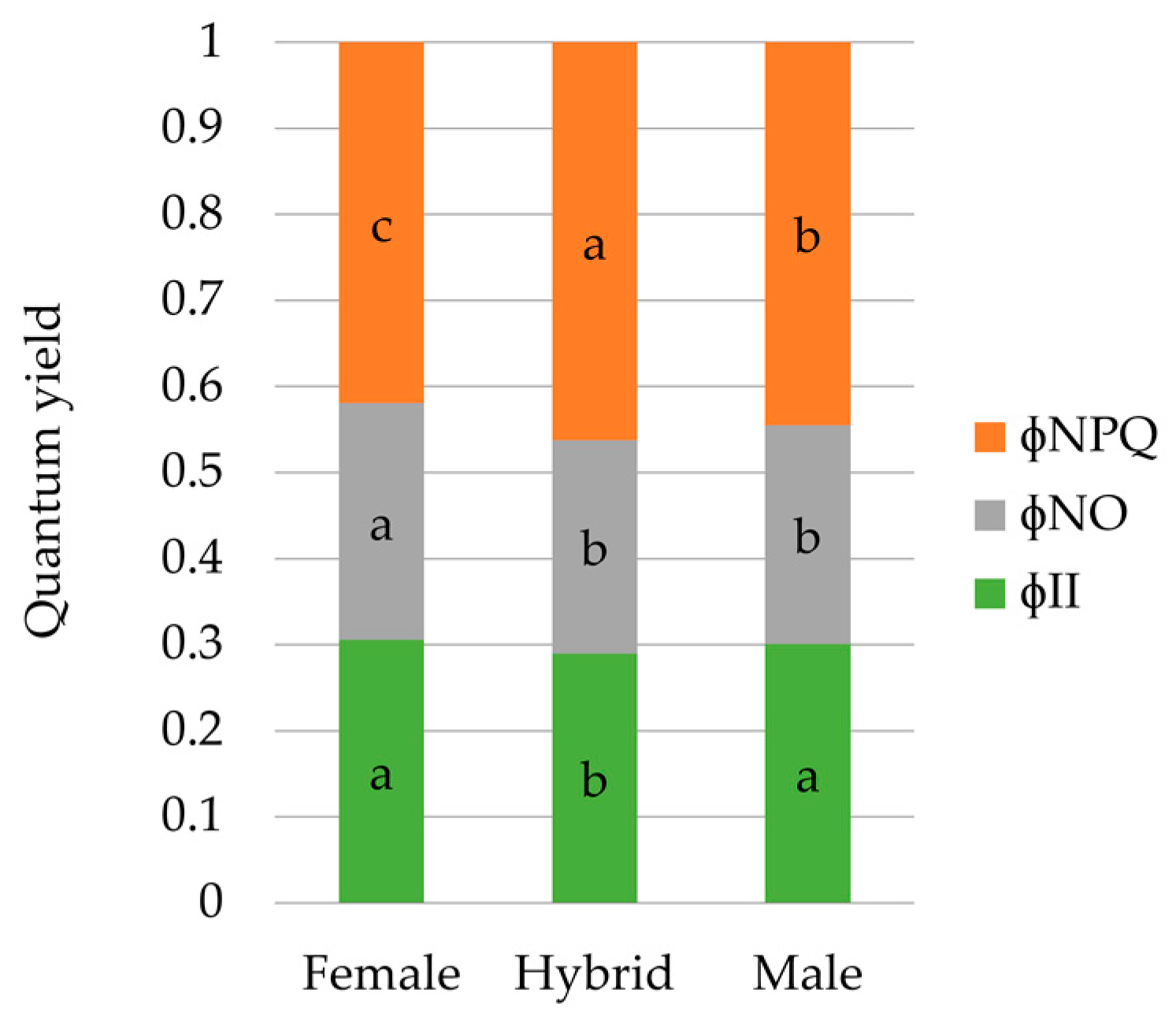
3.3. PSII Energy-Absorbed Allocation Proportions
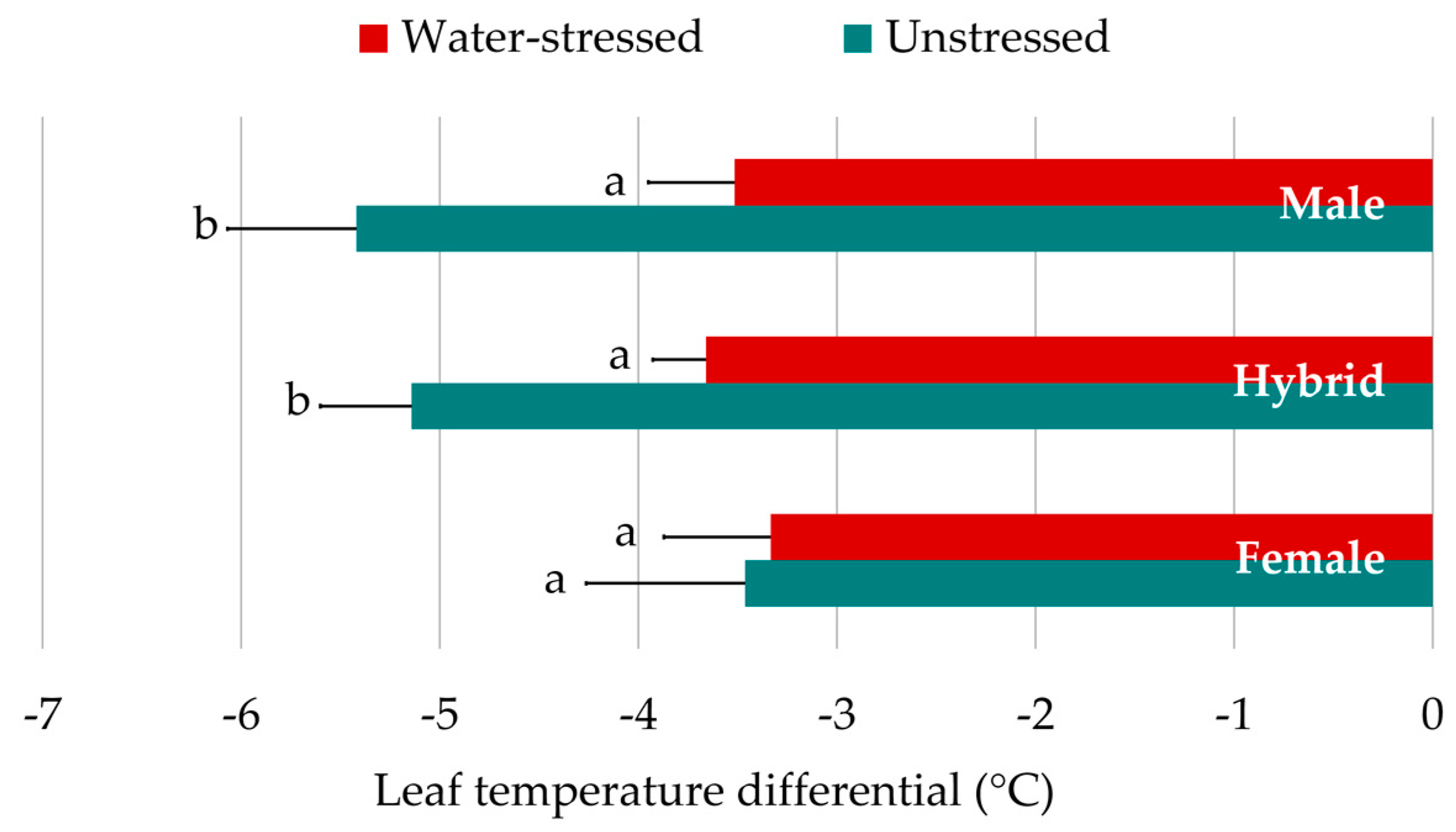
3.4. Leaf Temperature Differential
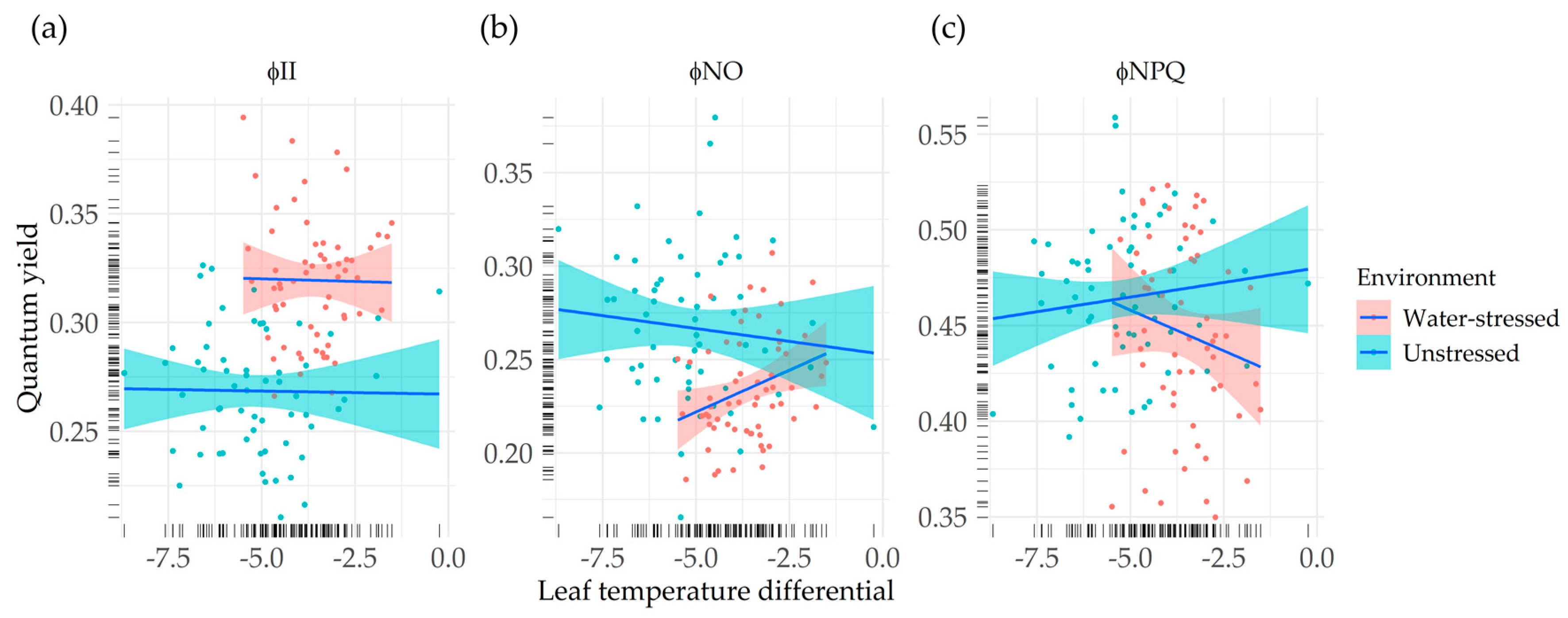
3.5. Relationship between Chlorophyll Fluorescence-Based Parameters and Crop Status Indicators
4. Discussion
4.1. Drought Stress Indicators
4.2. Photoprotective Response of Hybrids and Fitness under Unfavorable Conditions
4.3. Suitability of MultispeQ as a Tool to Screen Plant Populations for Stress Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Flavell, R.B. Innovations continuously enhance crop breeding and demand new strategic planning. Glob. Food Sec. 2017, 12, 15–21. [Google Scholar] [CrossRef]
- Desta, Z.A.; Ortiz, R. Genomic selection: Genome-wide prediction in plant improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Cobb, J.N.; DeClerck, G.; Greenberg, A.; Clark, R.; McCouch, S. Next-generation phenotyping: Requirements and strategies for enhancing our understanding of genotype-phenotype relationships and its relevance to crop improvement. Theor. Appl. Genet. 2013, 126, 867–887. [Google Scholar] [CrossRef] [PubMed]
- Fiorani, F.; Schurr, U. Future Scenarios for Plant Phenotyping. Annu. Rev. Plant Biol. 2013, 64, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Furbank, R.T.; Tester, M. Phenomics-technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef]
- Pauli, D.; Chapman, S.C.; Bart, R.; Topp, C.N.; Lawrence-Dill, C.J.; Poland, J.; Gore, M.A. The quest for understanding phenotypic variation via integrated approaches in the field environment. Plant Physiol. 2016, 172, 622–634. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef]
- Reynolds, M.; Langridge, P. Physiological breeding. Curr. Opin. Plant Biol. 2016, 31, 162–171. [Google Scholar] [CrossRef]
- Reynolds, M.; Tattaris, M.; Cossani, C.M.; Ellis, M.; Yamaguchi-Shinozaki, K.; Saint Pierre, C.; Reynolds, M.; Tattaris, M.; Cossani, C.M.; Ellis, M.; et al. Exploring genetic resources to increase adaptation of wheat to climate change. In Advances in wheat genetics: From genome to field; Springer: Tokio, Japan, 2015; pp. 355–368. [Google Scholar]
- Monteith, J.L. Climate and the Efficiency of Crop Production in Britain. Philos. Trans. R. Soc. B Biol. Sci. 1977, 281, 277–294. [Google Scholar]
