Toward an Embodied Medicine: A Portable Device with Programmable Interoceptive Stimulation for Heart Rate Variability Enhancement





Abstract
1. Introduction
2. Methods
2.1. Technical Development
2.2. Participants
2.3. Procedure
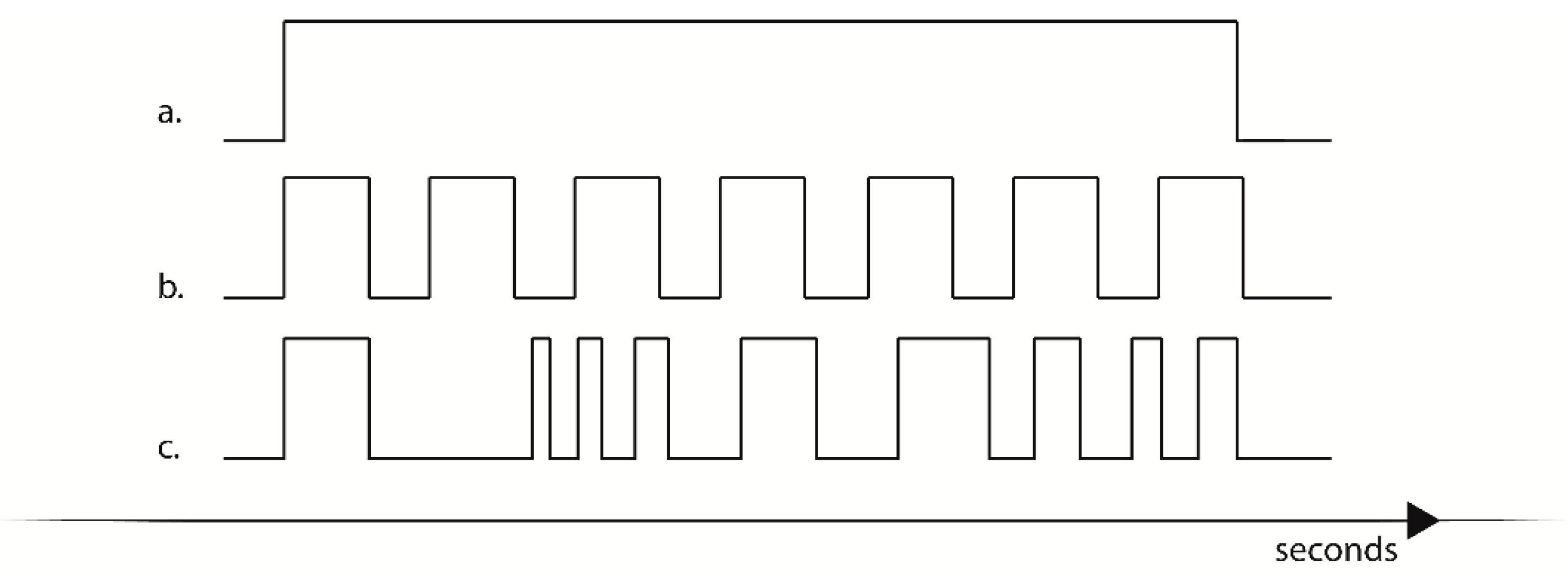
2.4. Experimental Design
2.5. Time and Frequency Domain Measurements of Heart Rate Variability
2.6. Statistical Analyses
3. Results
4. Discussion
5. Limitations
Author Contributions
Funding
Conflicts of Interest
References
- Riva, G.; Serino, S.; Di Lernia, D.; Pavone, E.F.; Dakanalis, A. Embodied Medicine: Mens Sana in Corpore Virtuale Sano. Front. Hum. Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Riva, G. The neuroscience of body memory: From the self through the space to the others. Cortex 2017, 104, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. Interoception: The sense of the physiological condition of the body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Cervero, F.; Janig, W. Visceral nociceptors: A new world order? Trends Neurosci. 1992, 15, 374–378. [Google Scholar] [CrossRef]
- Mense, S.; Meyer, H. Different types of slowly conducting afferent units in cat skeletal muscle and tendon. J. Physiol. 1985, 363, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.B.; Andrew, D.; Craig, A.D. Activation of spinobulbar lamina I neurons by static muscle contraction. J. Neurophysiol. 2002, 87, 1641–1645. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. How do you feel? Interoception: The sense of the physiological condition of the body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Iggo, A. Cutaneous mechanoreceptors with afferent C fibres. J. Physiol. 1960, 152, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Gordon, I.; Voos, A.C.; Bennett, R.H.; Bolling, D.Z.; Pelphrey, K.A.; Kaiser, M.D. Brain mechanisms for processing affective touch. Hum. Brain Mapp. 2013, 34, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. Emotional moments across time: A possible neural basis for time perception in the anterior insula. Philos. Trans. R. Soc. Lond. 2009, 364, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Di Lernia, D.; Serino, S.; Riva, G. Pain in the body. Altered interoception in chronic pain conditions: A systematic review. Neurosci. Biobehav. Rev. 2016, 71, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, S.; Piervincenzi, C.; Beomonte Zobel, B.; Romana Montecchi, F.; Riva, G.; Carducci, F.; Quattrocchi, C.C. Altered resting state functional connectivity of anterior cingulate cortex in drug naive adolescents at the earliest stages of anorexia nervosa. Sci. Rep. 2015, 5, 10818. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, S.; Quattrocchi, C.C.; Piervincenzi, C.; Zobel, B.B.; Montecchi, F.R.; Dakanalis, A.; Riva, G.; Carducci, F. White matter abnormalities in treatment-naive adolescents at the earliest stages of Anorexia Nervosa: A diffusion tensor imaging study. Psychiatry Res. 2017, 266, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Kerr, K.L.; Moseman, S.E.; Avery, J.A.; Bodurka, J.; Zucker, N.L.; Simmons, W.K. Altered Insula Activity during Visceral Interoception in Weight-Restored Patients with Anorexia Nervosa. Neuropsychopharmacology 2016, 41, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Wierenga, C.E.; Bischoff-Grethe, A.; Melrose, A.J.; Irvine, Z.; Torres, L.; Bailer, U.F.; Simmons, A.; Fudge, J.L.; McClure, S.M.; Ely, A.; et al. Hunger does not motivate reward in women remitted from anorexia nervosa. Biol. Psychiatry 2015, 77, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Dunn, B.D.; Stefanovitch, I.; Evans, D.; Oliver, C.; Hawkins, A.; Dalgleish, T. Can you feel the beat? Interoceptive awareness is an interactive function of anxiety- and depression-specific symptom dimensions. Behav. Res. Ther. 2010, 48, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Pollatos, O.; Traut-Mattausch, E.; Schandry, R. Differential effects of anxiety and depression on interoceptive accuracy. Depression Anxiety 2009, 26, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Sliz, D.; Hayley, S. Major depressive disorder and alterations in insular cortical activity: A review of current functional magnetic imaging research. Front. Hum. Neurosci. 2012, 6, 323. [Google Scholar] [CrossRef] [PubMed]
- Sprengelmeyer, R.; Steele, J.D.; Mwangi, B.; Kumar, P.; Christmas, D.; Milders, M.; Matthews, K. The insular cortex and the neuroanatomy of major depression. J. Affect. Disord. 2011, 133, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Stephan, K.E.; Manjaly, Z.M.; Mathys, C.D.; Weber, L.A.; Paliwal, S.; Gard, T.; Tittgemeyer, M.; Fleming, S.M.; Haker, H.; Seth, A.K.; et al. Allostatic Self-efficacy: A Metacognitive Theory of Dyshomeostasis-Induced Fatigue and Depression. Front. Hum. Neurosci. 2016, 10, 550. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, M.; Konrad, C.; Kugel, H.; Krug, A.; Schoning, S.; Ohrmann, P.; Uhlmann, C.; Postert, C.; Suslow, T.; Heindel, W.; et al. Insular and hippocampal gray matter volume reductions in patients with major depressive disorder. PLoS ONE 2014, 9, e102692. [Google Scholar] [CrossRef] [PubMed]
- Wiebking, C.; de Greck, M.; Duncan, N.W.; Tempelmann, C.; Bajbouj, M.; Northoff, G. Interoception in insula subregions as a possible state marker for depression-an exploratory fMRI study investigating healthy, depressed and remitted participants. Front. Behav. Neurosci. 2015, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, N.H.; Bechara, A. The hidden island of addiction: The insula. Trends Neurosci. 2009, 32, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Verdejo-Garcia, A.; Clark, L.; Dunn, B.D. The role of interoception in addiction: A critical review. Neurosci. Biobehav. Rev. 2012, 36, 1857–1869. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.C.; Shin, L.M. Functional neuroimaging studies of post-traumatic stress disorder. Expert Rev. Neurother. 2011, 11, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Chang, C.; Glover, G.H.; Gotlib, I.H. Increased insula coactivation with salience networks in insomnia. Biol. Psychol. 2014, 97, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.S.; Mitra, S. “I Do Not Exist”—Cotard Syndrome in Insular Cortex Atrophy. Biol. Psychiatry 2015, 77, 52–53. [Google Scholar] [CrossRef] [PubMed]
- Gorka, S.M.; Nelson, B.D.; Phan, K.L.; Shankman, S.A. Insula response to unpredictable and predictable aversiveness in individuals with panic disorder and comorbid depression. Biol. Mood Anxiety Disord. 2014, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Segerdahl, A.R.; Mezue, M.; Okell, T.W.; Farrar, J.T.; Tracey, I. The dorsal posterior insula subserves a fundamental role in human pain. Nat. Neurosci. 2015, 18, 499–500. [Google Scholar] [CrossRef] [PubMed]
