Design and Evaluation of Novel Polymyxin Fluorescent Probes

 , and
, and
Abstract
:1. Introduction
2. Methods
2.1. Chemical Reagents
2.2. HPLC Purification and LC-MS Analysis
2.3. Synthesis FADDI-285
2.4. Synthesis FADDI-286
2.5. Determination of MICs
3. Results
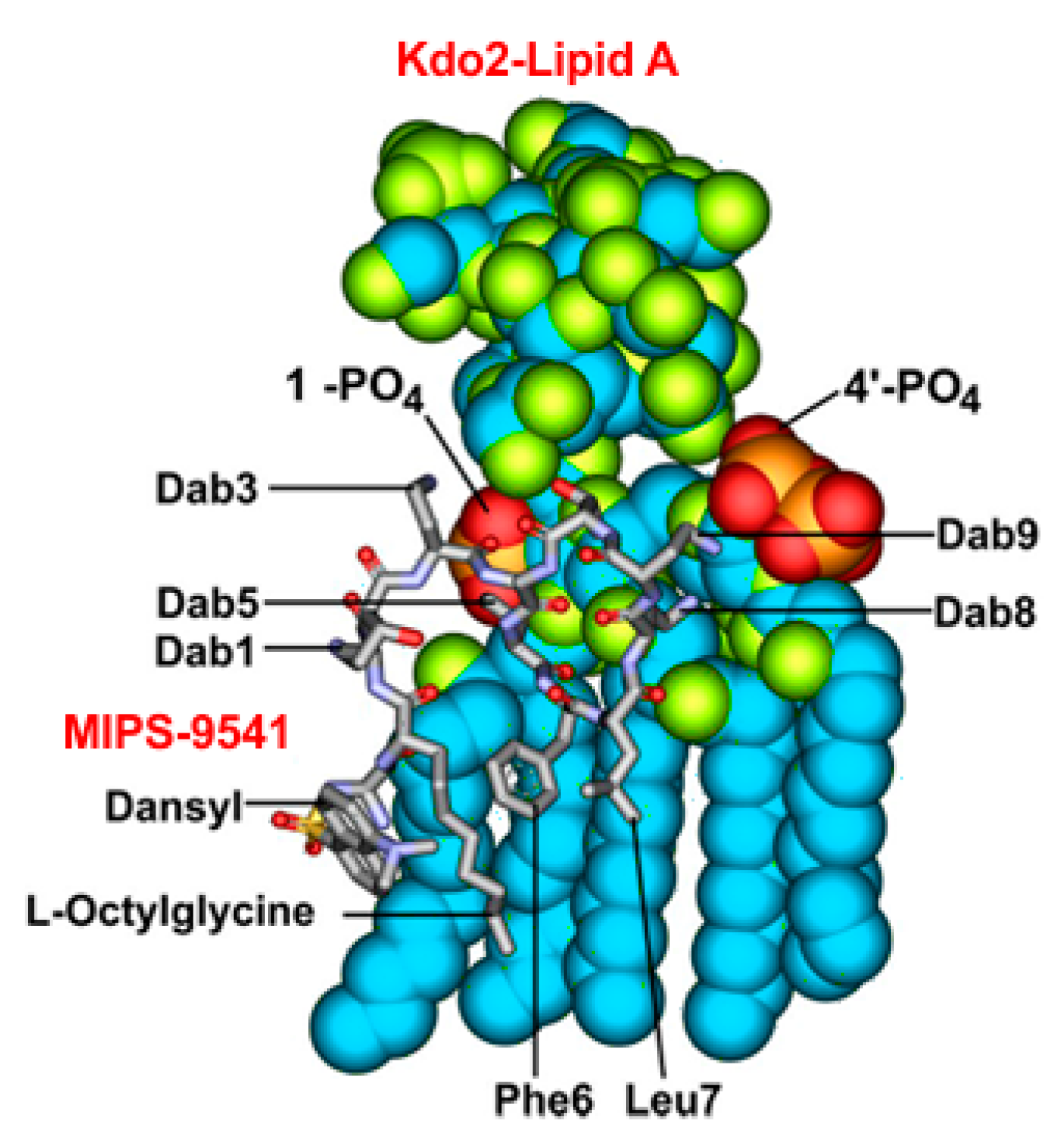
3.1. Probe Design and Synthesis
3.2. Antibacterial Activity of the Probes
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boucher, H.W.; Talbot, G.H.; Benjamin, D.K., Jr.; Bradley, J.; Guidos, R.J.; Jones, R.N.; Murray, B.E.; Bonomo, R.A.; Gilbert, D. 10 × ’20 Progress—Development of New Drugs Active Against Gram-Negative Bacilli: An Update From the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Roberts, K.D.; Nation, R.L.; Thompson, P.E.; Li, J. Pharmacology of polymyxins: New insights into an “old” class of antibiotics. Future Microbiol. 2013, 8, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Landman, D.; Georgescu, C.; Martin, D.A.; Quale, J. Polymyxins revisited. Clin. Microbiol. Rev. 2008, 21, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Akajagbor, D.S.; Wilson, S.L.; Shere-Wolfe, K.D.; Dakum, P.; Charurat, M.E.; Gilliam, B.L. Higher