- Furbank, R.T.; Jimenez-Berni, J.A.; George-Jaeggli, B.; Potgieter, A.B.; Deery, D.M. Field crop phenomics: Enabling breeding for radiation use efficiency and biomass in cereal crops. New Phytol. 2019, 223, 1714–1727. [Google Scholar] [CrossRef]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A tool for large-scale plant phenotyping connected to the open PhotosynQ network. R. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Brestic, M.; Zivcak, M. PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: Protocols and applications. In Molecular Stress Physiology of Plants; Springer: Delhi, India, 2013; pp. 87–131. ISBN 9788132208075. [Google Scholar]
- Kuckenberg, J.; Tartachnyk, I.; Noga, G. Temporal and spatial changes of chlorophyll fluorescence as a basis for early and precise detection of leaf rust and powdery mildew infections in wheat leaves. Precis. Agric. 2009, 10, 34–44. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Miehé, J. Fluorescence imaging as a diagnostic tool for plant stress. Trends Plant Sci. 1997, 2, 316–320. [Google Scholar] [CrossRef]
- Massacci, A.; Nabiev, S.M.; Pietrosanti, L.; Nematov, S.K.; Chernikova, T.N.; Thor, K.; Leipner, J. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol. Biochem. 2008, 46, 189–195. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence - a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Boyer, J.S.; Westgate, M.E. Grain yields with limited water. J. Exp. Bot. 2004, 55, 2385–2394. [Google Scholar] [CrossRef]
- Cattivelli, L.; Ceccarelli, S.; Romagosa, I.; Stanca, M. Abiotic Stresses in Barley: Problems and Solutions. In Barley: Production, Improvement, and Uses; Ullrich, S.E., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2011; pp. 282–306. [Google Scholar]
- Trenberth, K.E.; Jones, P.D.; Authors, L.; Ambenje, P.G.; Bojariu, R.; Easterling, D.R.; Klein Tank, A.M.G.; Renwick, J.A.; Rahimzadeh, F.; Rusticucci, M.M.; et al. Observations: Surface and Atmospheric Climate Change. In Climate Change 2007: The Physical Science Basis; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 235–336. [Google Scholar]
