
Detection of the Vascular Endothelial Growth Factor with a Novel Bioluminescence Resonance Energy Transfer Pair Using a Two-Component System

Abstract
:1. Introduction
2. Material and Methods
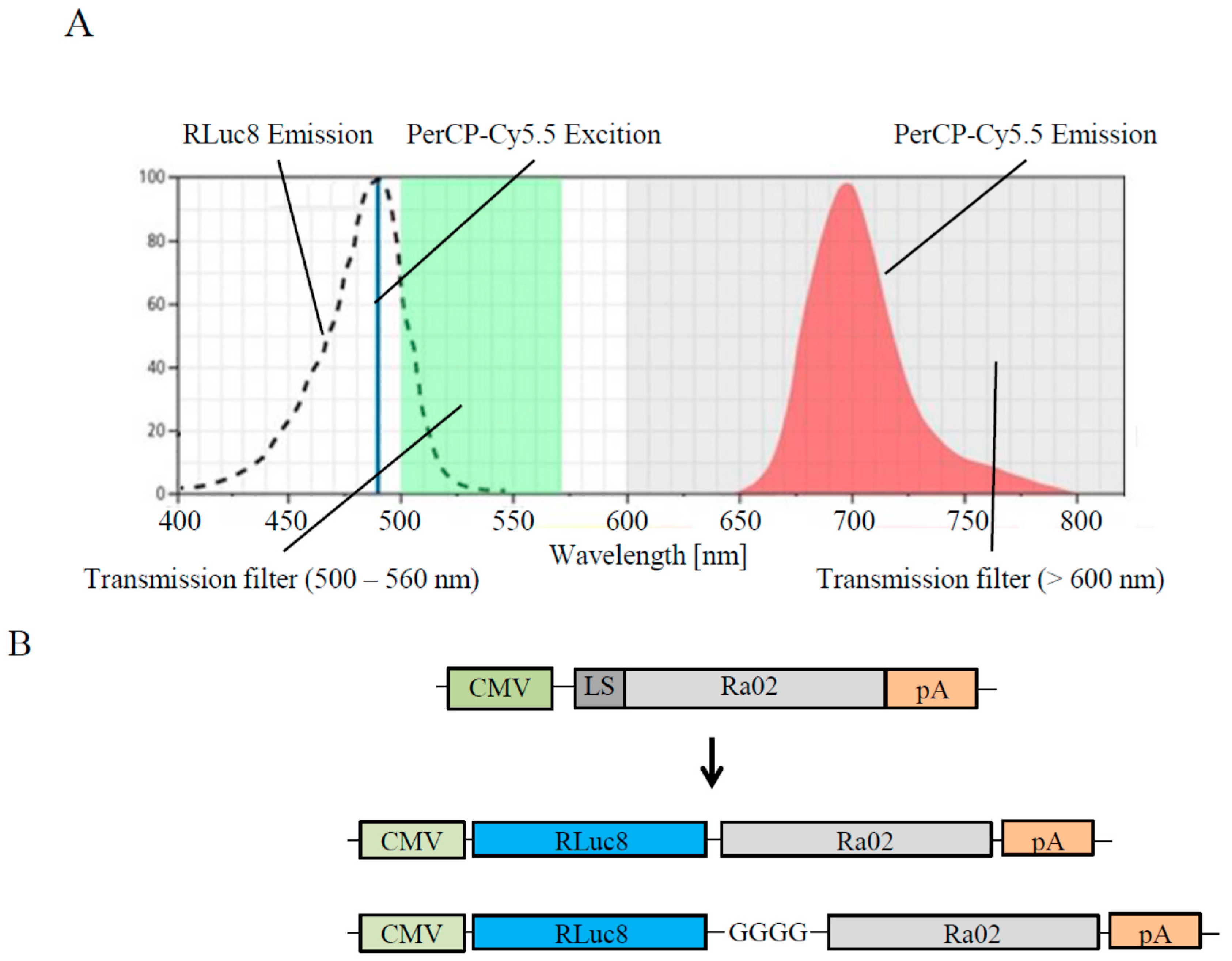
2.1. Preparation of BRET Constructs
2.2. Cell Culture and BRET Donor Expression
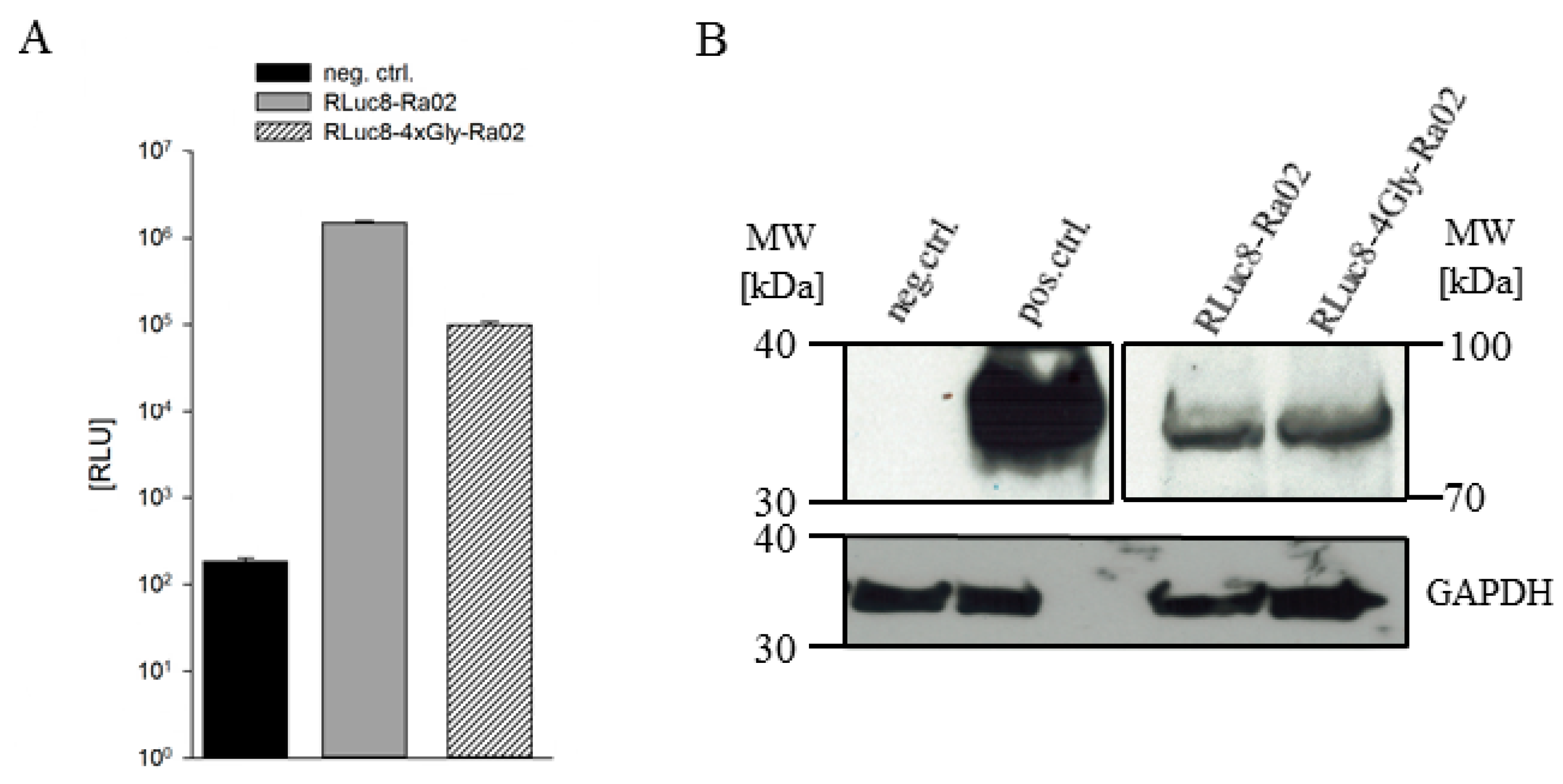
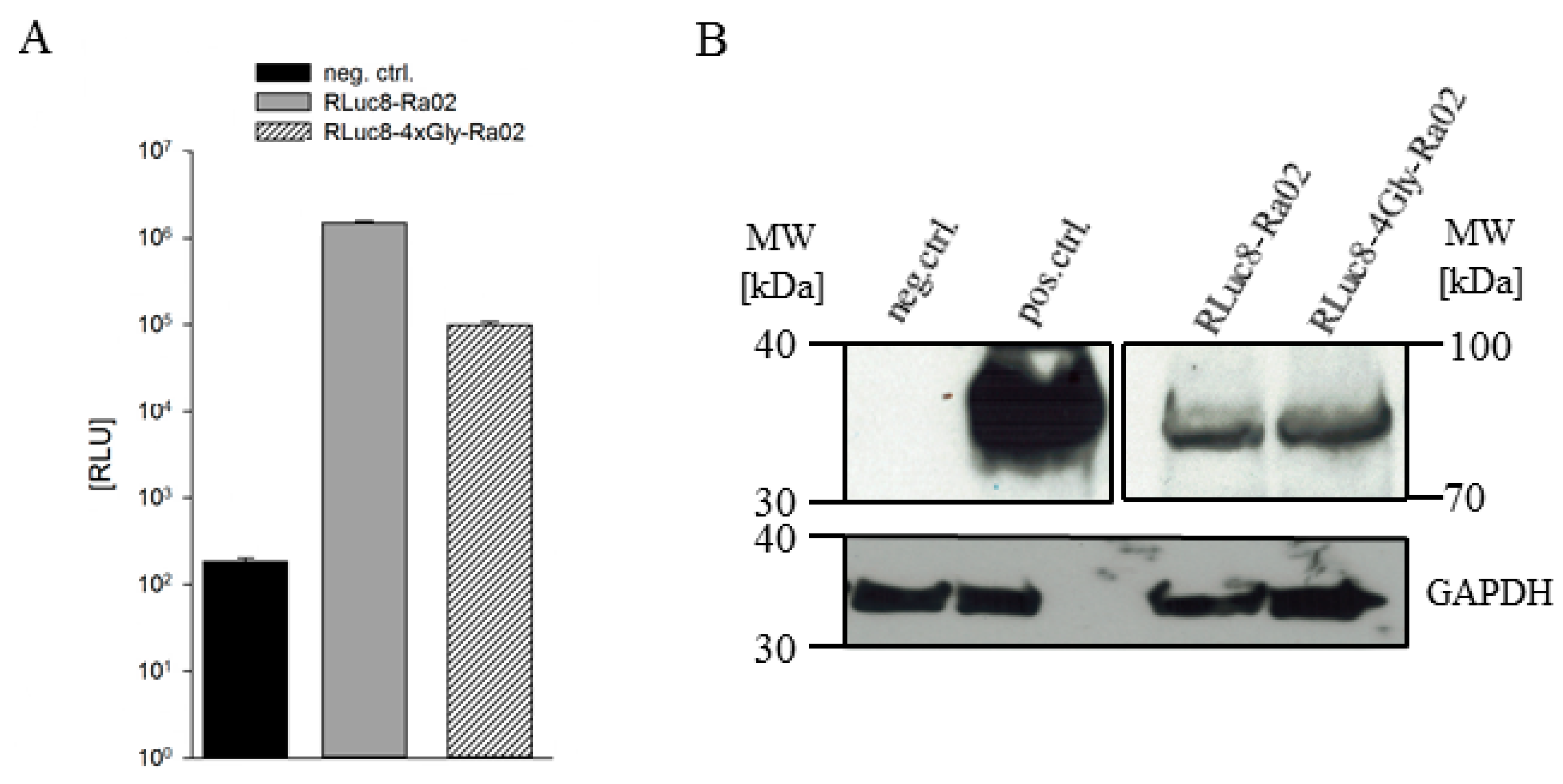
2.3. RLuc8 Activity
2.4. Western Blot Analysis
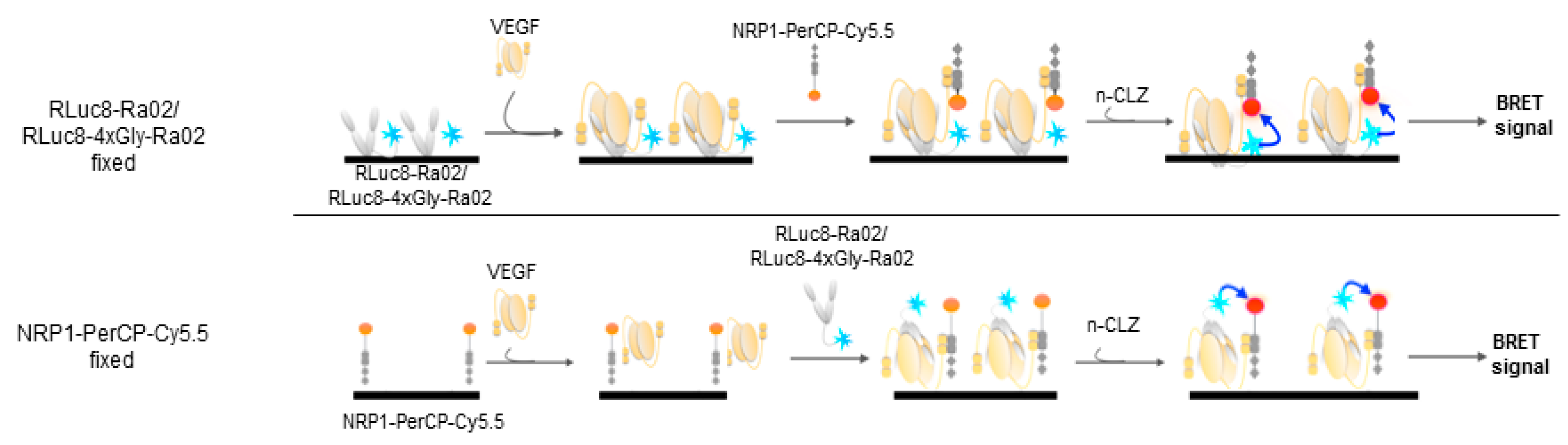
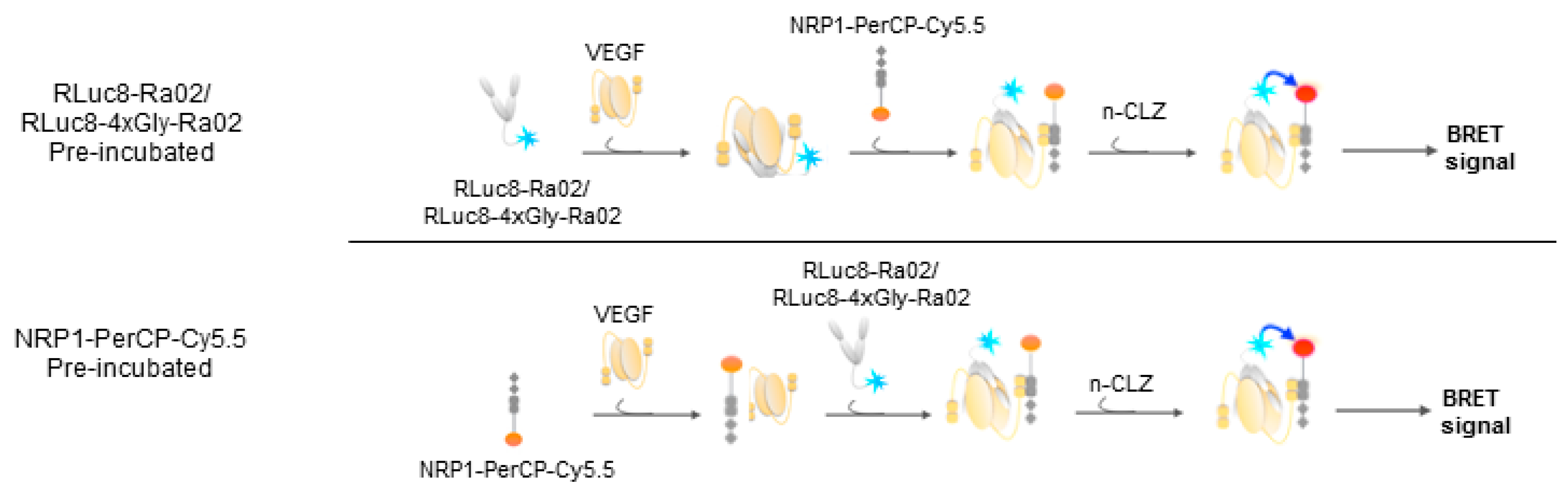
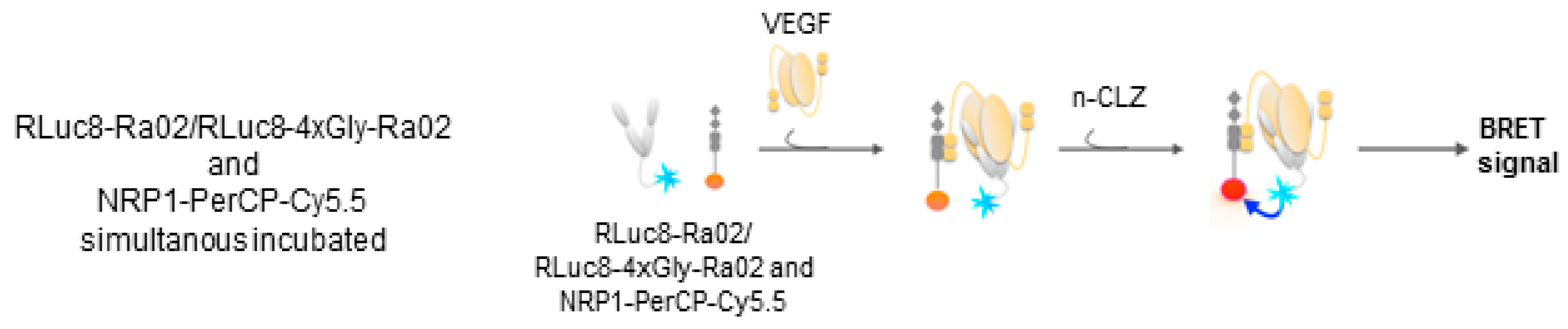
2.5. Experimental Setup VEGF Detection
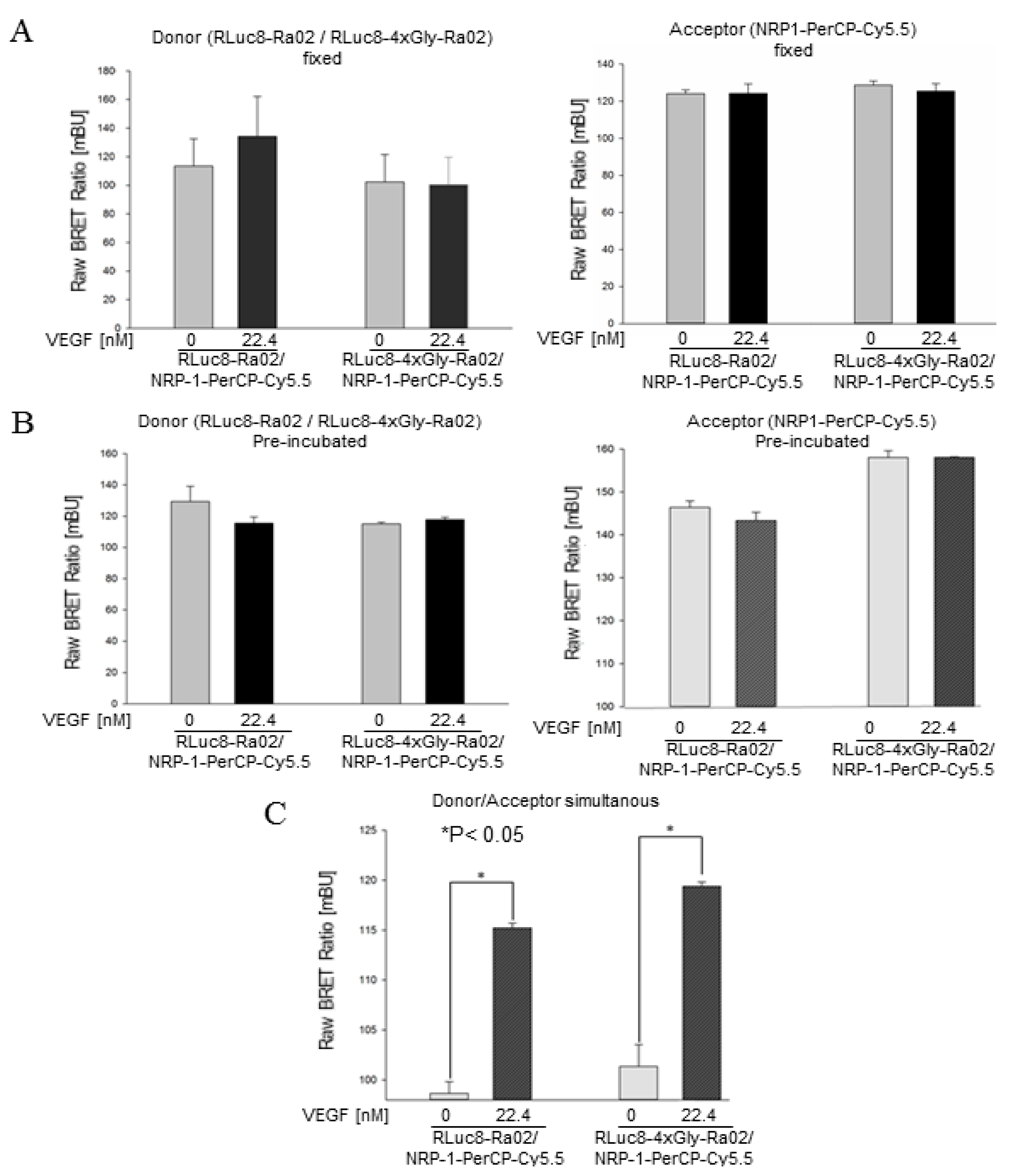
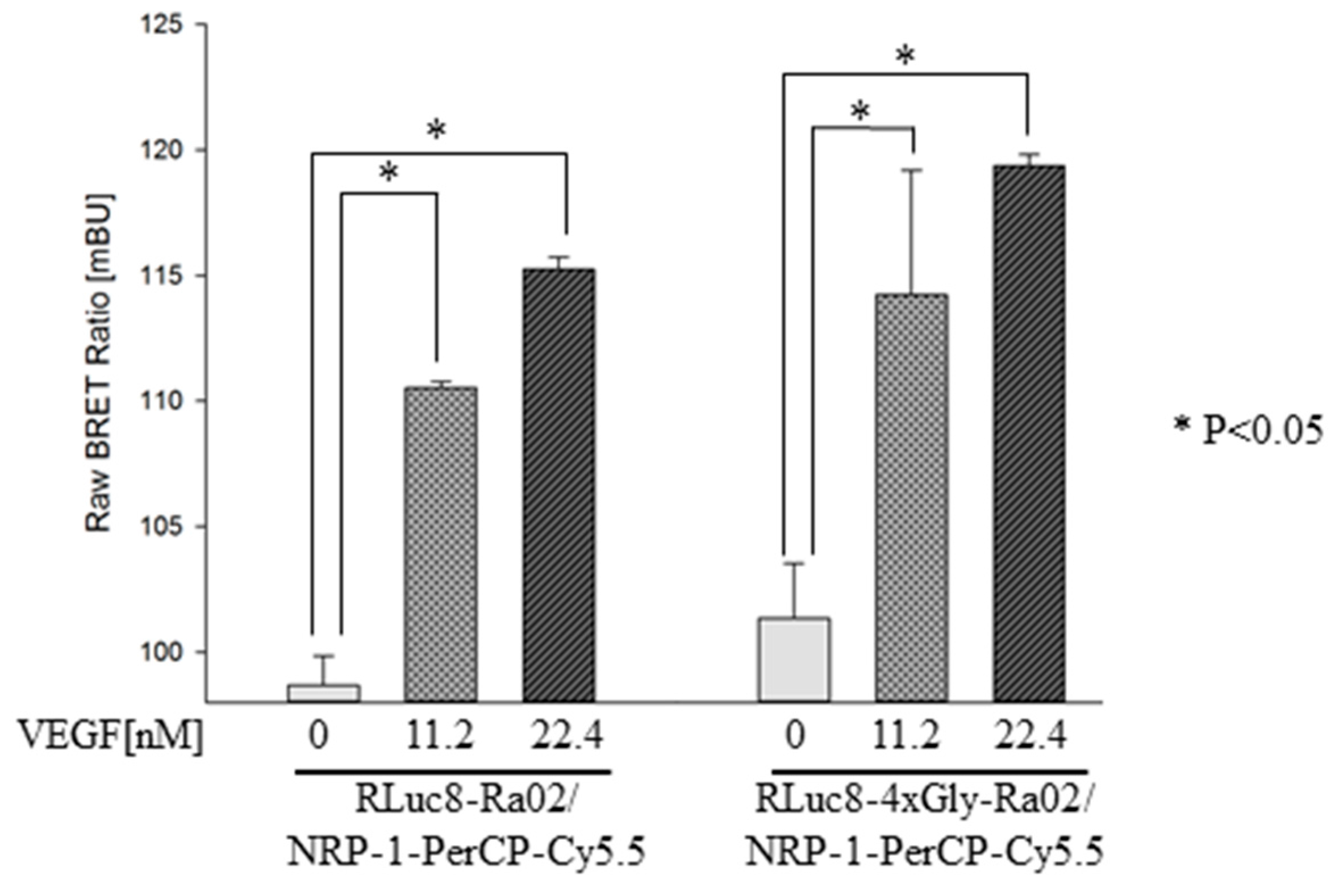
2.6. Measurement and Calculation of BRET Ratios
2.7. Statistics
3. Results and Discussion
3.1. Construct Generation, Expression and Functionality of Donor Molecules
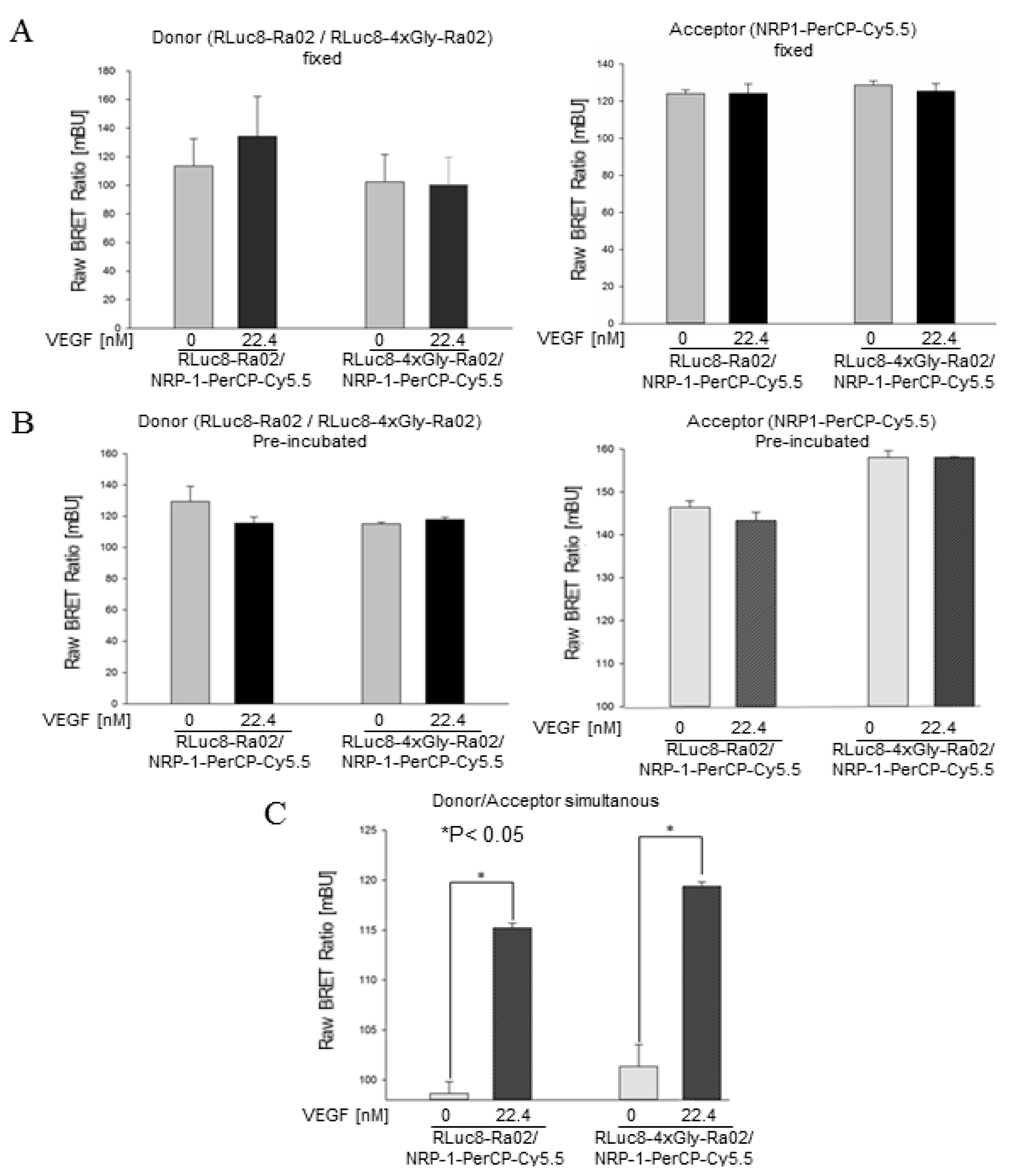
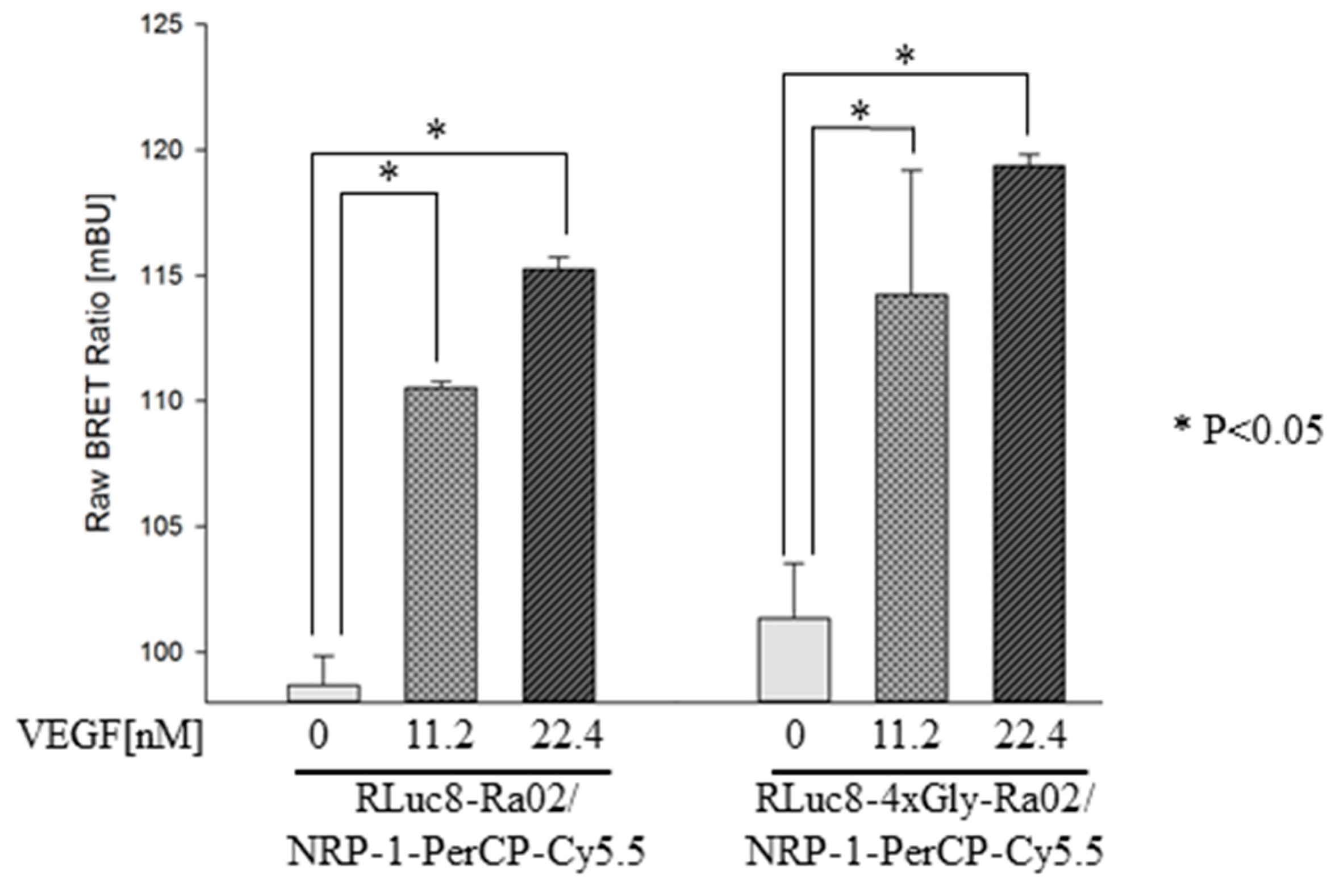
3.2. BRET Ratios of Different Experimental Setups