Incidence of Acute Kidney Injury With Intravenous Colistimethate Sodium Compared with Polymyxin B in Critically Ill Patients at a Tertiary Care Medical Center. Clin. Infect. Dis. 2013, 57, 1300–1303. [Google Scholar] [CrossRef] [PubMed]
- Hartzell, J.D.; Neff, R.; Ake, J.; Howard, R.; Olson, S.; Paolino, K.; Vishnepolsky, M.; Weintrob, A.; Wortmann, G. Nephrotoxicity Associated with Intravenous Colistin (Colistimethate Sodium) Treatment at a Tertiary Care Medical Center. Clin. Infect. Dis. 2009, 48, 1724–1728. [Google Scholar] [CrossRef] [PubMed]
- Kubin, C.J.; Ellman, T.M.; Phadke, V.; Haynes, L.J.; Calfee, D.P.; Yin, M.T. Incidence and predictors of acute kidney injury associated with intravenous polymyxin B therapy. J. Infect. 2012, 65, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Thompson, P.E.; Nation, R.L.; Li, J. Structure-activity relationships of polymyxin antibiotics. J. Med. Microbiol. 2010, 53, 1898–1916. [Google Scholar] [CrossRef] [PubMed]
- Ofek, I.; Cohen, S.; Rahmani, R.; Kabha, K.; Tamarkin, D.; Herzig, Y.; Rubinstein, E. Antibacterial synergism of polymyxin B nonapeptide and hydrophobic antibiotics in experimental gram-negative infections in mice. Antimicrob. Agents Chemother. 