Duplex Alu Screening for Degraded DNA of Skeletal Human Remains

Abstract
1. Introduction
2. Materials and Methods
2.1. Material
2.2. DNA Extraction from Skeletal Material with the QIAvac-24-Plus
2.3. Modern DNA
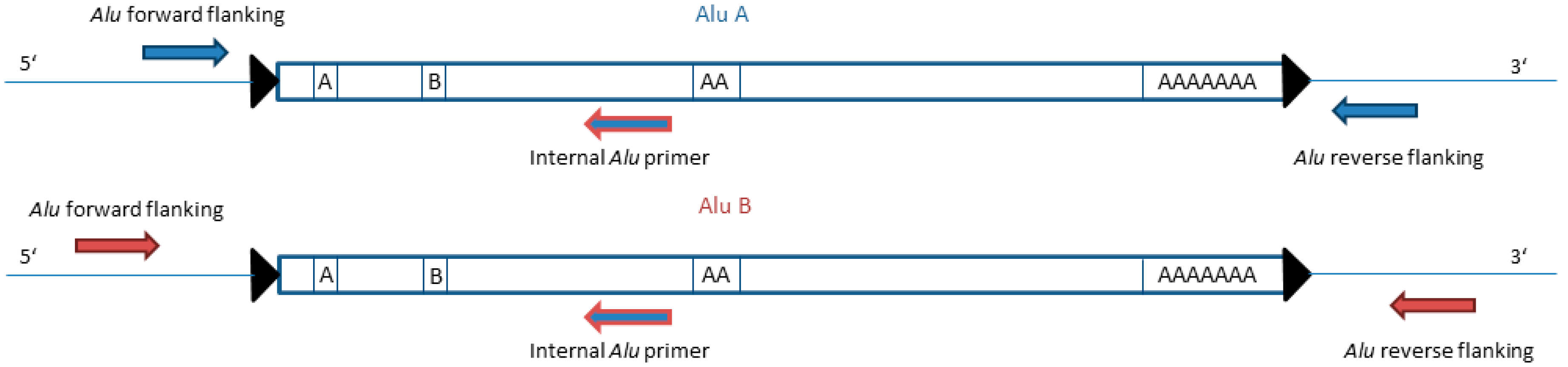
2.4. Primer Design for Alu Duplex PCRs
2.5. PCR
2.6. Gel Electrophoresis and Fragment-Length Determination
2.7. PCR Purification and Sequencing
2.8. Laboratory Conditions
3. Results
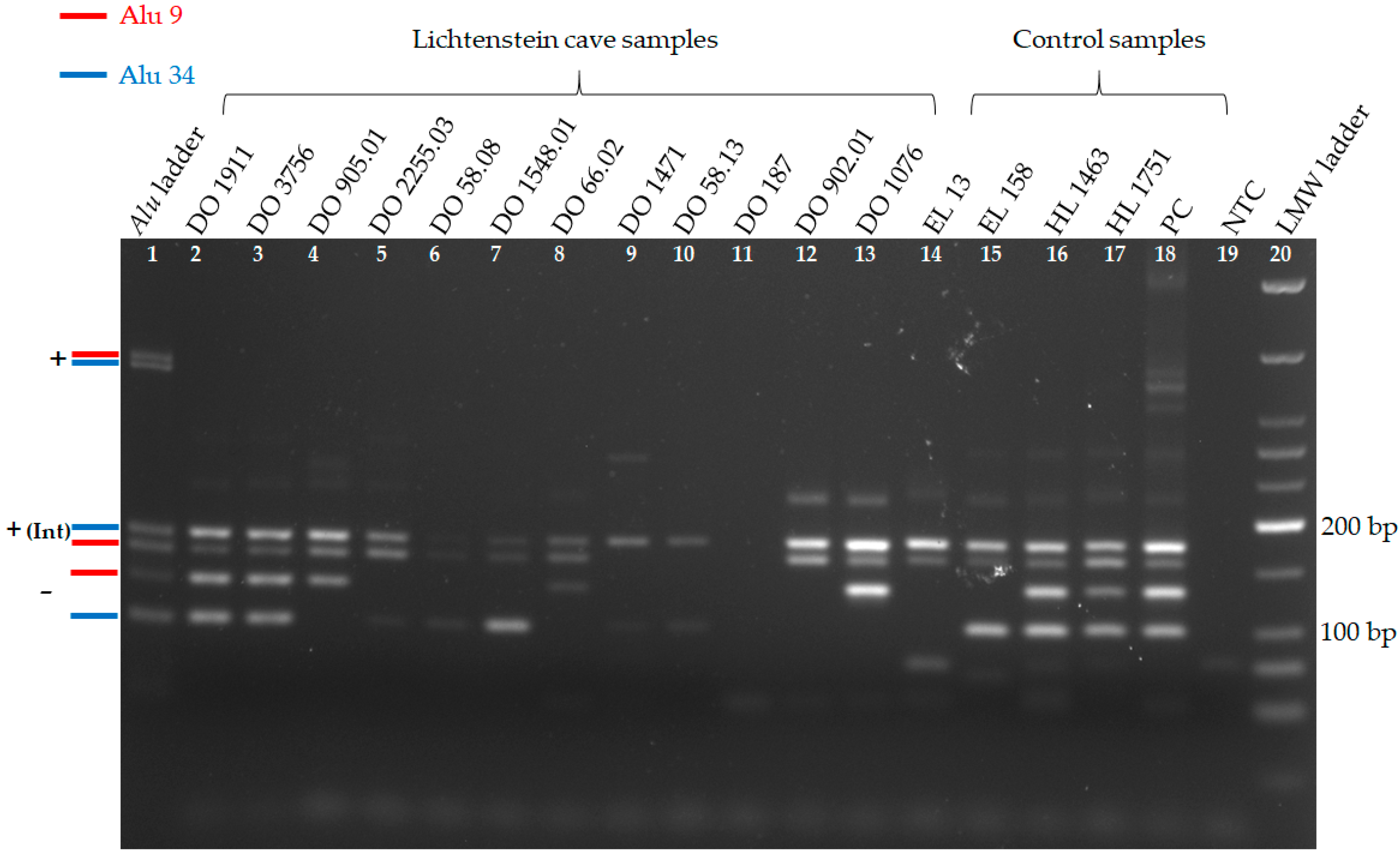