- Starr, C.J.; Sawaki, L.; Wittenberg, G.F.; Burdette, J.H.; Oshiro, Y.; Quevedo, A.S.; Coghill, R.C. Roles of the insular cortex in the modulation of pain: Insights from brain lesions. J. Neurosci. 2009, 29, 2684–2694. [Google Scholar] [CrossRef] [PubMed]
- Olausson, H.; Wessberg, J.; McGlone, F. Affective Touch and the Neurophysiology of CT Afferents; Springer: Berlin, Germany, 2016. [Google Scholar]
- Olausson, H.; Lamarre, Y.; Backlund, H.; Morin, C.; Wallin, B.G.; Starck, G.; Ekholm, S.; Strigo, I.; Worsley, K.; Vallbo, A.B.; et al. Unmyelinated tactile afferents signal touch and project to insular cortex. Nat. Neurosci. 2002, 5, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Crucianelli, L.; Cardi, V.; Treasure, J.; Jenkinson, P.M.; Fotopoulou, A. The perception of affective touch in anorexia nervosa. Psychiatry Res. 2016, 239, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Habig, K.; Schanzer, A.; Schirner, W.; Lautenschlager, G.; Dassinger, B.; Olausson, H.; Birklein, F.; Gizewski, E.R.; Kramer, H.H. Low threshold unmyelinated mechanoafferents can modulate pain. BMC Neurol. 2017, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- Krahé, C.; Drabek, M.M.; Paloyelis, Y.; Fotopoulou, A. Affective touch and attachment style modulate pain: A laser-evoked potentials study. Philos. Trans. R. Soc. B 2016, 371, 20160009. [Google Scholar] [CrossRef] [PubMed]
- Liljencrantz, J.; Olausson, H. Tactile C fibers and their contributions to pleasant sensations and to tactile allodynia. Front. Behav. Neurosci. 2014, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Delfini, M.-C.; Mantilleri, A.; Gaillard, S.; Hao, J.; Reynders, A.; Malapert, P.; Alonso, S.; François, A.; Barrere, C.; Seal, R. TAFA4, a chemokine-like protein, modulates injury-induced mechanical and chemical pain hypersensitivity in mice. Cell Rep. 2013, 5, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Vrontou, S.; Wong, A.M.; Rau, K.K.; Koerber, H.R.; Anderson, D.J. Genetic identification of C fibres that detect massage-like stroking of hairy skin in vivo. Nature 2013, 493, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Perl, E.R. A specific inhibitory pathway between substantia gelatinosa neurons receiving direct C-fiber input. J. Neurosci. 2003, 23, 8752–8758. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, R.; Backlund Wasling, H.; Liljencrantz, J.; Olausson, H.; Johnson, R.D.; Wessberg, J. Human C-tactile afferents are tuned to the temperature of a skin-stroking caress. J. Neurosci. 2014, 34, 2879–2883. [Google Scholar] [CrossRef] [PubMed]
- Crucianelli, L.; Metcalf, N.K.; Fotopoulou, A.K.; Jenkinson, P.M. Bodily pleasure matters: Velocity of touch modulates body ownership during the rubber hand illusion. Front. Psychol. 2013, 4, 703. [Google Scholar] [CrossRef] [PubMed]