1994, 38, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Tsubery, H.; Ofek, I.; Cohen, S.; Fridkin, M. Structure-activity relationship study of polymyxin B nonapeptide. Adv. Exp. Med. Biol. 2000, 479, 219–222. [Google Scholar] [PubMed]
- Tsubery, H.; Ofek, I.; Cohen, S.; Fridkin, M. N-terminal modifications of polymyxin B nonapeptide and their effect on antibacterial activity. Peptides 2001, 22, 1675–1681. [Google Scholar] [CrossRef]
- Deris, Z.Z.; Swarbrick, J.D.; Roberts, K.D.; Azad, M.A.K.; Akter, J.; Horne, A.S.; Nation, R.L.; Rogers, K.L.; Thompson, P.E.; Velkov, T.; Li, J. Probing the Penetration of Antimicrobial Polymyxin Lipopeptides into Gram-Negative Bacteria. Bioconj. Chem. 2014, 25, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Azad, M.A.K.; Nowell, C.J.; Nation, R.L.; Thompson, P.E.; Roberts, K.D.; Velkov, T.; Li, J. Cellular Uptake and Localization of Polymyxins in Renal Tubular Cells Using Rationally Designed Fluorescent Probes. Antimicrob. Agents Chemother. 2015, 59, 7489–7496. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Roberts, K.D.; Nation, R.L.; Wang, J.; Thompson, P.E.; Li, J. Teaching ‘Old’ Polymyxins New Tricks: New-Generation Lipopeptides Targeting Gram-Negative ‘Superbugs’. ACS Chem. Biol. 2014, 9, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, K.; Sato, Y.; Ohki, K.; Okimura, K.; Uchida, Y.; Shindo, M.; Sakura, N. Contribution of Each Amino Acid Residue in Polymyxin B3 to Antimicrobial and Lipopolysaccharide Binding Activity. Chem. Pharm. Bull. 2009, 57, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Nation, R.L.; Velkov, T.; Li, J. Colistin and Polymyxin B: Peas in a Pod, or Chalk and Cheese? Clin. Infect. Dis. 2014, 59, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Pristovsek, P.; Kidric, J. The search for molecular determinants of LPS inhibition by proteins and peptides. Curr. Top. Med. Chem. 2004, 4, 1185–1201. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R. The bacterial outer membrane as a drug barrier. Trends Microbiol. 1997, 5, 37–42. [Google Scholar] [CrossRef]
- Hancock, R.E.; Lehrer, R. Cationic peptides: A new source of antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Clausell, A.; Garcia-Subirats, M.; Pujol, M.; Busquets, M.A.; Rabanal, F.; Cajal, Y. Gram-negative