Effect of Lactamase Inhibitors on the Biosensor Penp during the Measurement of Lactam Antibiotics Concentration


 and
and
Abstract
1. Introduction
- (1)
- First, the biological recognition domain of PenP interacts with LA and a dynamic equilibrium occurs in time that is a function of LA concentration.
- (2)
- Next, a change in the intrinsic fluorescence level of PenP occurs because a conformational change occurs at the biological recognition domain of PenP associated with LA.
2. Materials and Methods
2.1. Antibiotics
2.2. Fluorescence Kinetics
2.3. Treatment of the Data
2.4. Stats
3. Results
3.1. Fluorescence Pattern Induced on PenP Exposed to Different LA and Its Relation with the Lactamase Resistance
3.2. Fluorescence Pattern Induced on PenP Exposed to Different LI
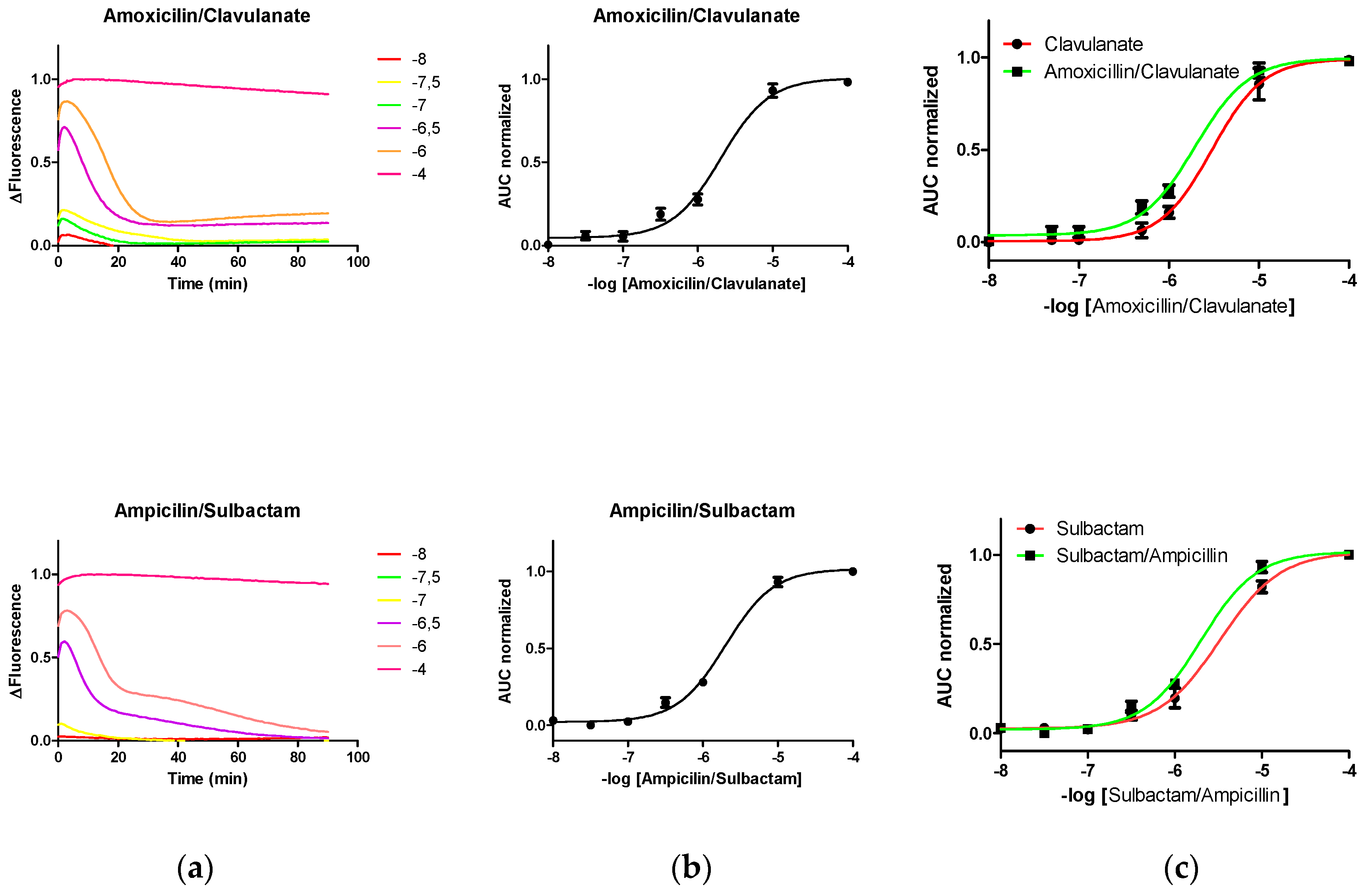
3.3. Fluorescence Pattern Induced on PenP Exposed to Different Commercial Compositions Comprising LA and LI
4. Discussion
5. Conclusions
- (a)
- The mixtures of LI with LA are common because potentiate the therapeutic effects of the antibiotic,
- (b)
- The mechanism of interaction among LI/lactamase is irreversible, and
- (c)
- The biological recognition domain of PenP is a lactamase mutant.
