Taxonomic and Metabolite Diversity of Actinomycetes Associated with Three Australian Ascidians



Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Ascidian Collection
2.2. Culture-Independent Microbial Diversity Analysis
2.3. Culture-Dependent Actinomycete Isolation and Identification
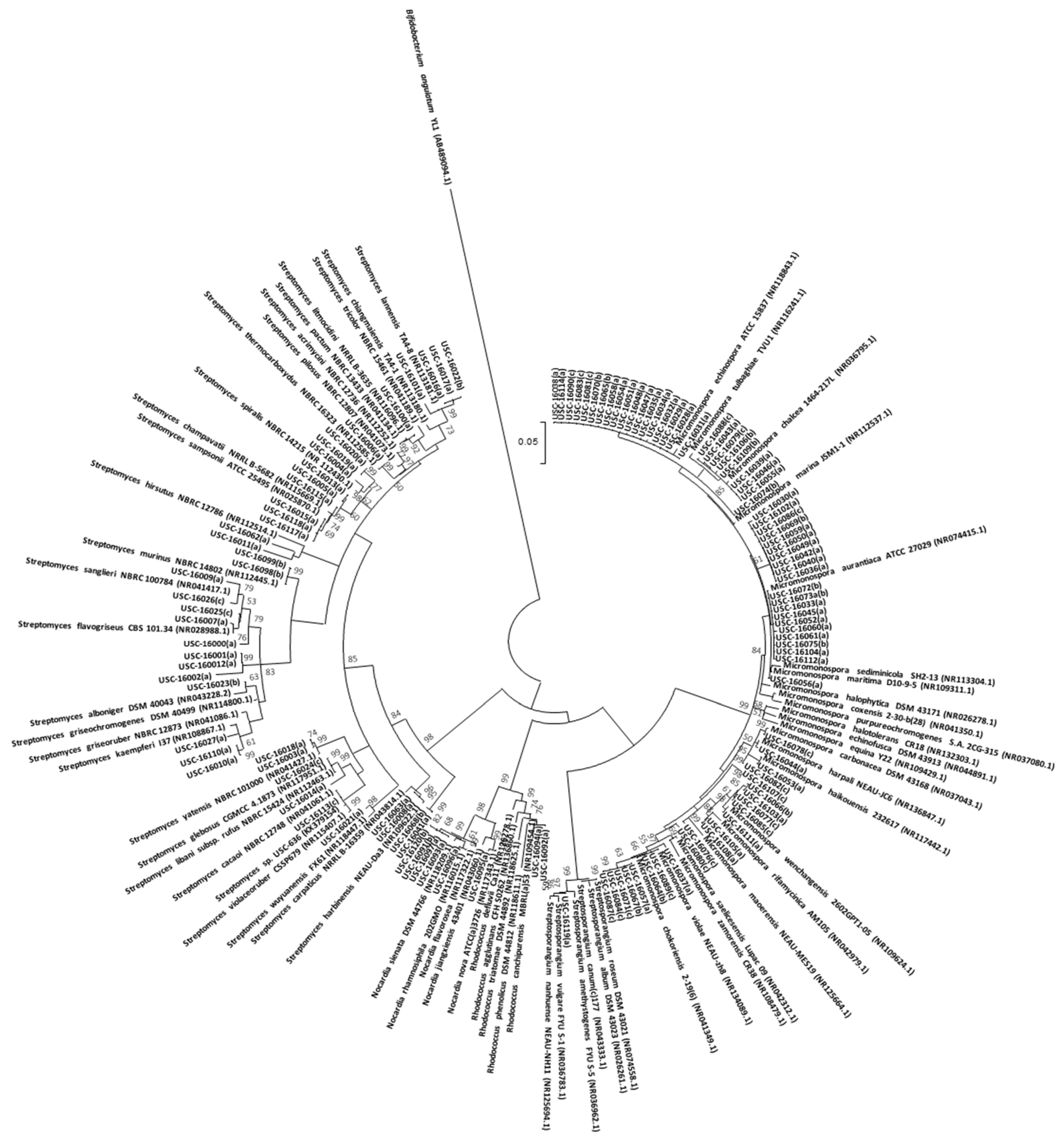
2.4. Molecular Identification and Phylogenetic Analysis of Actinomycete Isolates
2.5. Metabolite Diversity
3. Results
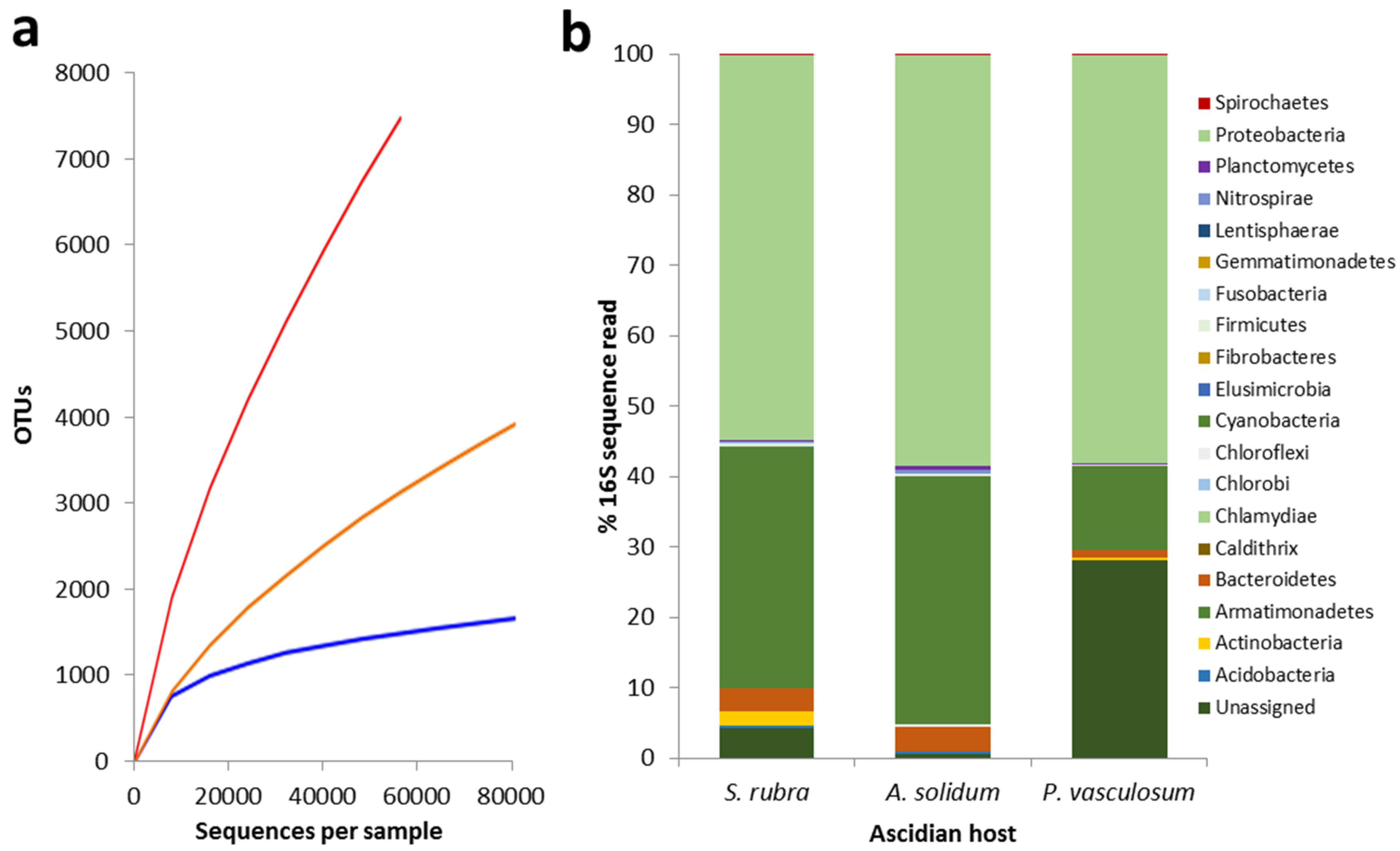
3.1. Microbial Diversity Associated with Three Australian Ascidians
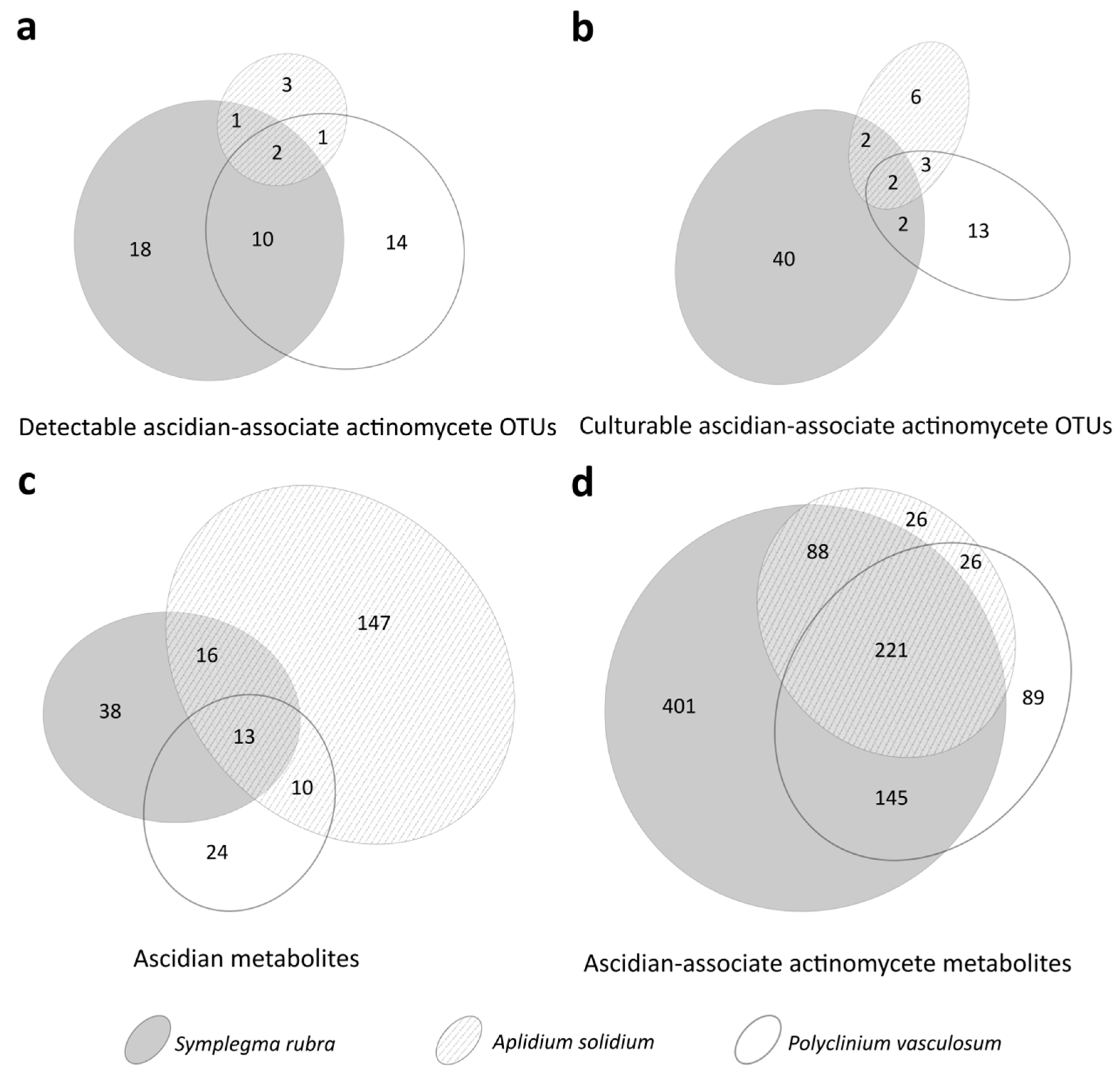
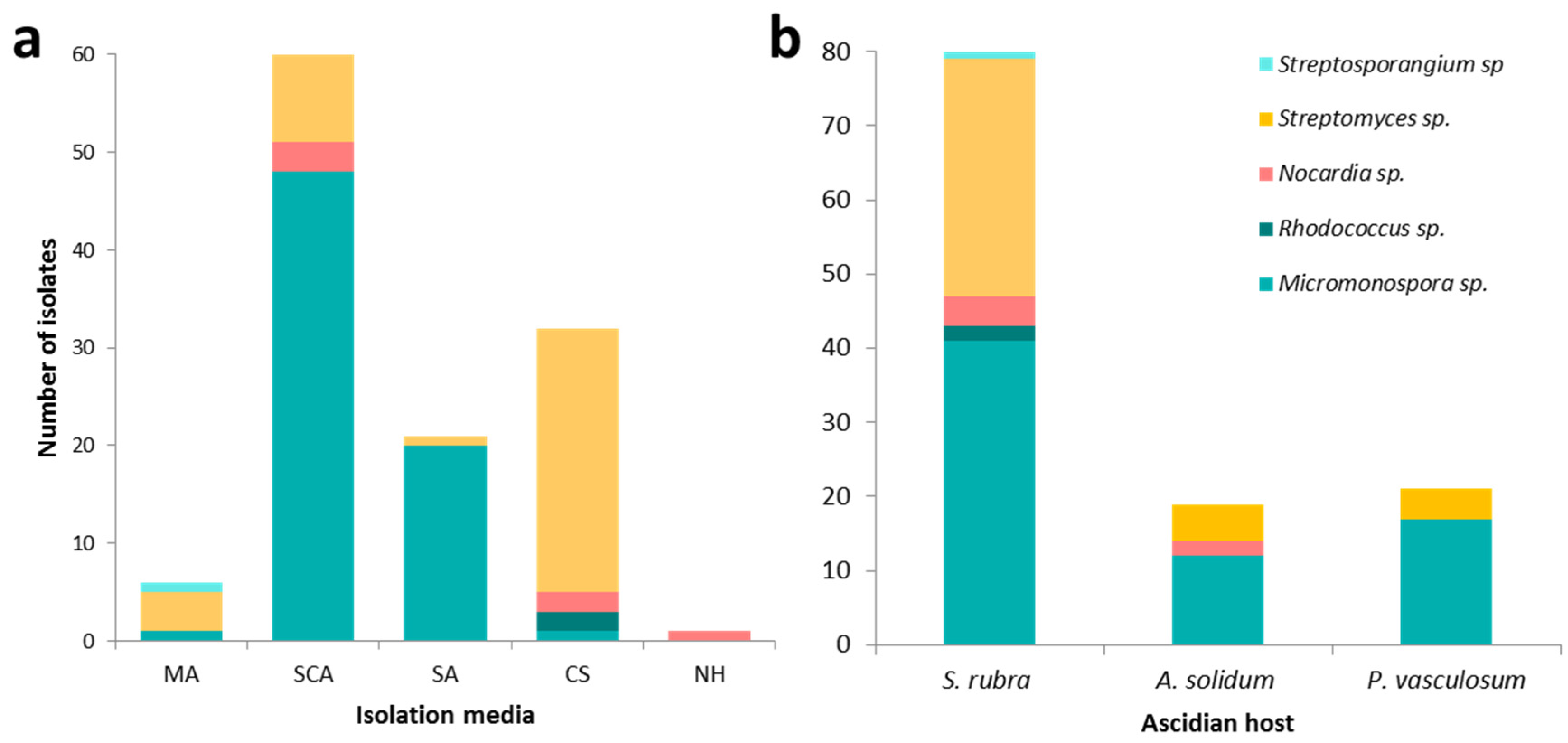
3.2. Culture-Dependent Actinomycete Diversity
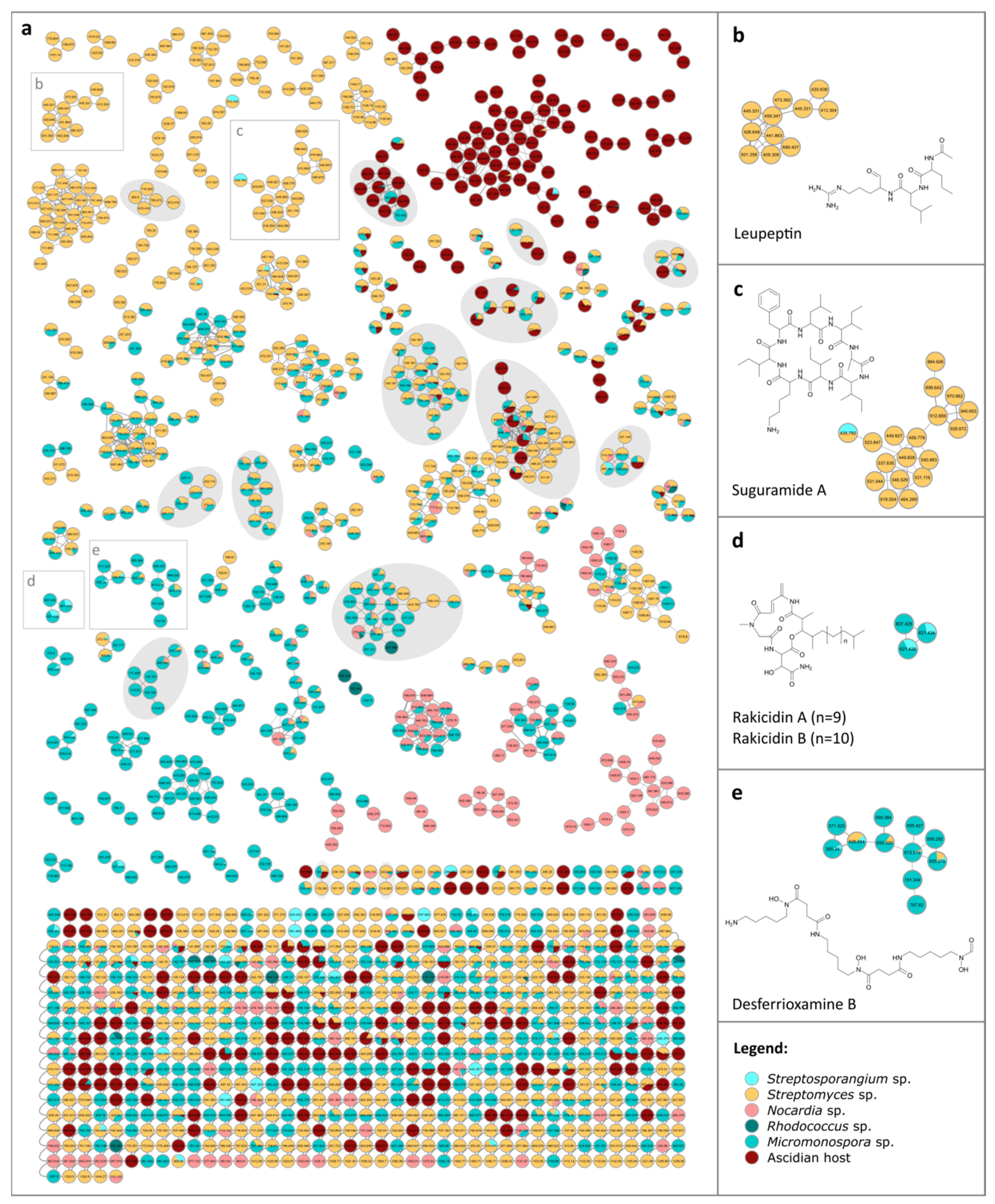
3.3. Metabolite Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paul, V.J.; Lindquist, N.; Fenical, W. Chemical defenses of the tropical ascidian Atapozoa sp. and its nudibranch predators Nembrotha spp. Mar. Ecol. Prog. Ser. 1990, 59, 109–118. [Google Scholar] [CrossRef]
- Lindquist, N.; Hay, M.E.; Fenical, W. Defense of ascidians and their conspicuous larvae: Adult vs. Larval chemical defenses. Ecol. Monogr. 1992, 62, 547–568. [Google Scholar] [CrossRef]
- Joullié, M.M.; Leonard, M.S.; Portonovo, P.; Liang, B.; Ding, X.; La Clair, J.J. Chemical defense in ascidians of the didemnidae family. J. Bioconj. Chem. 2003, 14, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.A. The Global Marine Pharmaceuticals Pipeline. Available online: http://marinepharmacology.midwestern.edu/clinPipeline.htm (accessed on 4 April 2017).
- Schmidt, E.W.; Nelson, J.T.; Rasko, D.A.; Sudek, S.; Eisen, J.A.; Haygood, M.G.; Ravel, J. Patellamide A and C biosynthesis by a microcin-like pathway in prochloron didemni, the cyanobacterial symbiont of lissoclinum patella. Proc. Natl. Acad. Sci. USA 2005, 102, 7315–7320. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.W. The secret to a successful relationship: Lasting chemistry between ascidians and their symbiotic bacteria. Invertebr. Biol. 2014, 134, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Sheridan, C.; Osinga, R.; Dionísio, G.; Rocha, R.J.M.; Silva, B.; Rosa, R.; Calado, R. Marine microorganism-invertebrate assemblages: Perspectives to solve the “supply problem” in the initial steps of drug discovery. Mar. Drugs 2014, 12, 3929–3952. [Google Scholar] [CrossRef] [PubMed]
- Radjasa, O.K.; Vaske, Y.M.; Navarro, G.; Vervoort, H.C.; Tenney, K.; Linington, R.G.; Crews, P. Highlights of marine invertebrate-derived biosynthetic products: Their biomedical potential and possible production by microbial associants. Bioorg. Med. Chem. 2011, 19, 6658–6674. [Google Scholar] [CrossRef] [PubMed]
- Tianero, M.D.B.; Kwan, J.C.; Wyche, T.P.; Presson, A.P.; Koch, M.; Barrows, L.R.; Bugni, T.S.; Schmidt, E.W. Species specificity of symbiosis and secondary metabolism in ascidians. ISME J. 2014, 9, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Erwin, P.M.; Pineda, M.C.; Webster, N.; Turon, X.; Lopez-Legentil, S. Down under the tunic: Bacterial biodiversity hotspots and widespread ammonia-oxidizing archaea in coral reef ascidians. ISME J. 2014, 8, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Fenical, W. Strategies for the discovery of secondary metabolites from marine bacteria: Ecological perspectives. Annu. Rev. Microbiol. 1994, 48, 559–584. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Schmidt, E.W. Bacterial endosymbiosis in a chordate host: Long-term co-evolution and conservation of secondary metabolism. PLoS ONE 2013, 8, e80822. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Tianero, M.D.B.; Donia, M.S.; Wyche, T.P.; Bugni, T.S.; Schmidt, E.W. Host control of symbiont natural product chemistry in cryptic populations of the tunicate lissoclinum patella. PLoS ONE 2014, 9, e95850. [Google Scholar] [CrossRef] [PubMed]