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Förster, T. Zwischenmolekulare Energiewanderung und Fluoreszenz. Ann. Phys. 1948, 437, 55–75. [Google Scholar] [CrossRef]
- Heyduk, T.; Heyduk, E. Molecular beacons for detecting DNA binding proteins. Nat. Biotechnol. 2002, 20, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Piston, D.W.; Johnson, C.H. A bioluminescence resonance energy transfer (BRET) system: Application to interacting circadian clock proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 151–156. [Google Scholar] [CrossRef] [PubMed]
- De, A.; Loening, A.M.; Gambhir, S.S. An improved bioluminescence resonance energy transfer strategy for imaging intracellular events in single cells and living subjects. Cancer Res. 2007, 67, 7175–7183. [Google Scholar] [CrossRef] [PubMed]
- Dragulescu-Andrasi, A.; Chan, C.T.; De, A.; Massoud, T.F.; Gambhir, S.S. Bioluminescence resonance energy transfer (BRET) imaging of protein-protein interactions within deep tissues of living subjects. Proc. Natl. Acad. Sci. USA 2011, 108, 12060–12065. [Google Scholar] [CrossRef] [PubMed]
- Dionne, P.; Mireille, C.; Labonte, A.; Carter-Allen, K.; Houle, B.; Joly, E.; Taylor, S.C.; Menard, L. BRET2: Efficient energy transfer from Renilla luciferase to GFP2 to measure protein-protein interactions and intracellular signalling events in live cells. In Luminescenc Biotechnology: Instruments and Applications; van Dyke, K., van Dyke, C., Woodfork, K., Eds.; CRC Press: Boca Ranton, FL, USA, 2002; pp. 539–555. [Google Scholar]
- Dacres, H.; Michie, M.; Wang, J.; Pfleger, K.D.; Trowell, S.C. Effect of enhanced Renilla luciferase and fluorescent protein variants on the Förster distance of Bioluminescence resonance energy transfer (BRET). Biochem. Biophys. Res. Commun. 2012, 425, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Machleidt, T.; Woodroofe, C.C.; Schwinn, M.K.; Méndez, J.; Robers, M.B.; Zimmerman, K.; Wood, K.V. NanoBRET A Novel BRET