- Ogden, R.S.; Moore, D.; Redfern, L.; McGlone, F. The effect of pain and the anticipation of pain on temporal perception: A role for attention and arousal. Cogn. Emot. 2015, 29, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Ogden, R.S.; Moore, D.; Redfern, L.; McGlone, F. Stroke me for longer this touch feels too short: The effect of pleasant touch on temporal perception. Conscious. Cogn. 2015, 36, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Di Lernia, D.; Serino, S.; Pezzulo, G.; Pedroli, E.; Cipresso, P.; Riva, G. Feel the Time. Time Perception as a Function of Interoceptive Processing. Front. Hum. Neurosci. 2018, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Triscoli, C.; Croy, I.; Steudte-Schmiedgen, S.; Olausson, H.; Sailer, U. Heart rate variability is enhanced by long-lasting pleasant touch at CT-optimized velocity. Biol. Psychol. 2017, 128, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bigger, J.T.; Fleiss, J.L.; Steinman, R.C.; Rolnitzky, L.M.; Kleiger, R.E.; Rottman, J.N. Frequency domain measures of heart period variability and mortality after myocardial infarction. Circulation 1992, 85, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Karmakar, C.; Gray, R.; Jindal, R.; Lim, T.; Bryant, C. Heart rate variability alterations in late life depression: A meta-analysis. J. Affect. Disord. 2018, 235, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, F.; Ginsberg, J.P. An Overview of Heart Rate Variability Metrics and Norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef] [PubMed]
- Vallbo, A.B.; Olausson, H.; Wessberg, J. Unmyelinated afferents constitute a second system coding tactile stimuli of the human hairy skin. J. Neurophysiol. 1999, 81, 2753–2763. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, R.; Carlsson, I.; Wester, H.; Olausson, H.; Backlund Wasling, H. Touch perceptions across skin sites: Differences between sensitivity, direction discrimination and pleasantness. Front. Behav. Neurosci. 2014, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and affective touch: Sensing and feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef] [PubMed]
- Wessberg, J.; Olausson, H.; Fernstrom, K.W.; Vallbo, A.B. Receptive field properties of unmyelinated tactile afferents in the human skin. J. Neurophysiol. 2003, 89, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Vallbo, A.B.; Olausson, H.; Wessberg, J.; Kakuda, N. Receptive field characteristics of tactile units with myelinated afferents in hairy skin of human subjects. J. Physiol. 1995, 483 Pt 3, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Macefield, V.G. Tactile C Fibers. In Encyclopedia of Neuroscience; Springer: Berlin, Germany, 2009; pp. 3995–3998. [Google Scholar]
- Nordin, M. Low-threshold mechanoreceptive and nociceptive units with unmyelinated (C) fibres in the human supraorbital nerve. J. Physiol. 1990, 426, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Goldberger, J.J.; KIM, Y.H.; Ahmed, M.W.; Kadish, A.H. Effect of graded increases in parasympathetic tone on heart rate variability. J. Cardiovasc. Electrophysiol. 1996, 7, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Roudaut, Y.; Lonigro, A.; Coste, B.; Hao, J.; Delmas, P.; Crest, M. Touch sense: Functional organization and molecular determinants of mechanosensitive receptors. Channels 2012, 6, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, F.; McCraty, R.; Zerr, C.L. A healthy heart is not a metronome: An integrative review of the heart’s anatomy and heart rate variability. Front. Psychol. 2014, 5, 1040. [Google Scholar] [CrossRef] [PubMed]
