Bioenergetic Health Assessment of a Single Caenorhabditis elegans from Postembryonic Development to Aging Stages via Monitoring Changes in the Oxygen Consumption Rate within a Microfluidic Device


Abstract
1. Introduction
2. Materials and Methods
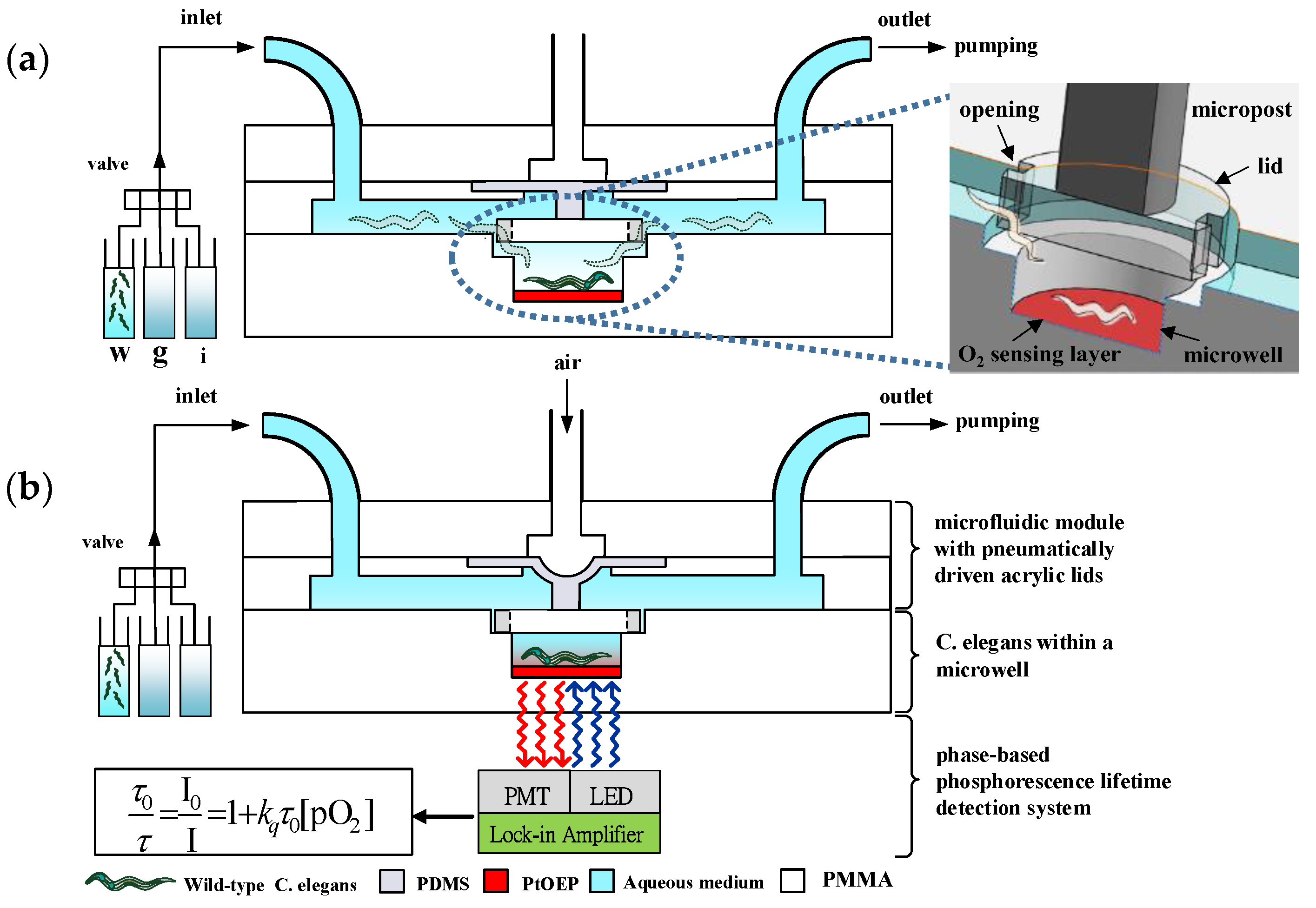
2.1. Principle of Operation
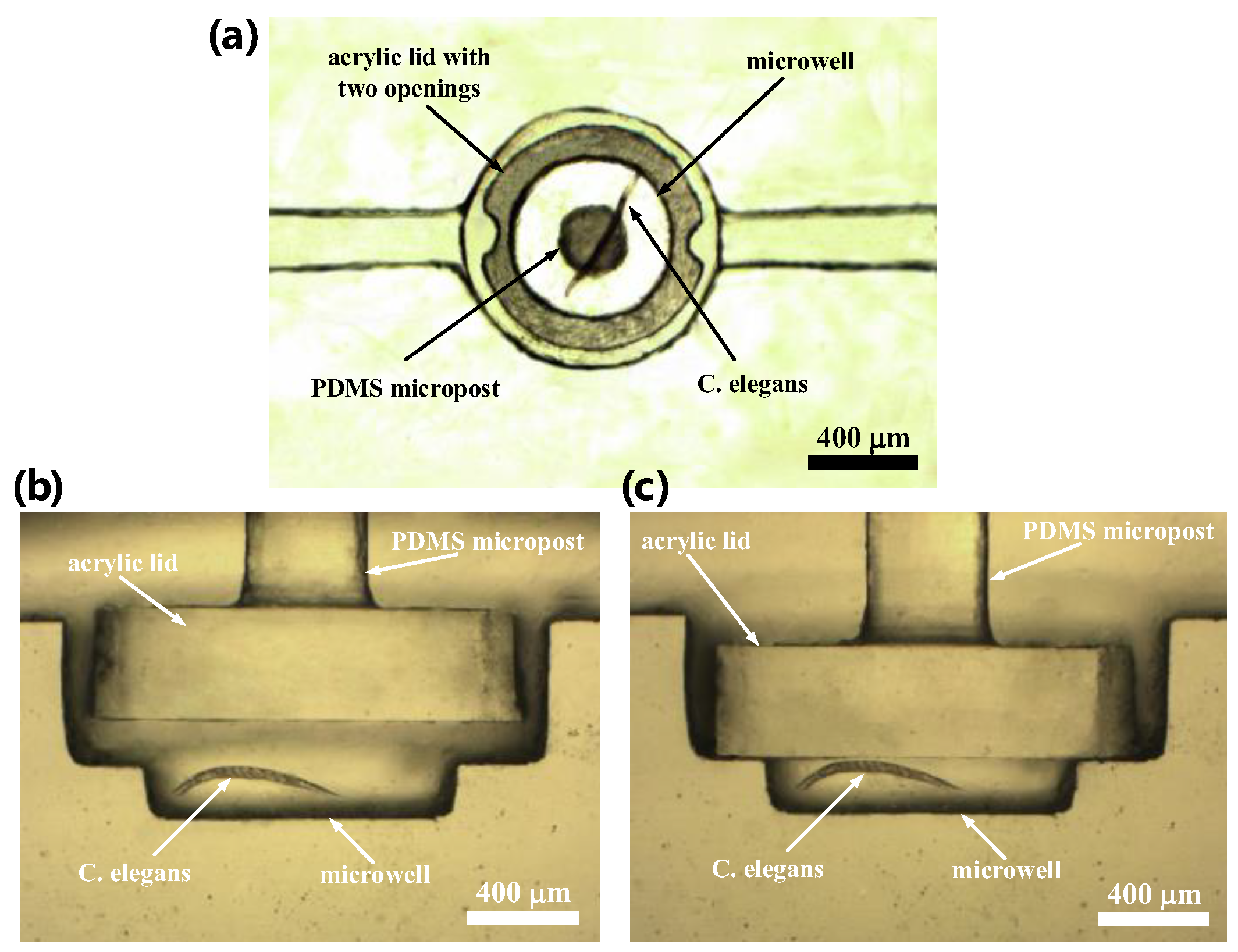
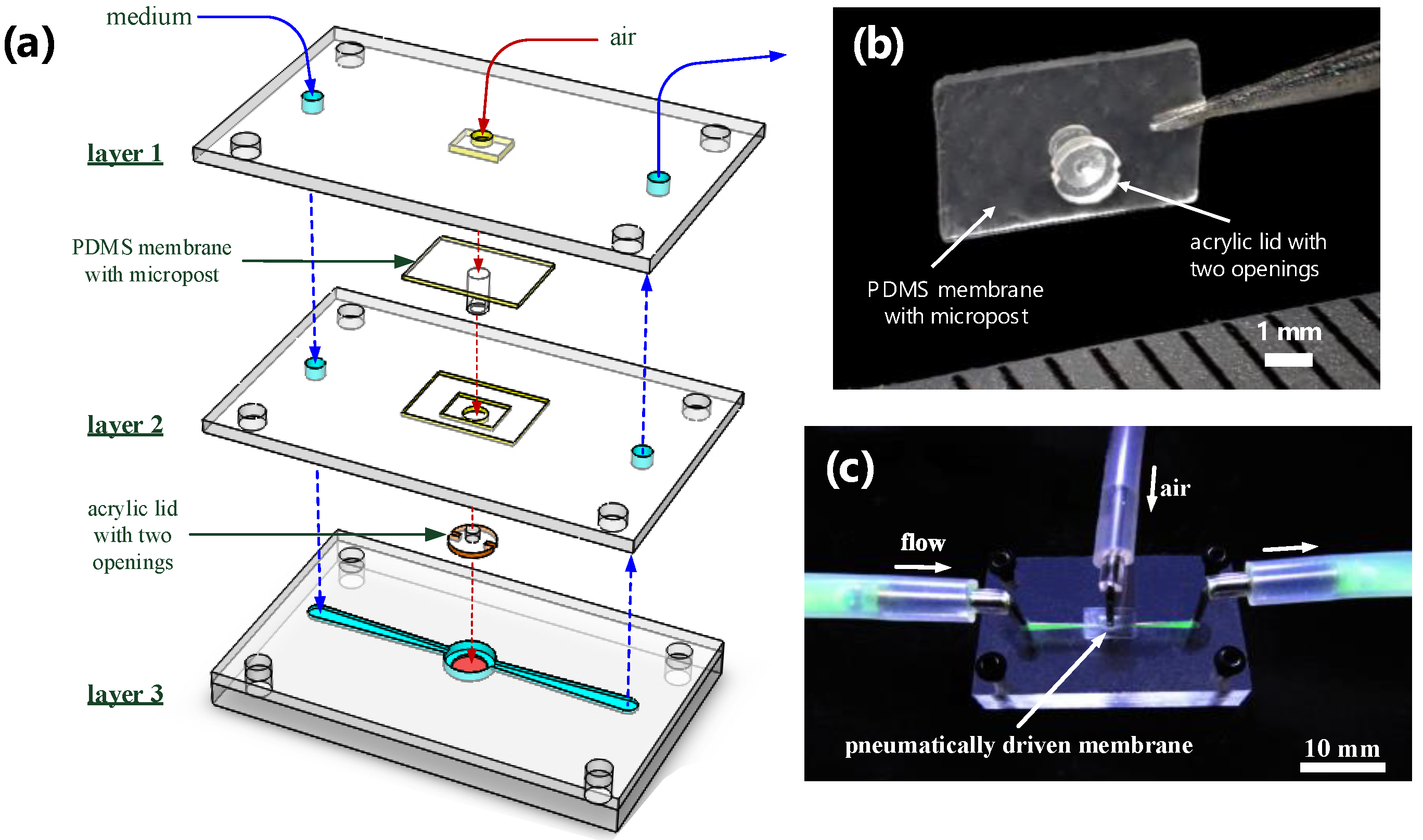
2.2. Fabrication of the Microfluidic Device
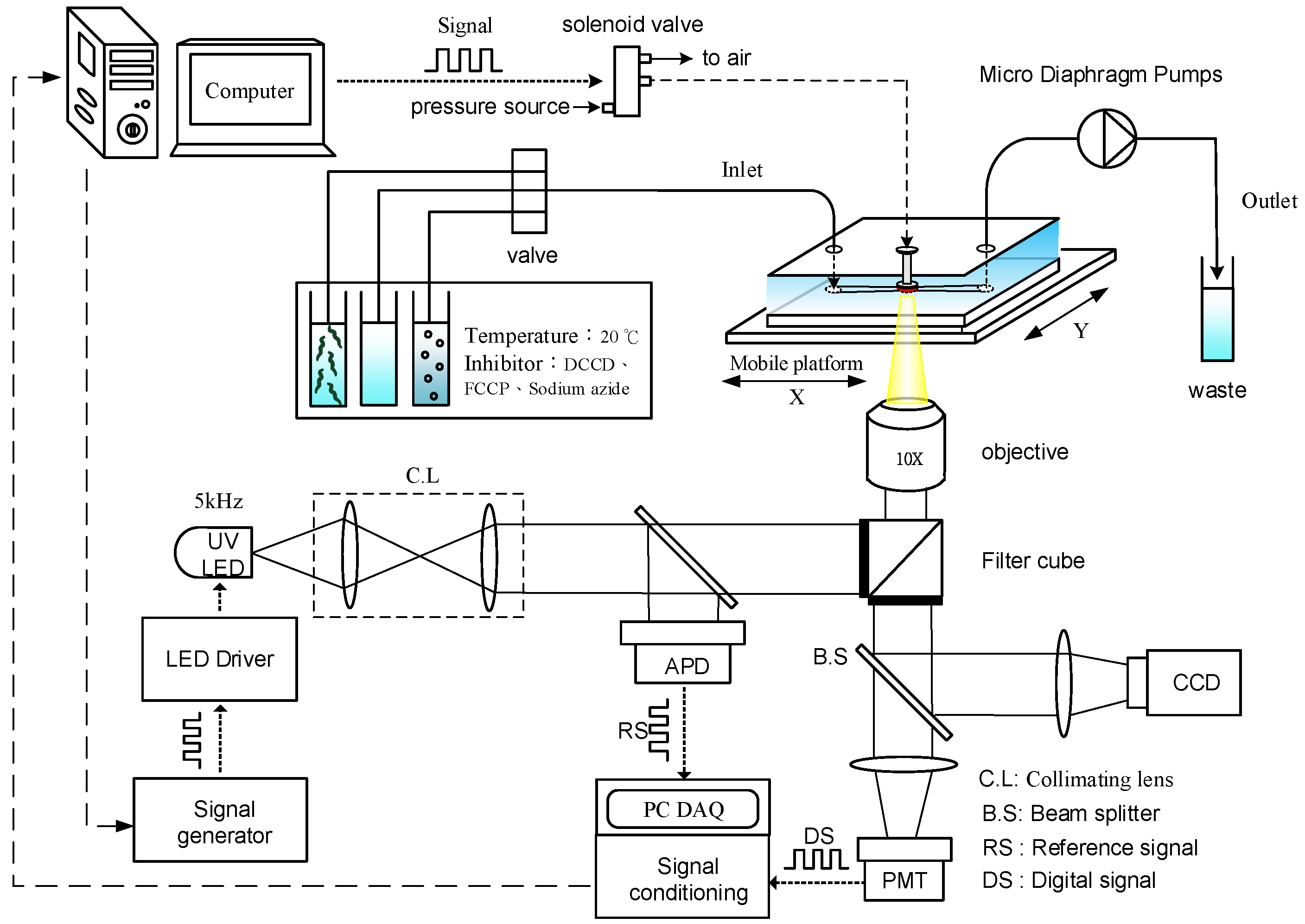
2.3. Optical Detection System for Oxygen Consumption Rate Measurement
2.4. Stern-Volmer Calibration Curve
2.5. Worm Culture and Metabolic Inhibitors
3. Results
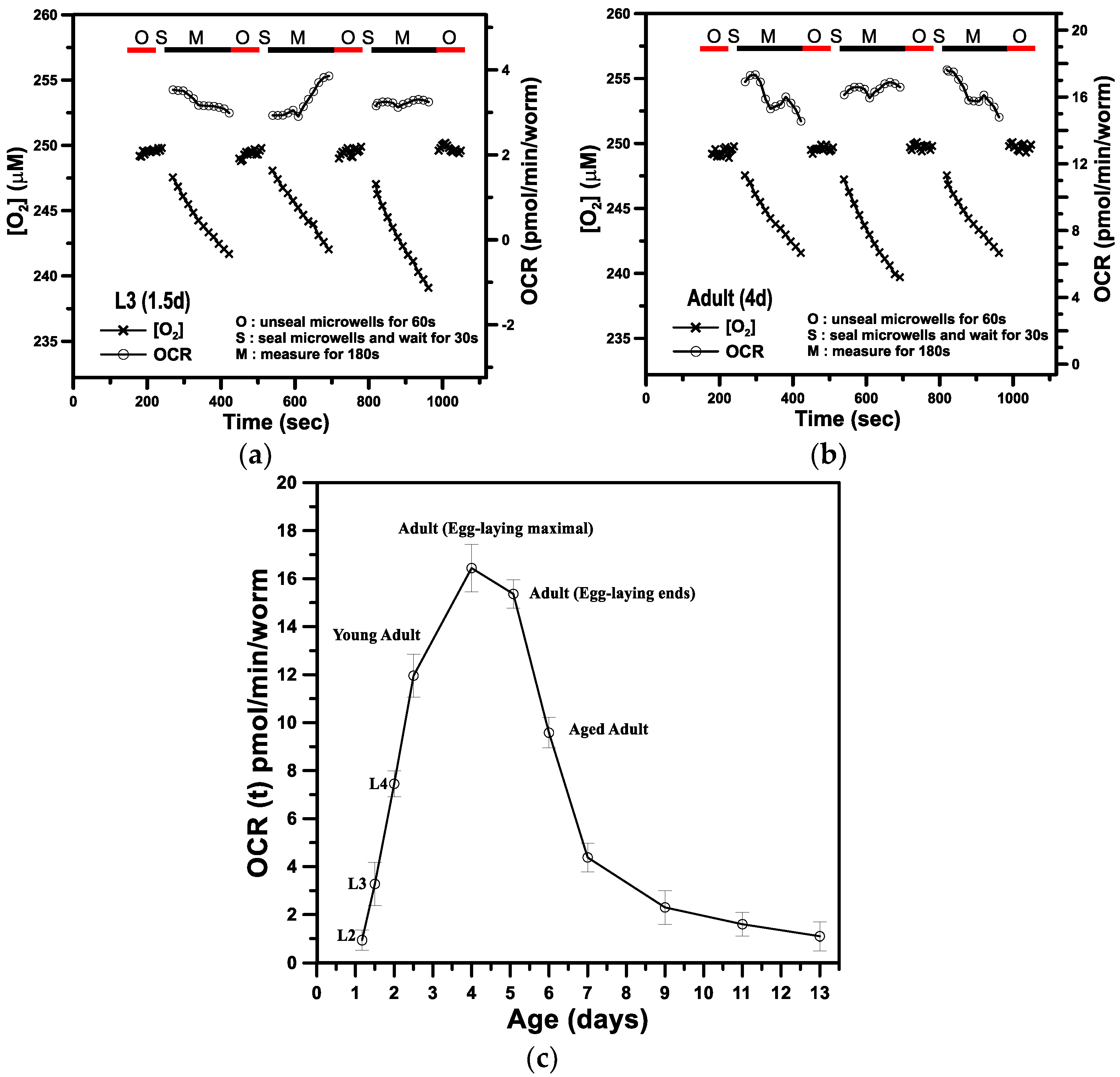
3.1. The Basal Oxygen Consumption Rate of a Single C. elegans within a Microwell
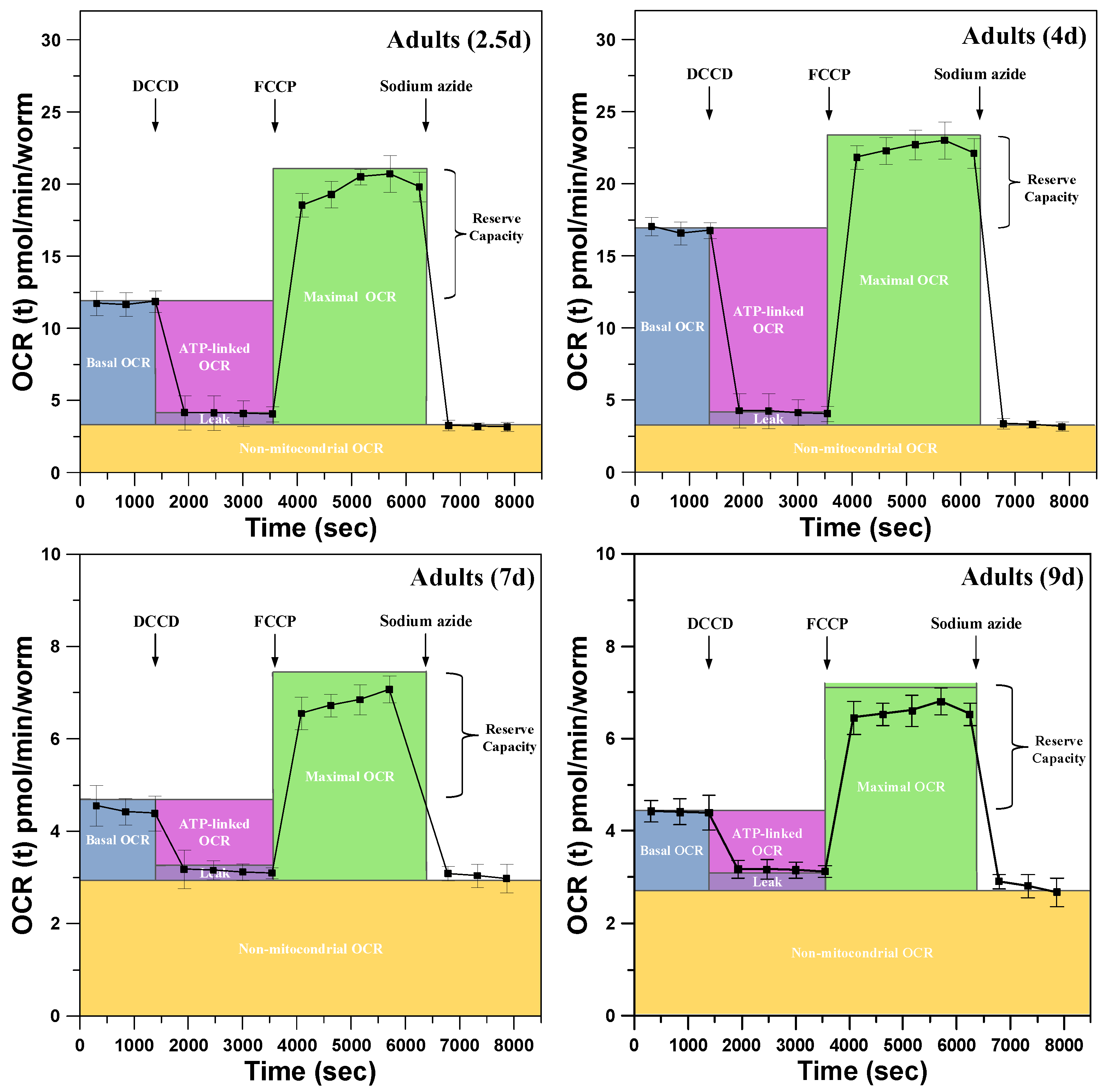
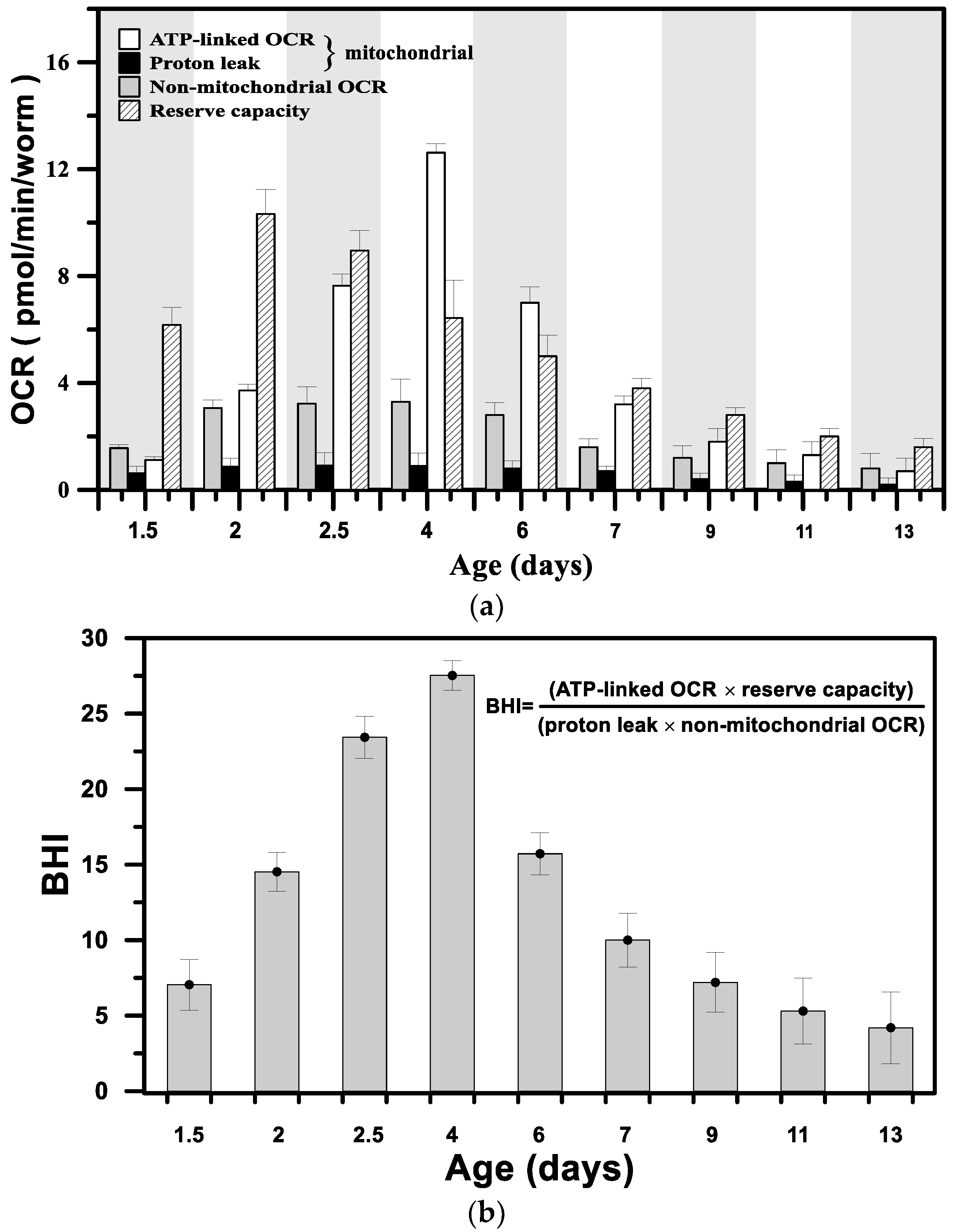
3.2. Metabolic Profiles and the Bioenergetic Health Index of a Single Developing C. elegans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wilson, D.F.; Harrison, D.K.; Vinogradov, S.A. Oxygen, ph, and mitochondrial oxidative phosphorylation. J. Appl. Physiol. 2012, 113, 1838–1845. [Google Scholar] [CrossRef] [PubMed]
- Suda, H.; Shouyama, T.; Yasuda, K.; Ishii, N. Direct measurement of oxygen consumption rate on the nematode caenorhabditis elegans by using an optical technique. Biochem. Biophys. Res. Commun. 2005, 330, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Stackley, K.D.; Beeson, C.C.; Rahn, J.J.; Chan, S.S.L. Bioenergetic profiling of zebrafish embryonic development. PLoS ONE 2011, 6, e25652. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Huang, K.-S.; Yu, C.-H.; Gong, H.-Y. Metabolic profile analysis of a single developing zebrafish embryo via monitoring of oxygen consumption rates within a microfluidic device. Biomicrofluidics 2013, 7, 064107. [Google Scholar] [CrossRef] [PubMed]
- Luz, A.L.; Rooney, J.P.; Kubik, L.L.; Gonzalez, C.P.; Song, D.H.; Meyer, J.N. Mitochondrial morphology and fundamental parameters of the mitochondrial respiratory chain are altered in caenorhabditis elegans strains deficient in mitochondrial dynamics and homeostasis processes. PLoS ONE 2015, 10, e0130940. [Google Scholar] [CrossRef] [PubMed]
- Koopman, M.; Michels, H.; Dancy, B.M.; Kamble, R.; Mouchiroud, L.; Auwerx, J.; Nollen, E.A.A.; Houtkooper, R.H. A screening-based platform for the assessment of cellular respiration in caenorhabditis elegans. Nat. Protoc. 2016, 11, 1798–1816. [Google Scholar] [CrossRef] [PubMed]
- Payne, B.A.I.; Chinnery, P.F. Mitochondrial dysfunction in aging: Much progress but many unresolved questions. BBA Bioenerg. 2015, 1847, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- McBride, H.M.; Neuspiel, M.; Wasiak, S. Mitochondria: More than just a powerhouse. Curr. Biol. 2006, 16, R551–R560. [Google Scholar] [CrossRef] [PubMed]
- Felix, M.A.; Braendle, C. The natural history of caenorhabditis elegans. Curr. Biol. 2010, 20, R965–R969. [Google Scholar] [CrossRef] [PubMed]