- Hu, H.; Xiong, L. Genetic Engineering and Breeding of Drought-Resistant Crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Mühleisen, J.; Maurer, H.P.; Stiewe, G.; Bury, P.; Reif, J.C. Hybrid breeding in barley. Crop Sci. 2013, 53, 819–824. [Google Scholar] [CrossRef]
- Longin, C.F.H.; Mühleisen, J.; Maurer, H.; Zhang, H.; Gowda, M.; Reif, J. Hybrid breeding in autogamous cereals. Theor. Appl. Genet. 2012, 125, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Mühleisen, J.; Piepho, H.-P.; Maurer, H.P.; Longin, C.F.H.; Reif, J.C. Yield stability of hybrids versus lines in wheat, barley, and triticale. Theor. Appl. Genet. 2014, 127, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Gracia, M.P.; Mansour, E.; Casas, A.M.; Lasa, J.M.; Medina, B.; Molina-Cano, J.L.; Moralejo, M.A.; López, A.; López-Fuster, P.; Escribano, J.; et al. Progress in the Spanish National Barley Breeding Program. Spanish J. Agric. Res. 2012, 10, 741. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Rinderle, U.; Schmuck, G. Application of chlorophyll fluorescence in ecophysiology. Radiat. Environ. Biophys. 1986, 25, 297–308. [Google Scholar] [CrossRef]
- Tietz, S.; Hall, C.C.; Cruz, J.A.; Kramer, D.M. NPQ(T): A chlorophyll fluorescence parameter for rapid estimation and imaging of non-photochemical quenching of excitons in photosystem-II-associated antenna complexes. Plant. Cell Environ. 2017, 40, 1243–1255. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Krall, J.P.; Edwards, G.E. Relationship between photosystem II activity and CO2 fixation in leaves. Physiol. Plant. 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Lyman Ott, R. An Introduction to Statistical Methods and Data Analysis, 3rd ed.; PWS-KENT Publishing Co: Boston, MA, USA, 1991. [Google Scholar]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley landraces exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef]
- Sarkar, R.K.; Panda, D.; Reddy, J.N.; Patnaik, S.S.C.; Mackill, D.J.; Ismail, A.M. Performance of submergence tolerant rice (Oryza sativa) genotypes carrying the Sub1 quantitative trait locus under stressed and non-stressed natural field conditions. Indian J. Agric. Sci. 2009, 79, 876–883. [Google Scholar]
- O’Neill, P.M.; Shanahan, J.F.; Schepers, J.S. Use of chlorophyll fluorescence assessments to differentiate corn hybrid response to variable water conditions. Crop Sci. 2006, 46, 681–687. [Google Scholar] [CrossRef]