- Kremer, B.; Pardy, R.; Lewin, R. Carbon fixation and photosynthates of prochloron, a green alga symbiotic with an ascidian, lissoclinum patella. Phycologia 1982, 21, 258–263. [Google Scholar] [CrossRef]
- Koike, I.; Suzuki, T. Nutritional diversity of symbiotic ascidians in a fijian seagrass meadow. Ecol. Res. 1996, 11, 381–386. [Google Scholar] [CrossRef]
- Münchhoff, J.; Hirose, E.; Maruyama, T.; Sunairi, M.; Burns, B.P.; Neilan, B.A. Host specificity and phylogeography of the prochlorophyte Prochloron sp., an obligate symbiont in didemnid ascidians. Environ. Microbiol. 2007, 9, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Fricke, W.F.; Partensky, F.; Cox, J.; Elshahawi, S.I.; White, J.R.; Phillippy, A.M.; Schatz, M.C.; Piel, J.; Haygood, M.G. Complex microbiome underlying secondary and primary metabolism in the tunicate-prochloron symbiosis. Proc. Natl. Acad. Sci. USA 2011, 108, E1423–E1432. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Fricke, W.F.; Ravel, J.; Schmidt, E.W. Variation in tropical reef symbiont metagenomes defined by secondary metabolism. PLoS ONE 2011, 6, 17897. [Google Scholar] [CrossRef] [PubMed]
- Steinert, G.; Taylor, M.W.; Schupp, P.J. Diversity of actinobacteria associated with the marine ascidian eudistoma toealensis. Mar. Biotechnol. 2015, 17, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Fu, C.; Wang, G. Microbial diversity associated with ascidians: A review of research methods and application. Symbiosis 2016, 71, 19–26. [Google Scholar] [CrossRef]
- Jimenez, P.C.; Ferreira, E.G.; Araújo, L.A.; Guimarães, L.A.; Sousa, T.S.; Pessoa, O.D.L.; Lotufo, T.M.; Costa-Lotufo, L.V. Cytotoxicity of actinomycetes associated with the ascidian eudistoma vannamei (millar, 1977), endemic of northeastern coast of brazil/citotoxicidad de actinomicetos asociada a la ascidia eudistoma vannamei (millar, 1977), endémica de la costa noreste de Brasil. Lat. Am. J. Aquat. Res. 2013, 41, 335. [Google Scholar]