- Aubry, G.; Lu, H. A perspective on optical developments in microfluidic platforms for caenorhabditis elegans research. Biomicrofluidics 2014, 8, 011301. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.M.; Santra, T.S. Microfluidic devices in advanced caenorhabditis elegans research. Molecules 2016, 21, 1006. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.P.; Rezai, P. Microfluidic approaches for manipulating, imaging, and screening c. Elegans. Micromachines 2016, 7, 123. [Google Scholar] [CrossRef]
- Yang, J.P.; Chen, Z.G.; Ching, P.Y.; Shi, Q.J.; Li, X.C. An integrated microfluidic platform for evaluating in vivo antimicrobial activity of natural compounds using a whole-animal infection model. Lab Chip 2013, 13, 3373–3382. [Google Scholar] [CrossRef] [PubMed]
- Crane, M.M.; Chung, K.; Stirman, J.; Lu, H. Microfluidics-enabled phenotyping, imaging, and screening of multicellular organisms. Lab Chip 2010, 10, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Song, P.F.; Zhang, W.Z.; Sobolevski, A.; Bernard, K.; Hekimi, S.; Liu, X.Y. A microfluidic device for efficient chemical testing using caenorhabditis elegans. Biomed. Microdevices 2015, 17, 38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.B.; Li, Y.B.; He, Q.D.; Qin, J.; Yu, Y.Y.; Li, X.C.; Zhang, L.; Yao, M.C.; Liu, J.S.; Chen, Z.G. Microfluidic platform integrated with worm-counting setup for assessing manganese toxicity. Biomicrofluidics 2014, 8, 054110. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Huang, S.H. Metabolic profile analysis of a single caenorhabditis elegans across the lifespan within a microfluidic device. In Proceedings of the 2017 19th International Conference on Solid-State Sensors, Actuators and Microsystems (TRANSDUCERS), Kaohsiung, Taiwan, 18–22 June 2017; pp. 1660–1663. [Google Scholar]
- Ji, S.; Wu, W.; Wu, Y.; Zhao, T.; Zhou, F.; Yang, Y.; Zhang, X.; Liang, X.; Wu, W.; Chi, L.; et al. Real-time monitoring of luminescent lifetime changes of ptoep oxygen sensing film with led/photodiode-based time-domain lifetime device. Analyst 2009, 134, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Gruber, P.; Marques, M.P.C.; Szita, N.; Mayr, T. Integration and application of optical chemical sensors in microbioreactors. Lab Chip 2017, 17, 2693–2712. [Google Scholar] [CrossRef] [PubMed]
- Jahn, K.; Buschmann, V.; Hille, C. Simultaneous fluorescence and phosphorescence lifetime imaging microscopy in living cells. Sci. Rep. 2015, 5, 14334. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.C.; Kwon, M.S.; Jung, J.; Zeng, Y.Y.; Kim, M.; Chung, K.; Gierschner, J.; Youk, J.H.; Borisov, S.M.; Kim, J. Room-temperature-phosphorescence-based dissolved oxygen detection by core-shell polymer nanoparticles containing metal-free organic phosphors. Angew. Chem. Int. Ed. 2017, 56, 16207–16211. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bassett, W.P.; Askim, J.R.; Suslick, K.S. Differentiation among peroxide explosives with an optoelectronic nose. Chem. Commun. 2015, 51, 15312–15315. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Hsu, Y.-H.; Wu, C.-W.; Wu, C.-J. Light-addressable measurements of cellular oxygen consumption rates in microwell arrays based on phase-based phosphorescence lifetime detection. Biomicrofluidics 2012, 6, 44118. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Tsai, C.-H.; Wu, C.-W.; Wu, C.-J. Light-directed, spatially addressable oxygen detection in a hydrogel microarray based on phase-based lifetime detection using a digital micromirror device. Sens. Actuators A Phys. 2011, 165, 139–146. [Google Scholar] [CrossRef]
- Huang, S.H.; Huang, K.S.; Liou, Y.M. Simultaneous monitoring of oxygen consumption and acidification rates of a single zebrafish embryo during embryonic development within a microfluidic device. Microfluid Nanofluid 2017, 21, 3. [Google Scholar] [CrossRef]
- Meier, R.J.; Schreml, S.; Wang, X.D.; Landthaler, M.; Babilas, P.; Wolfbeis, O.S. Simultaneous photographing of oxygen and ph in vivo using sensor films. Angew. Chem. Int. Ed. 2011, 50, 10893–10896. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Lu, S.S.; Chen, Y.Y.; Zhao, T.T.; Jiang, Y.Q.; Wang, Y.R.; Chen, X. Simultaneous color sensing of o-2 and ph using a smartphone. Sens. Actuators B Chem. 2015, 220, 326–330. [Google Scholar] [CrossRef]