- Bilger, W.; Schreiber, U.; Bock, M. Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyll fluorescence in the field. Oecologia 1995, 102, 425–432. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Guo, P. Chlorophyll fluorescence: A useful tool in barley plant breeding programs. In Photochemistry Research Progress; Sánchez, A., Gutiérrez, S.J., Eds.; Nova Sci. Publ. Inc: New York, NY, USA, 2008; pp. 439–463. ISBN 978-1-60456-568-3. [Google Scholar]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Yao, J.; Sun, D.; Cen, H.; Xu, H.; Weng, H.; Yuan, F.; He, Y. Phenotyping of Arabidopsis drought stress response using kinetic chlorophyll fluorescence and multicolor fluorescence imaging. Front. Plant Sci. 2018, 9, 603. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7. [Google Scholar] [CrossRef]
- Brestic, M.; Cornic, G.; Fryer, M.J.; Baker, N.R. Does photorespiration protect the photosynthetic apparatus in French bean leaves from photoinhibition during drought stress? Planta 1995, 196, 450–457. [Google Scholar] [CrossRef]
- Morales, F.; Abadía, A.; AbadÞa, J. Photoinhibition and Photoprotection under Nutrient Deficiencies, Drought and Salinity. In Photoprotection, Photoinhibition, Gene Regulation, and Environment; Demmig-Adams, B., Adams III, W.W., Mattoo, A.K., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2006; pp. 65–85. [Google Scholar]
- Kitao, M.; Lei, T.T. Circumvention of over-excitation of PSII by maintaining electron transport rate in leaves of four cotton genotypes developed under long-term drought. Plant Biol. (Stuttg.) 2007, 9, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.; Lichtenthaler, H. Photosynthetic CO2-assimilation, chlorophyll fluorescence and zeaxanthin accumulation in field grown maple trees in the course of a sunny and a cloudy day. J. Plant Physiol. 1996, 148, 399–412. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. In Vivo Chlorophyll Fluorescence as a Tool for Stress Detection in Plants. In Applications of Chlorophyll Fluorescence in Photosynthesis Research, Stress Physiology, Hydrobiology and Remote Sensing; Springer Netherlands: Dordrecht, The Netherlands, 1988; pp. 129–142. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Baker, N.R.; Oxborough, K. Chlorophyll Fluorescence as a Probe of Photosynthetic Productivity. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 65–82. [Google Scholar]