- List of Prokaryotic Names with Standing in Nomenclature. Available online: http://www.bacterio.net/actinomycetales.html (accessed on 16 November 2017).
- Kurtböke, D.I. Ecology and habitat disctribution of actinobacteria. In Biology and Biotechnology of Actinobacteria; Wink, J., Hamedi, J., Eds.; Springer: Berlin, Germany, 2017; pp. 123–149. [Google Scholar]
- He, H.; Ding, W.-D.; Bernan, V.S.; Richardson, A.D.; Ireland, C.M.; Greenstein, M.; Ellestad, G.A.; Carter, G.T. Lomaiviticins a and b, potent antitumor antibiotics from micromonospora lomaivitiensis. J. Am. Chem. Soc. 2001, 123, 5362–5363. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Piel, J. A gene cluster from a marine streptomyces encoding the biosynthesis of the aromatic spiroketal polyketide griseorhodin A. Chem. Biol. 2002, 9, 1017–1026. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Shirasaki, S.; Kawasaki, T.; Matsuo, Y.; Adachi, K.; Shizuri, Y. Structures of new cytotoxic antibiotics, piericidins C7 and C8. J. Antibiot. 2007, 60, 201. [Google Scholar] [CrossRef] [PubMed]
- Wyche, T.P.; Hou, Y.; Vazquez-Rivera, E.; Braun, D.; Bugni, T.S. Peptidolipins bf, antibacterial lipopeptides from an ascidian-derived Nocardia sp. J. Nat. Prod. 2012, 75, 735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Adnani, N.; Braun, D.R.; Ellis, G.A.; Barns, K.J.; Parker-Nance, S.; Guzei, I.A.; Bugni, T.S. Micromonohalimanes A and B: Antibacterial halimane-type diterpenoids from a marine micromonospora species. J. Nat. Prod. 2016, 79, 2968–2972. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Longley, R.; McConnell, O.; Ballas, L. Staurosporine aglycone (K252-c) and arcyriaflavin a from the marine ascidian, Eudistoma sp. Cell. Mol. Life Sci. 1994, 50, 843–845. [Google Scholar] [CrossRef]