- Chu, C.S.; Syu, J.J. Optical sensor for dual sensing of oxygen and carbon dioxide based on sensing films coated on filter paper. Appl. Opt. 2017, 56, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. Genetics of caenorhabditis-elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Zhu, G.L.; Yin, F.C.; Wang, L.; Wei, W.B.; Jiang, L.; Qin, J.H. Modeling type 2 diabetes-like hyperglycemia in c-elegans on a microdevice. Integr. Biol. 2016, 8, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Khanna, N.; Cressman, C.P.; Tatara, C.P.; Williams, P.L. Tolerance of the nematode caenorhabditis elegans to ph, salinity, and hardness in aquatic media. Arch. Environ. Contam. Toxicol. 1997, 32, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, H.L.; Wang, Y.; Tian, Y.Q.; Zhao, J.P.; Li, Y. High sensitivity and accuracy dissolved oxygen (do) detection by using ptoep/poly(mma-co-tfema) sensing film. Spectrochim. Acta A 2017, 170, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Nock, V.; Blaikie, R.J.; David, T. Patterning, integration and characterisation of polymer optical oxygen sensors for microfluidic devices. Lab Chip 2008, 8, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.G.; Benavides, G.A.; Lancaster, J.R., Jr.; Ballinger, S.; Dell’Italia, L.; Jianhua, Z.; Darley-Usmar, V.M. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biol. Chem. 2012, 393, 1485–1512. [Google Scholar] [CrossRef] [PubMed]