- Li, R.H.; Guo, P.G.; Baum, M.; Grando, S.; Ceccarelli, S. Evaluation of Chlorophyll Content and Fluorescence Parameters as Indicators of Drought Tolerance in Barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Nyachiro, J.M.; Briggs, K.G.; Hoddinott, J.; Johnson-Flanagan, A.M. Chlorophyll Content, Chlorophyll Fluorescence and Water Deficit in Spring Wheat. Cereal Res. Commun. 2001, 29, 135–142. [Google Scholar] [CrossRef]
- Benbella, M.; Paulsen, G.M. Efficacy of treatments for delaying senescence of wheat leaves: II. Senescence and grain yield under field conditions. Agron. J. 1998, 90, 332–338. [Google Scholar] [CrossRef]
- Borrell, A.K.; Hammer, G.L.; Henzell, R.G. Does maintaining green leaf area in sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci. 2000, 40, 1037–1048. [Google Scholar] [CrossRef]
- Liu, Y.; Subhash, C.; Yan, J.; Song, C.; Zhao, J.; Li, J. Maize leaf temperature responses to drought: Thermal imaging and quantitative trait loci (QTL) mapping. Environ. Exp. Bot. 2011, 71, 158–165. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Nasyrov, Y.S. Genetic Control of Photosynthesis and Improving of Crop Productivity. Annu. Rev. Plant Physiol. 1978, 29, 215–237. [Google Scholar] [CrossRef]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F.; Sachetto-Martins, G.; Surpin, M.; Lim, J.; Mittler, R.; Chory, J. Signals from Chloroplasts Converge to Regulate Nuclear Gene Expression. Science 2007, 316, 715–719. [Google Scholar] [CrossRef]
- Mohammed, G.H.; Colombo, R.; Middleton, E.M.; Rascher, U.; van der Tol, C.; Nedbal, L.; Goulas, Y.; Pérez-Priego, O.; Damm, A.; Meroni, M.; et al. Remote sensing of solar-induced chlorophyll fluorescence (SIF) in vegetation: 50 years of progress. Remote Sens. Environ. 2019, 231, 111177. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping Plant Responses to Biotic Stress by Chlorophyll Fluorescence Imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment | PAR (μmol m−2 s−1)* | Tm (°C)* | P (mm)* | R (W m−2)* | RH (%)* |