- Andréo, M.A.; Jimenez, P.C.; Siebra, J.B.; Costa-Lotufo, L.V.; Vessecchi, R.; Niehues, M.; Lopes, J.L.; Lopes, N.P. Systematic UPLC-ESI-MS/MS study on the occurrence of staurosporine and derivatives in associated marine microorganisms from eudistoma vannamei. J. Braz. Chem. Soc. 2012, 23, 335–343. [Google Scholar] [CrossRef]
- Leal, M.C.; Puga, J.; Serodio, J.; Gomes, N.C.M.; Calado, R. Trends in the Discovery of New Marine Natural Products from Invertebrates over the Last Two Decades—Where and What Are We Bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef] [PubMed]
- Sherman, C.D.; Hunt, A.; Ayre, D.J. Is life history a barrier to dispersal? Contrasting patterns of genetic differentiation along an oceanographically complex coast. Biol. J. Linn. Soc. 2008, 95, 106–116. [Google Scholar] [CrossRef]
- Hastings Point. Microsoft 2017. Available online: www.bing.com/maps (accessed on 10 August 2017).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. Pandaseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Küster, E.; Williams, S. Selection of media for isolation of streptomycetes. Nature 1964, 202, 928–929. [Google Scholar] [CrossRef]
- English, A.L.; Boufridi, A.; Quinn, R.; Kurtböke, D. Evaluation of fermentation conditions triggering increased antibacterial activity from a near-shore marine intertidal environment-associated streptomyces species. Synth. Syst. Biotechnol. 2017, 2, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Microbiological Media; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Williams, S.T.; Wellington, E.M.H. Actinomycetes. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 969–987. [Google Scholar]
- Wellington, E. Preservation of actinomycete inoculum in frozen glycerol. Microbios. Lett. 1979, 6, 151–157. [Google Scholar]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T. Sharing and community curation of mass spectrometry data with global natural products social molecular networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Ninomiya, A.; Naruse, M.; Sun, Y.; Miyazaki, M.; Nogi, Y.; Okada, S.; Matsunaga, S. Surugamides a–e, cyclic octapeptides with four d-amino acid residues, from a marine Streptomyces sp.: Lc–ms-aided inspection of partial hydrolysates for the distinction of d-and l-amino acid residues in the sequence. J. Org. Chem. 2013, 78, 6746–6750. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, T.; Takeuchi, T.; Matsuzaki, A.; Kawamura, K.; Kondo, S.; Hamada, M.; Maeda, K.; Umezawa, H. Leupeptins, new protease inhibitors from actinomycetes. J. Antibiot. 1969, 22, 283–286. [Google Scholar] [CrossRef] [PubMed]
- McBrien, K.D.; Berry, R.L.; Lowe, S.E.; Neddermann, K.M.; Bursuker, I.; Huang, S.; Klohr, S.E.; Leet, J.E. Rakicidins, new cytotoxic lipopeptides from Micromonospora sp. Fermentation, isolation and characterization. J. Antibiot. 1995, 48, 1446–1452. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, M.; Díaz-Valdés, M.; Wanner, G.; Ramos-Esplá, A.; Antón, J. Microbial community associated with the colonial ascidian cystodytes dellechiajei. Environ. Microbiol. 2007, 9, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Tait, E.; Carman, M.; Sievert, S.M. Phylogenetic diversity of bacteria associated with ascidians in EEL pond (Woods Hole, Massachusetts, USA). J. Exp. Mar. Biol. Ecol. 2007, 342, 138–146. [Google Scholar] [CrossRef]
- Behrendt, L.; Larkum, A.W.; Trampe, E.; Norman, A.; Sørensen, S.J.; Kühl, M. Microbial diversity of biofilm communities in microniches associated with the didemnid ascidian lissoclinum patella. ISME J. 2012, 6, 1222–1237. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.L.; Darling, A.E.; Eisen, J.A. Metagenomic sequencing of an in vitro-simulated microbial community. PLoS ONE 2010, 5, e10209. [Google Scholar] [CrossRef] [PubMed]