Platform for the Analysis of Protein-Protein Interactions. ACS Chem. Biol. 2015, 10, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Flajolet, M.; Agnati, L.F.; Greengard, P.; Fuxe, K. Bioluminescence Resonance Energy Transfer (BRET) Methods to study G Protein-coupled Receptor-Receptor Tyrosine Kinase Heteroreceptor Complexes. Methods Cell Biol. 2013, 117, 141–164. [Google Scholar] [PubMed]
- Wimmer, T.; Lorenz, B.; Stieger, K. Functional characterization of AAV-expressed recombinant anti-VEGF single-chain variable fragments in vitro. J. Ocul. Pharmacol. Ther. 2015, 31, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, T.; Lorenz, B.; Stieger, K. Quantification of the vascular endothelial growth factor with a bioluminescence resonance energy transfer (BRET) based single molecule biosensor. Biosens. Bioelectron. 2016, 86, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. Vascular endothelial growth factor: Basic science and clinical progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, C.; Fantin, A.; Lampropoulou, A.; Denti, L.; Chikh, A.; Ruhrberg, C. Imatinib inhibits VEGF-independent angiogenesis by targeting neuropilin 1-dependent ABL1 activation in endothelial cells. J. Exp. Med. 2014, 211, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Muether, P.S.; Hermann, M.M.; Viebahn, U.; Kirchhof, B.; Fauser, S. Vascular endothelial growth factor in patients with exudative age-related macular degeneration treated with ranibizumab. Ophthalmology 2012, 119, 2082–2086. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, K.; Chen, Z.; Shi, J.; Zhou, D.; Xie, G. A fluorescence biosensor for VEGF detection based on DNA assembly structure switching and isothermal amplification. Biosens. Bioelectron. 2017, 89 Pt 2, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Leung, K.H.; Lin, L.; Lin, L.; Lin, S.; Leung, C.H.; Lin, J.M. Determination of cell metabolite VEGF 165 and dynamic analysis of protein-DNA interactions by combination of microfluidic technique and luminescent switch-on probe. Biosens. Bioelectron. 