|---|---|---|---|---|---|
| Water-stressed | 1350 | 13.8 | 9.6 | 260.4 | 64.7 |
| Unstressed | 1404 | 17.4 | 90.2 | 285.8 | 75.6 |
| Source of Variation | df | RC | LTD | ΦII | ΦNO | ΦNPQ | NPQt | qL | qP |
|---|---|---|---|---|---|---|---|---|---|
| Sqrt (PAR) | 1 | 1655 *** | 3.5 | 3.7 × 10−1*** | 9.2 × 10−3 | 2.6 × 10−1*** | 3.6 | 0.341 *** | 0.661 *** |
| Genotypic sets (GS) | 3 | 323 ** | 26.1 * | 5.6 × 10−3* | 2.3 × 10−2*** | 2.3 × 10−2** | 5.5 ** | 0.026 *** | 0.015 ** |
| F1 vs Parents | 1 | 67 | 8.8 | 5.5 × 10−3· | 2.4 × 10−2* | 5.3 × 10−2** | 10.4 ** | 0.017 * | 0.000 |
| F1 vs Females | 1 | 906 *** | 64.9 ** | 8.3 × 10−6 | 4.4 × 10−2*** | 4.6 × 10−2** | 10.6 ** | 0.049 *** | 0.020 * |
| F1 vs Males | 1 | 30 | 0.3 | 6.9 × 10−3* | 5.3 × 10−3 | 2.4 × 10−2* | 4.3· | 0.002 | 0.003 |
| Females vs Males | 1 | 958 *** | 65.7 ** | 2.0 × 10−4 | 3.8 × 10−2** | 3.3 × 10−2* | 7.9 * | 0.044 ** | 0.023 * |
| F1 (FemA) vs F1 (FemB) | 1 | 11 | 12.0 | 9.9 × 10−3* | 2.1 × 10−2* | 2.0 × 10−3 | 2.3 | 0.027 ** | 0.021 * |
| Genotypes within GS | 57 | 53 ** | 6.7 * | 1.7 × 10−3 | 3.6 × 10−3 | 5.0 × 10−3 | 1.1 | 0.004 | 0.003 |
| Environment (ENV) | 1 | 54,009 *** | 317.3 *** | 2.7 × 10−1*** | 1.3 × 10−1*** | 2.3 × 10−2· | 14.6 ** | 0.049 *** | 0.636 *** |
| Device | 1 | 1363 *** | 359.6 *** | 1.0 × 10−1*** | 1.6 × 10−1*** | 5.2 × 10−1*** | 80.8*** | 0.069*** | 0.000 |
| Genotypic set*ENV | 3 | 86· | 7.43 | 2.9 × 10−3 | 1.1 × 10−2 | 3.0 × 10−3 | 0.6 | 0.013 | 0.010 |
| F1 vs Parents*ENV | 1 | 170 * | 0.3 | 3.0 × 10−4 | 3.4 × 10−3 | 1.7 × 10−3 | 0.1 | 0.000 | 0.000 |
| F1 vs Females*ENV | 1 | 95 | 19.0· | 2.7 × 10−5 | 1.0 × 10−4 | 2.0 × 10−4 | 0.2 | 0.001 | 0.002 |
| F1 vs Males*ENV | 1 | 102· | 2.3 | 5.0 × 10−4 | 3.9 × 10−3 | 1.6 × 10−3 | 0.0 | 0.001 | 0.000 |
| Females vs Males*ENV | 1 | 59 | 21.4· | 1.0 × 10−4 | 1.0 × 10−5 | 4.0 × 10−5 | 0.1 | 0.002 | 0.002 |
| F1 (FemA) vs F1 (FemB)*ENV | 1 | 75 | 0.0 | 8.1 × 10−3* | 3.1 × 10−2* | 7.2 × 10−3 | 1.5 | 0.035 * | 0.027 * |
| Genotypes within GS*ENV | 57 | 35 | 5.7 | 1.7 × 10−3 | 6.1 × 10−3 | 7.5 × 10−3 | 1.7 | 0.006 * | 0.004· |
| Residuals | 412 | 31 | 4.5 | 1.9 × 10−3 | 4.8 × 10−3 | 6.6 × 10−3 | 1.5 | 0.004 | 0.003 |
| Source of variation | df | LEF | RFd | Fm′ | F0′ | Fs | Fv′/Fm′ | ||