- Sponga, F.; Cavaletti, L.; Lazzarini, A.; Borghi, A.; Ciciliato, I.; Losi, D.; Marinelli, F. Biodiversity and potentials of marine-derived microorganisms. J. Biotechnol. 1999, 70, 65–69. [Google Scholar] [CrossRef]
- Jensen, P.R.; Gontang, E.; Mafnas, C.; Mincer, T.J.; Fenical, W. Culturable marine actinomycete diversity from tropical pacific ocean sediments. Environ. Microbiol. 2005, 7, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Penn, K.; Jensen, P.R. Comparative genomics reveals evidence of marine adaptation in salinispora species. BMC Genom. 2012, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Kurtböke, D.İ.; Grkovic, T.; Quinn, R.J. Marine actinomycetes in biodiscovery. In Springer Handbook of Marine Biotechnology; Kim, S.-K., Ed.; Springer: Berlin, Germany, 2015; pp. 663–676. [Google Scholar]
- Sunagawa, S.; Woodley, C.M.; Medina, M. Threatened corals provide underexplored microbial habitats. PLoS ONE 2010, 5, e9554. [Google Scholar] [CrossRef] [PubMed]
- Okami, Y.; Okazaki, T.; Kitahara, T.; Umezawa, H. Studies on marine microorganisms. V. J. Antibiot. 1976, 29, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Imada, C.; Koseki, N.; Kamata, M.; Kobayashi, T.; Hamada-Sato, N. Isolation and characterization of antibacterial substances produced by marine actinomycetes in the presence of seawater. Actinomycetologica 2007, 21, 27–31. [Google Scholar] [CrossRef]
- Kott, P. Ascidiacea (Tunicata) from deep waters of the continental shelf of Western Australia. J. Nat. Hist. 2008, 42, 1103–1217. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Triggering cryptic natural product biosynthesis in microorganisms. Org. Biomol. Chem. 2009, 7, 1753–1760. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No of OTUs | Family | Ascidian Host |
|---|---|---|
| 1 | Acidothermaceae | Polyclinum vasculosum |
| 1 | Actinomycetaceae | Aplidium solidum |
| 1 | Cellulomonadaceae | Polyclinum vasculosum |