- Chacko, B.K.; Kramer, P.A.; Ravi, S.; Benavides, G.A.; Mitchell, T.; Dranka, B.P.; Ferrick, D.; Singal, A.K.; Ballinger, S.W.; Bailey, S.M.; et al. The bioenergetic health index: A new concept in mitochondrial translational research. Clin. Sci. (Lond.) 2014, 127, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Chacko, B.K.; Zhi, D.; Darley-Usmar, V.M.; Mitchell, T. The bioenergetic health index is a sensitive measure of oxidative stress in human monocytes. Redox Biol. 2016, 8, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Beeson, C.C.; Beeson, G.C.; Schnellmann, R.G. A high-throughput respirometric assay for mitochondrial biogenesis and toxicity. Anal. Biochem. 2010, 404, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Facucho-Oliveira, J.M.; St. John, J.C. The relationship between pluripotency and mitochondrial DNA proliferation during early embryo development and embryonic stem cell differentiation. Stem Cell Rev. Rep. 2009, 5, 140–158. [Google Scholar] [CrossRef] [PubMed]
- Bratic, A.; Larsson, N.G. The role of mitochondria in aging. J. Clin. Investig. 2013, 123, 951–957. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Feature | Mean Age (day) | OCR (pmol/min/worm) |
|---|---|---|---|
| L1 | 558 somatic nuclei | 0.5 | NA |
| L2 | germ cell divisions continue | 1.2 | 0.94 ± 0.41 |
| L3 | forms spermathecal/uterus | 1.5 | 3.28 ± 0.89 |
| L4 | generate oocytes | 2 | 7.45 ± 0.55 |
| Young adults | 959 somatic nuclei | 2.5 | 11.96 ± 0.91 |
| Adults | egg-laying maximum | 4 | 16.44 ± 0.96 |
| Adults | egg-laying ends | 5 | 15.36 ± 0.59 |