| sqrt(PAR) | 1 | 164,327 *** | 1.753 *** | 2.2 × 108 *** | 1.3 × 107 *** | 3.0 × 106 | 1.78 × 10−2· | ||
| Genotypic sets (GS) | 3 | 2329 * | 0.020* | 8.1 × 106 | 8.7 × 105 | 4.9 × 106 | 2.26 × 10−2 ** | ||
| F1 vs Parents | 1 | 2271· | 0.022· | 2.1 × 107 | 6.7 × 104 | 5.5 × 106 | 3.97 × 10−2** | ||
| F1 vs Females | 1 | 12 | 0.000 | 4.0 × 106 | 2.3 × 106· | 2.1 × 106 | 3.97 × 10−2** | ||
| F1 vs Males | 1 | 3149 * | 0.026· | 1.8 × 107 | 2.0 × 105 | 3.8 × 106 | 4.67 × 10−2** | ||
| Females vs Males | 1 | 216 | 0.000 | 2.0 × 105 | 2.5 × 106· | 1.1 × 106 | 1.46 × 10−2· | ||
| F1 (FemA) vs F1 (FemB) | 1 | 3792 * | 0.033 * | 3.6 × 106 | 8.0 × 103 | 9.0 × 106 | 3.62 × 10−2** | ||
| Genotypes within GS | 57 | 633 | 0.007 | 1.0 × 107 | 7.3 × 105 | 5.4 × 106 | 4.08 × 10−3 | ||
| Environment (ENV) | 1 | 111,993 *** | 1.120 *** | 1.0 × 108** | 2.2 × 107*** | 7.4 × 105 | 3.07 × 10−2* | ||
| Device | 1 | 34,605 *** | 0.494 *** | 4.3 × 108*** | 3.2 × 106* | 1.0 × 108*** | 3.27 × 10−1*** | ||
| Genotypic set*ENV | 3 | 1519 | 0.008 | 1.4 × 107 | 9.8 × 105 | 8.7 × 106 | 4.43 × 10−3 | ||
| F1 vs Parents*ENV | 1 | 56 | 0.002 | 6.6 × 106 | 1.9 × 106 | 2.3 × 106 | 3.94 × 10−1*** | ||
| F1 vs Females*ENV | 1 | 57 | 0.000 | 2.5 × 107 | 2.5 × 106· | 1.1 × 107 | 3.00 × 10−4 | ||
| F1 vs Males*ENV | 1 | 153 | 0.003 | 2.0 × 105 | 6.2 × 105 | 7.0 × 103 | 5.47 × 10−6 | ||
| Females vs Males*ENV | 1 | 99 | 0.000 | 2.4 × 107 | 2.0 × 106 | 1.1 × 107 | 4.00 × 10−4 | ||
| F1 (FemA) vs F1 (FemB)*ENV | 1 | 4336 * | 0.021· | 1.6 × 107 | 6.9 × 102 | 1.5 × 107 | 4.09 × 10−5 | ||
| Genotypes within GS*ENV | 57 | 739 | 0.006 | 1.5 × 107 | 8.6 × 105· | 7.7 × 106· | 6.68 × 10−3 | ||
| Residuals | 412 | 768 | 0.007 | 1.2 × 107 | 6.6 × 105 | 6.1 × 106 | 5.48 × 10−3 |
| Environment Effect | Genotypic Effect | |||||
|---|---|---|---|---|---|---|
| Trait | Unstressed | Water-Stressed | Female | Hybrid | Male | |
| RC | 63.5 ± 0.82 | 43.0 ± 0.64 | *** | 56.0 ± 2.49a | 53.1 ± 1.38b | 52.4 ± 1.81b |
| LTD | −5.02 ± 0.35 | −3.58 ± 0.21 | *** | −3.40 ± 0.47a | −4.40 ± 0.28b | −4.47 ± 0.41b |
| φII | 0.270 ± 0.006 | 0.321 ± 0.006 | *** | 0.306 ± 0.016a | 0.290 ± 0.006b | 0.301 ± 0.009a |
| ΦNO | 0.270 ± 0.010 | 0.236 ± 0.008 | *** | 0.275 ± 0.017a | 0.248 ± 0.009b | 0.254 ± 0.011b |