2016, 79, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Muller, Y.A.; Chen, Y.; Christinger, H.W.; Li, B.; Cunningham, B.C.; Lowman, H.B.; de Vos, A.M. VEGF and the Fab fragment of a humanized neutralizing antibody: Crystal structure of the complex at 2.4 Å resolution and mutational analysis of the interface. Structure 1998, 6, 1153–1167. [Google Scholar] [CrossRef]
- Geretti, E.; Shimizu, A.; Kurschat, P.; Klagsbrun, M. Site-directed mutagenesis in the B-neuropilin-2 domain selectively enhances its affinity to VEGF165, but not to semaphorin 3F. J. Biol. Chem. 2007, 282, 25698–25707. [Google Scholar] [CrossRef] [PubMed]
- Fuh, G.; Wu, P.; Liang, W.C.; Ultsch, M.; Lee, C.V.; Moffat, B.; Wiesmann, C. Structure-function studies of two synthetic anti-vascular endothelial growth factor Fabs and comparison with the Avastin™ Fab. J. Biol. Chem. 2006, 281, 6625–6631. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, H.; Waheed, N. The evolving role of vascular endothelial growth factor inhibitors in the treatment of neovascular age-related macular degeneration. Eye 2008, 22, 761–767. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Energy Donor | Energy Acceptor | Luciferase Substrate | Spectral Resolution ∆λem (nm) | Ref. |
|---|---|---|---|---|---|
| BRET 1 | RLuc | eYFP | n-CLZ | 50 | [3] |
| BRET 2 | RLuc | GFP2 | CLZ400a | 115 | [6] |
| eBRET 2 | RLuc8 | GFP2 | CLZ400a | 115 | [4] |
| BRET 3 | RLuc8 | mOrange | n-CLZ | 85 | [5] |
| BRET 3.1 | RLuc8 | mOrange | CLZ-v | 50 | [5] |
| BRET 4.1 | RLuc8 | TagRFP | CLZ-v | 70 | [5] |
| BRET 5 | RLuc8.6 | TagRFP | n-CLZ | 50 | [5] |
| BRET 6 | RLuc8.6 | TurboFP | n-CLZ | 100 | [5] |
| BRET 6.1 | RLuc8.6 | TurboFP | CLZ-v | 65 | [5] |
| BRET 7 | RLuc8 | PerCP-Cy5.5 | n-CLZ | 300 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wimmer, T.; Schroeter, E.; Lorenz, B.; Stieger, K. Detection of the Vascular Endothelial Growth Factor with a Novel Bioluminescence Resonance Energy Transfer Pair Using a Two-Component System. Sensors 2017, 17, 145. https://doi.org/10.3390/s17010145
Wimmer T, Schroeter E, Lorenz B, Stieger K. Detection of the Vascular Endothelial Growth Factor with a Novel Bioluminescence Resonance Energy Transfer Pair Using a Two-Component System. Sensors. 2017; 17(1):145. https://doi.org/10.3390/s17010145
Chicago/Turabian StyleWimmer, Tobias, Eva Schroeter, Birgit Lorenz, and Knut Stieger. 2017. "Detection of the Vascular Endothelial Growth Factor with a Novel Bioluminescence Resonance Energy Transfer Pair Using a Two-Component System" Sensors 17, no. 1: 145. https://doi.org/10.3390/s17010145
APA StyleWimmer, T., Schroeter, E., Lorenz, B., & Stieger, K. (2017). Detection of the Vascular Endothelial Growth Factor with a Novel Bioluminescence Resonance Energy Transfer Pair Using a Two-Component System. Sensors, 17(1), 145. https://doi.org/10.3390/s17010145