Author Contributions
Funding
Conflicts of Interest
References
- Chan, P.-H.; Liu, H.-B.; Chen, Y.W.; Chan, K.-C.; Tsang, C.-W.; Leung, Y.-C.; Wong, K.-Y. Rational design of a novel fluorescent biosensor for beta-lactam antibiotics from a class A beta-lactamase. J. Am. Chem. Soc. 2004, 126, 4074–4075. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.C.; Robinson, C.V.; Aplin, R.T.; Waley, S.G. Site-directed mutagenesis of beta-lactamase I: Role of Glu-166. Biochem. J. 1994, 299 Pt 3, 671–678. [Google Scholar] [CrossRef]
- Petrosino, J.F.; Palzkill, T. Systematic mutagenesis of the active site omega loop of TEM-1 beta-lactamase. J. Bacteriol. 1996, 178, 1821–1828. [Google Scholar] [CrossRef]
- Tsang, M.-W.; Chan, P.-H.; Liu, S.-Y.; Wong, K.-Y.; Leung, Y.-C. A fluorescein-labeled AmpC β-lactamase allows rapid characterization of β-lactamase inhibitors by real-time fluorescence monitoring of the β-lactamase-inhibitor interactions. Biotechnol. J. 2016, 11, 257–265. [Google Scholar] [CrossRef]
- Chan, P.-H.; Chan, K.-C.; Liu, H.-B.; Chung, W.-H.; Leung, Y.-C.; Wong, K.-Y. Fluorescein-labeled beta-lactamase mutant for high-throughput screening of bacterial beta-lactamases against beta-lactam antibiotics. Anal. Chem. 2005, 77, 5268–5276. [Google Scholar] [CrossRef]
- Andresen, M.; Wong, K.Y.; Leung, Y.C.; Wong, W.T.; Chan, P.H.; Andresen-Vasquez, M.; Alegria, L.; Silva, C.; Tapia, P.; Downey, P.; et al. Method Based on the β-Lactamase PenPC Fluorescent Labeled for β-Lactam Antibiotic Quantification in Human Plasma. BioMed Res. Int. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Drawz, S.M.; Bonomo, R.A. Three Decades of -Lactamase Inhibitors. Clin. Microbiol. Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [PubMed]
- Palzkill, T.; Botstein, D. Identification of amino acid substitutions that alter the substrate specificity of TEM-1 beta-lactamase. J. Bacteriol. 1992, 174, 5237–5243. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Taracila, M.A.; Smith, K.M.; Xu, Y.; Bonomo, R.A. Understanding the molecular determinants of substrate and inhibitor specificities in the Carbapenemase KPC-2: Exploring the roles of Arg220 and Glu276. Antimicrob. Agents Chemother. 2012, 56, 4428–4438. [Google Scholar] [CrossRef]
- Fisher, J.F.; Mobashery, S. Three decades of the class A beta-lactamase acyl-enzyme. Curr. Protein Pept. Sci. 2009, 10, 401–407. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| LA (Lactam Antibiotics) | Structure | LI (Lactamase Inhibitors) | Structure |
|---|---|---|---|
| Meropenem |  | Clavulanate |  |
| Cefazolin |  | Sulbactam |  |
| Amoxicillin |  | ||
| Ampicillin |  | ||
| Lactam Ring |  |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, D.; Silva, C.; Ugalde, C.; Wong, K.-Y.; Leung, Y.-C.; So, L.-Y.; Andresen, M. Effect of Lactamase Inhibitors on the Biosensor Penp during the Measurement of Lactam Antibiotics Concentration. Sensors 2019, 19, 1237. https://doi.org/10.3390/s19051237
Soto D, Silva C, Ugalde C, Wong K-Y, Leung Y-C, So L-Y, Andresen M. Effect of Lactamase Inhibitors on the Biosensor Penp during the Measurement of Lactam Antibiotics Concentration. Sensors. 2019; 19(5):1237. https://doi.org/10.3390/s19051237
Chicago/Turabian StyleSoto, Dagoberto, Camila Silva, Cristian Ugalde, Kwok-Yin Wong, Yun-Chung Leung, Lok-Yan So, and Max Andresen. 2019. "Effect of Lactamase Inhibitors on the Biosensor Penp during the Measurement of Lactam Antibiotics Concentration" Sensors 19, no. 5: 1237. https://doi.org/10.3390/s19051237
APA StyleSoto, D., Silva, C., Ugalde, C., Wong, K.-Y., Leung, Y.-C., So, L.-Y., & Andresen, M. (2019). Effect of Lactamase Inhibitors on the Biosensor Penp during the Measurement of Lactam Antibiotics Concentration. Sensors, 19(5), 1237. https://doi.org/10.3390/s19051237