- Variability, H.R. Standards of Measurement, Physiological Interpretation, and Clinical Use. Circulation 1996, 93, 1043–1065. [Google Scholar]
- Dwivedi, A.K.; Mallawaarachchi, I.; Alvarado, L.A. Analysis of small sample size studies using nonparametric bootstrap test with pooled resampling method. Stat. Med. 2017, 36, 2187–2205. [Google Scholar] [CrossRef] [PubMed]
- Nunan, D.; Sandercock, G.R.; Brodie, D.A. A quantitative systematic review of normal values for short-term heart rate variability in healthy adults. Pacing Clin. Electrophysiol. 2010, 33, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Ruxton, G.D.; Neuhäuser, M. Improving the reporting of p-values generated by randomization methods. Methods Ecol. Evol. 2013, 4, 1033–1036. [Google Scholar] [CrossRef]
- Bernardi, L.; Valle, F.; Coco, M.; Calciati, A.; Sleight, P. Physical activity influences heart rate variability and very-low-frequency components in Holter electrocardiograms. Cardiovasc. Res. 1996, 32, 234–237. [Google Scholar] [CrossRef]
- O’Reilly, J.X.; Schuffelgen, U.; Cuell, S.F.; Behrens, T.E.; Mars, R.B.; Rushworth, M.F. Dissociable effects of surprise and model update in parietal and anterior cingulate cortex. Proc. Natl. Acad. Sci. USA 2013, 110, 3660–3669. [Google Scholar] [CrossRef] [PubMed]
- Rosso, I.M.; Weiner, M.R.; Crowley, D.J.; Silveri, M.M.; Rauch, S.L.; Jensen, J.E. Insula and anterior cingulate GABA levels in posttraumatic stress disorder: Preliminary findings using magnetic resonance spectroscopy. Depression Anxiety 2014, 31, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Di Lernia, D.; Serino, S.; Cipresso, P.; Riva, G. Ghosts in the Machine. Interoceptive Modeling for Chronic Pain Treatment. Front. Neurosci. 2016, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Riva, G.; Villani, D.; Cipresso, P.; Repetto, C.; Triberti, S.; Di Lernia, D.; Chirico, A.; Serino, S.; Gaggioli, A. Positive and Transformative Technologies for Active Ageing. Stud. Health Technol. Inf. 2016, 220, 308–315. [Google Scholar]
- Castelnuovo, G.; Giusti, E.M.; Manzoni, G.M.; Saviola, D.; Gabrielli, S.; Lacerenza, M.; Pietrabissa, G.; Cattivelli, R.; Spatola, C.A.M.; Rossi, A.; et al. What is the role of the placebo effect for pain relief in neurorehabilitation? Clinical implications from the Italian Consensus Conference on Pain in Neurorehabilitation. Front. Neurol. 2018, 9, 310. [Google Scholar] [CrossRef] [PubMed]
- Zanier, E.R.; Zoerle, T.; Di Lernia, D.; Riva, G. Virtual Reality for Traumatic Brain Injury. Front. Neurol. 2018, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Serino, S.; Scarpina, F.; Dakanalis, A.; Keizer, A.; Pedroli, E.; Castelnuovo, G.; Chirico, A.; Catallo, V.; di Lernia, D.; Riva, G. The Role of Age on Multisensory Bodily Experience: An Experimental Study with a Virtual Reality Full-Body Illusion. Cyberpsychol. Behav. Soc. Netw. 2018, 21, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Grossman, P.; Kollai, M. Respiratory sinus arrhythmia, cardiac vagal tone, and respiration: Within- and between-individual relations. Psychophysiology 1993, 30, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Grossman, P.; Karemaker, J.; Wieling, W. Prediction of tonic parasympathetic cardiac control using respiratory sinus arrhythmia: The need for respiratory control. Psychophysiology 1991, 28, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Grossman, P.; Taylor, E.W. Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution and biobehavioral functions. Biol. Psychol. 2007, 74, 263–285. [Google Scholar] [CrossRef] [PubMed]