| ΦNPQ | 0.461 ± 0.009 | 0.443 ± 0.013 | ns | 0.419 ± 0.019c | 0.462 ± 0.010a | 0.445 ± 0.013b |
| NPQt | 1.93 ± 0.10 | 2.27 ± 0.19 | ** | 1.70 ± 0.22b | 2.22 ± 0.16a | 2.02 ± 0.18ab |
| qL | 0.226 ± 0.009 | 0.295 ± 0.008 | *** | 0.246 ± 0.020a | 0.263 ± 0.009a | 0.262 ± 0.012a |
| qP | 0.442 ± 0.010 | 0.522 ± 0.006 | *** | 0.483 ± 0.023a | 0.480 ± 0.009a | 0.490 ± 0.012a |
| LEF | 168.97 ± 3.23 | 192.51 ± 4.93 | *** | 172.2 ± 8.9b | 180.7 ± 4.1a | 184.3 ± 5.6a |
| RFd | 0.375 ± 0.011 | 0.481 ± 0.014 | *** | 0.454 ± 0.034a | 0.417 ± 0.013b | 0.439 ± 0.017a |
| Fm′ | 11657 ± 421 | 12656 ± 457 | ** | 12635 ± 1025a | 11930 ± 413a | 12406 ± 537a |
| F0′ | 4139 ± 108 | 4576 ± 93 | *** | 4247 ± 266a | 4352 ± 97a | 4413 ± 120a |
| Fs | 8541 ± 334 | 8457 ± 269 | ns | 8684 ± 686a | 8407 ± 288a | 8605 ± 357a |
| Fv′/Fm′ | 0.631 ± 0.008 | 0.617 ± 0.011 | * | 0.651 ± 0.016a | 0.616 ± 0.009b | 0.628 ± 0.011b |
| n | 268 | 268 | 64 | 312 | 160 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Calleja, M.; Monteagudo, A.; Casas, A.M.; Boutin, C.; Pin, P.A.; Morales, F.; Igartua, E. Rapid On-Site Phenotyping via Field Fluorimeter Detects Differences in Photosynthetic Performance in a Hybrid—Parent Barley Germplasm Set. Sensors 2020, 20, 1486. https://doi.org/10.3390/s20051486
Fernández-Calleja M, Monteagudo A, Casas AM, Boutin C, Pin PA, Morales F, Igartua E. Rapid On-Site Phenotyping via Field Fluorimeter Detects Differences in Photosynthetic Performance in a Hybrid—Parent Barley Germplasm Set. Sensors. 2020; 20(5):1486. https://doi.org/10.3390/s20051486
Chicago/Turabian StyleFernández-Calleja, Miriam, Arantxa Monteagudo, Ana M. Casas, Christophe Boutin, Pierre A. Pin, Fermín Morales, and Ernesto Igartua. 2020. "Rapid On-Site Phenotyping via Field Fluorimeter Detects Differences in Photosynthetic Performance in a Hybrid—Parent Barley Germplasm Set" Sensors 20, no. 5: 1486. https://doi.org/10.3390/s20051486
APA StyleFernández-Calleja, M., Monteagudo, A., Casas, A. M., Boutin, C., Pin, P. A., Morales, F., & Igartua, E. (2020). Rapid On-Site Phenotyping via Field Fluorimeter Detects Differences in Photosynthetic Performance in a Hybrid—Parent Barley Germplasm Set. Sensors, 20(5), 1486. https://doi.org/10.3390/s20051486