| Aged adults | slow motion | 6 | 9.58 ± 0.64 |
| Aged adults | slow motion | 7 | 4.38 ± 0.41 |
| Aged adults | less motion | 9 | 2.33 ± 0.71 |
| Aged adults | less motion | 11 | 1.65 ± 0.52 |
| Aged adults | no motion | 13 | 1.12 ± 0.62 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-H.; Lin, Y.-W. Bioenergetic Health Assessment of a Single Caenorhabditis elegans from Postembryonic Development to Aging Stages via Monitoring Changes in the Oxygen Consumption Rate within a Microfluidic Device. Sensors 2018, 18, 2453. https://doi.org/10.3390/s18082453
Huang S-H, Lin Y-W. Bioenergetic Health Assessment of a Single Caenorhabditis elegans from Postembryonic Development to Aging Stages via Monitoring Changes in the Oxygen Consumption Rate within a Microfluidic Device. Sensors. 2018; 18(8):2453. https://doi.org/10.3390/s18082453
Chicago/Turabian StyleHuang, Shih-Hao, and Yu-Wei Lin. 2018. "Bioenergetic Health Assessment of a Single Caenorhabditis elegans from Postembryonic Development to Aging Stages via Monitoring Changes in the Oxygen Consumption Rate within a Microfluidic Device" Sensors 18, no. 8: 2453. https://doi.org/10.3390/s18082453
APA StyleHuang, S.-H., & Lin, Y.-W. (2018). Bioenergetic Health Assessment of a Single Caenorhabditis elegans from Postembryonic Development to Aging Stages via Monitoring Changes in the Oxygen Consumption Rate within a Microfluidic Device. Sensors, 18(8), 2453. https://doi.org/10.3390/s18082453