- Gąsior, J.S.; Sacha, J.; Jeleń, P.J.; Zieliński, J.; Przybylski, J. Heart Rate and Respiratory Rate Influence on Heart Rate Variability Repeatability: Effects of the Correction for the Prevailing Heart Rate. Front. Physiol. 2016, 7, 356. [Google Scholar] [CrossRef] [PubMed]
- Quintana, D.S.; Elstad, M.; Kaufmann, T.; Brandt, C.L.; Haatveit, B.; Haram, M.; Nerhus, M.; Westlye, L.T.; Andreassen, O.A. Resting-state high-frequency heart rate variability is related to respiratory frequency in individuals with severe mental illness but not healthy controls. Sci. Rep. 2016, 6, 37212. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| N | Min | Max | Mean | SD | |
|---|---|---|---|---|---|
| Age | 13 | 23 | 71 | 36.15 | 17.59 |
| BMI | 13 | 19.81 | 25.91 | 22.26 | 1.92 |
| SDNN | 13 | 28 | 69 | 49.54 | 11.20 |
| rMSSD | 13 | 12 | 69 | 40.46 | 14.39 |
| AVG_IHR | 13 | 66 | 91 | 74.23 | 6.82 |
| VLF | 13 | 140 | 2207 | 901.23 | 570.32 |
| LF | 13 | 86 | 3624 | 881.77 | 1035.95 |
| HF | 13 | 33 | 3436 | 726.46 | 904.83 |
| Baseline | Stimulation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| EXP (N = 7) | SHAM (N = 6) | p | EXP (N = 7) | SHAM (N = 6) | p | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| Age | 35.57 | 18.1 | 36.83 | 18.6 | 0.904 | << | << | << | << | << |
| BMI | 22.58 | 2.35 | 21.88 | 1.37 | 0.830 | << | << | << | << | << |
| SDNN | 48.29 | 11.10 | 51.00 | 12.18 | 0.774 | 57.00 | 12.98 | 50.50 | 10.65 | 0.352 |
| rMSSD | 41.00 | 16.94 | 39.83 | 12.31 | 0.886 | 51.57 | 14.44 | 36.83 | 8.88 | 0.036 * |
| AVG_IHR | 75.29 | 4.19 | 73.00 | 9.34 | 0.132 | 68.86 | 11.64 | 73.17 | 6.97 | 0.886 |
| VLF | 851.00 | 409.98 | 959.83 | 755.83 | 0.775 | 849.66 | 389.58 | 779.33 | 309.67 | 0.775 |
| LF | 809.00 | 791.02 | 966.67 | 1344.89 | 0.775 | 1045.71 | 473.39 | 802.83 | 408.15 | 0.475 |
| HF | 1041.29 | 1141.45 | 359.17 | 317.51 | 0.153 | 1164.86 | 602.69 | 536.17 | 527.43 | 0.037 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lernia, D.; Cipresso, P.; Pedroli, E.; Riva, G. Toward an Embodied Medicine: A Portable Device with Programmable Interoceptive Stimulation for Heart Rate Variability Enhancement. Sensors 2018, 18, 2469. https://doi.org/10.3390/s18082469
Di Lernia D, Cipresso P, Pedroli E, Riva G. Toward an Embodied Medicine: A Portable Device with Programmable Interoceptive Stimulation for Heart Rate Variability Enhancement. Sensors. 2018; 18(8):2469. https://doi.org/10.3390/s18082469
Chicago/Turabian StyleDi Lernia, Daniele, Pietro Cipresso, Elisa Pedroli, and Giuseppe Riva. 2018. "Toward an Embodied Medicine: A Portable Device with Programmable Interoceptive Stimulation for Heart Rate Variability Enhancement" Sensors 18, no. 8: 2469. https://doi.org/10.3390/s18082469
APA StyleDi Lernia, D., Cipresso, P., Pedroli, E., & Riva, G. (2018). Toward an Embodied Medicine: A Portable Device with Programmable Interoceptive Stimulation for Heart Rate Variability Enhancement. Sensors, 18(8), 2469. https://doi.org/10.3390/s18082469