outer and inner membrane models: Insertion of cyclic cationic lipopeptides. J. Phys. Chem. B 2007, 111, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.P.; Hancock, R.E. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
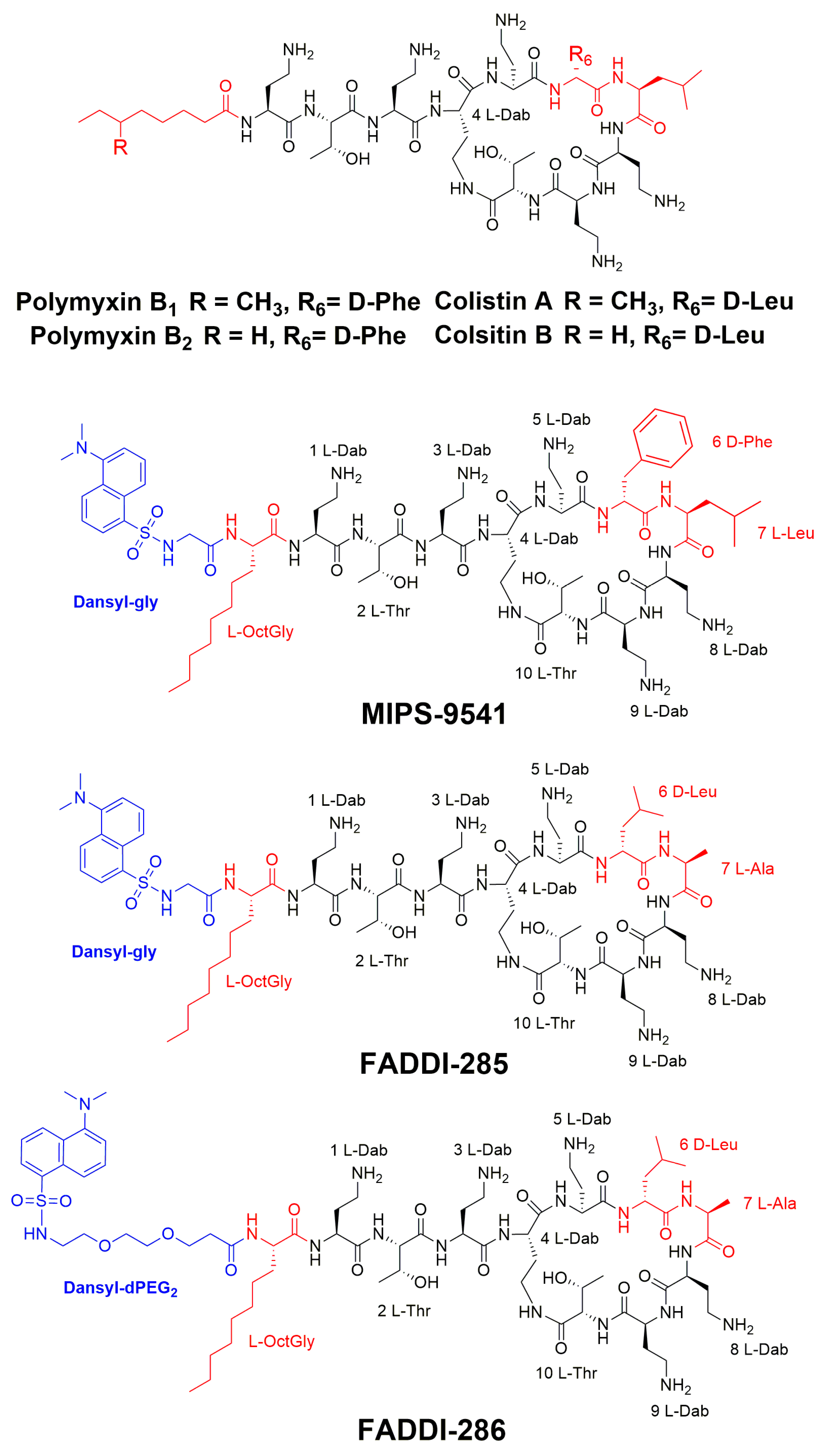

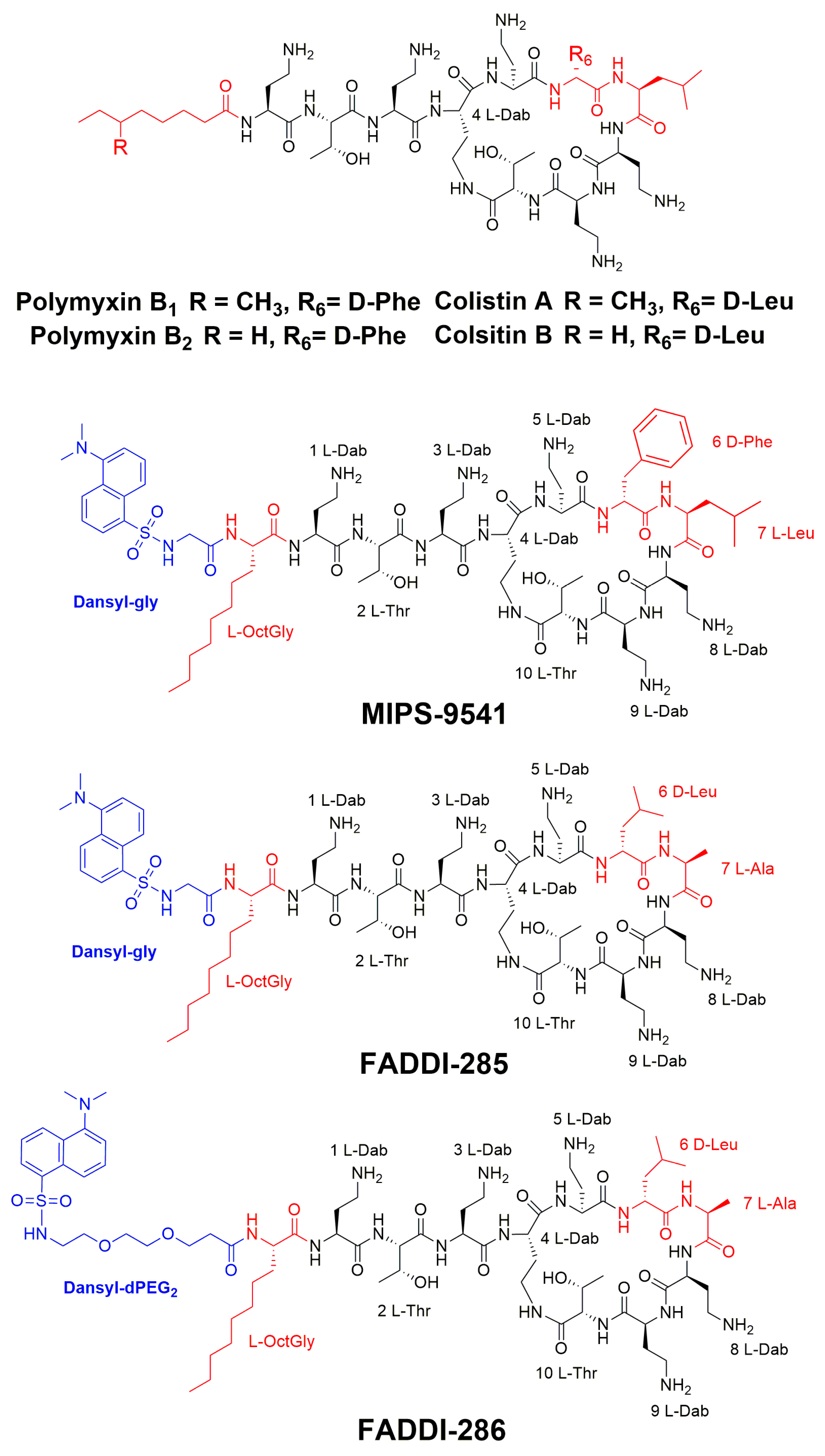{kind=link}
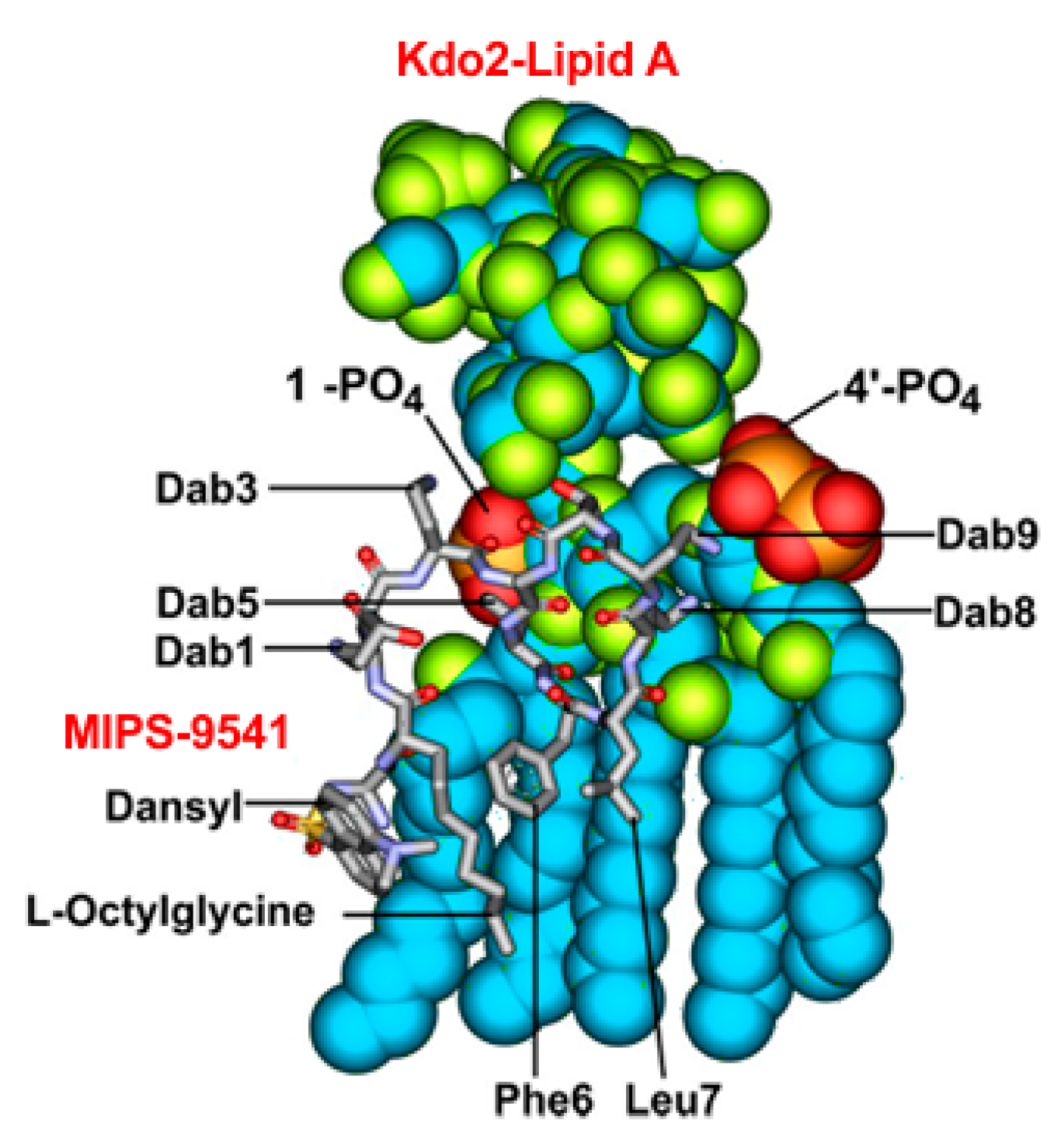{kind=link}
| Pseudomonas aeruginosa ATCC 27853 | Pseudomonas aeruginosa FADDI-PA022 | Pseudomonas aeruginosa FADDI-PA025 | Pseudomonas aeruginosa FADDI-PA070 | Pseudomonas aeruginosa FADDI-PA060 | Pseudomonas aeruginosa FADI-PA090 | Acinetobacter baumannii ATCC 19606 | Acinetobacter baumannii FADDI-AB034 | Acinetobacter baumannii ATCC 17978 | Acinetobacter baumannii ATCC 19606 Col 10 | Acinetobacter baumannii FADDI-AB156 | Acinetobacter baumannii FADDI-AB167 | Klebsiella pneumoniae ATCC 13883 | Klebsiella pneumoniae FADDI-KP027 | Klebsiella pneumoniae FADDI-KP003 | Klebsiella pneumoniae FADDI-KP012 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Peptide | MIC (mg/L) | |||||||||||||||
| Colistin | 1 | 1 | 2 | >128 | >128 | 8 | 1 | 0.5 | 0.5 | 128 | 16 | 8 | 1 | >128 | 128 | 32 |