| 2 | Corynebacteriaceae | Symplegma rubra, Polyclinum vasculosum, Aplidium solidum |
| 1 | Gordoniaceae | Symplegma rubra |
| 2 | Intrasporangiaceae | Polyclinum vasculosum |
| 1 | Kineosporiaceae | Symplegma rubra, Polyclinum vasculosum |
| 5 | Microbacteriaceae | Symplegma rubra, Polyclinum vasculosum, Aplidium solidum |
| 1 | Micrococcaceae | Aplidium solidum |
| 3 | Micromonosporaceae | Symplegma rubra, Polyclinum vasculosum |
| 6 | Mycobacteriaceae | Symplegma rubra, Polyclinum vasculosum |
| 5 | Nocaridaceae | Symplegma rubra, Polyclinum vasculosum, Aplidium solidum |
| 1 | Promicromonosporaceae | Polyclinum vasculosum |
| 1 | Pseudonocardiaceae | Symplegma rubra Aplidium solidum |
| 4 | Streptomycetaceae | Symplegma rubra, Polyclinum vasculosum, Aplidium solidum |
| 3 | Streptosporangiaceae | Symplegma rubra, Polyclinum vasculosum |
| 6 | Thermomonosporaceae | Symplegma rubra, Polyclinum vasculosum |
| 4 | Ambiguous assignment | Symplegma rubra, Polyclinum vasculosum |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buedenbender, L.; Carroll, A.R.; Ekins, M.; Kurtböke, D.İ. Taxonomic and Metabolite Diversity of Actinomycetes Associated with Three Australian Ascidians. Diversity 2017, 9, 53. https://doi.org/10.3390/d9040053
Buedenbender L, Carroll AR, Ekins M, Kurtböke Dİ. Taxonomic and Metabolite Diversity of Actinomycetes Associated with Three Australian Ascidians. Diversity. 2017; 9(4):53. https://doi.org/10.3390/d9040053
Chicago/Turabian StyleBuedenbender, Larissa, Anthony R. Carroll, Merrick Ekins, and D. İpek Kurtböke. 2017. "Taxonomic and Metabolite Diversity of Actinomycetes Associated with Three Australian Ascidians" Diversity 9, no. 4: 53. https://doi.org/10.3390/d9040053
APA StyleBuedenbender, L., Carroll, A. R., Ekins, M., & Kurtböke, D. İ. (2017). Taxonomic and Metabolite Diversity of Actinomycetes Associated with Three Australian Ascidians. Diversity, 9(4), 53. https://doi.org/10.3390/d9040053