| PolymyxinB | 1 | 1 | 1 | 32 | >32 | 4 | 1 | 0.5 | 1 | 128 | 8 | 16 | 1 | 128 | >32 | 16 |
| FADDI-285 | 2 | 2 | 4 | >32 | 2 | 8 | 4 | 4 | 8 | >32 | 8 | 8 | 4 | >32 | >32 | >32 |
| FADDI-286 | 2 | 2 | 4 | >32 | 2 | 4 | 4 | 4 | 4 | >32 | 8 | 8 | 4 | >32 | >32 | >32 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, B.; Roberts, K.D.; Thompson, P.E.; Nation, R.L.; Velkov, T.; Li, J. Design and Evaluation of Novel Polymyxin Fluorescent Probes. Sensors 2017, 17, 2598. https://doi.org/10.3390/s17112598
Yun B, Roberts KD, Thompson PE, Nation RL, Velkov T, Li J. Design and Evaluation of Novel Polymyxin Fluorescent Probes. Sensors. 2017; 17(11):2598. https://doi.org/10.3390/s17112598
Chicago/Turabian StyleYun, Bo, Kade D. Roberts, Philip E. Thompson, Roger L. Nation, Tony Velkov, and Jian Li. 2017. "Design and Evaluation of Novel Polymyxin Fluorescent Probes" Sensors 17, no. 11: 2598. https://doi.org/10.3390/s17112598
APA StyleYun, B., Roberts, K. D., Thompson, P. E., Nation, R. L., Velkov, T., & Li, J. (2017). Design and Evaluation of Novel Polymyxin Fluorescent Probes. Sensors, 17(11), 2598. https://doi.org/10.3390/s17112598