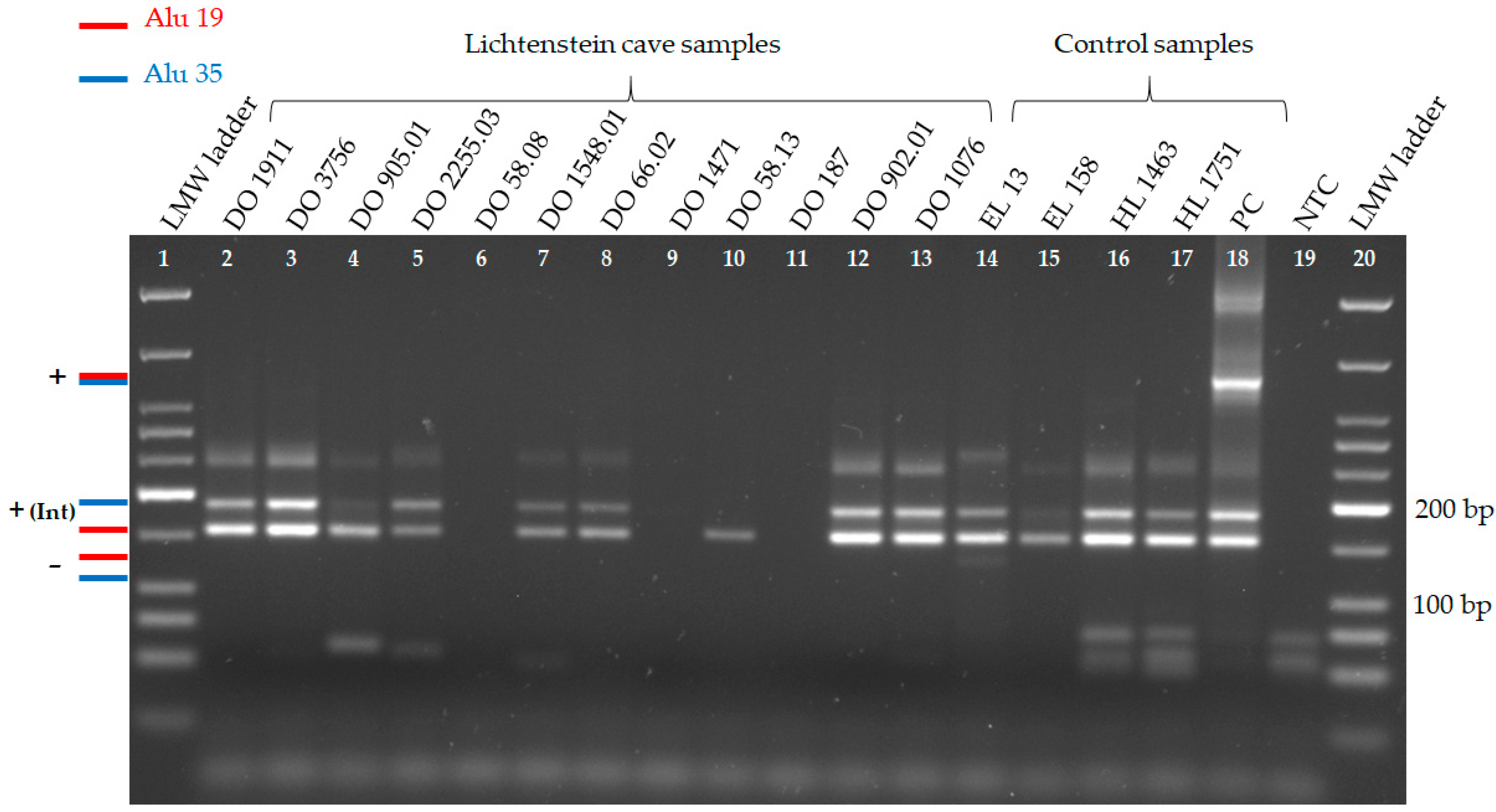
3.1. Alu Duplex PCRs
3.2. Proof of Authenticity
4. Discussion
4.1. Alu Duplex PCRs
4.2. Authenticity of Samples
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miller, W.J.; Capy, P. Mobile Genetic Elements as Natural Tools for Genome Evolution. In Mobile Genetic Elements: Protocols and Genomic Applications; Miller, W.J., Capy, P., Eds.; Humana Press: Totowa, NJ, USA, 2004; Volume 260, pp. 1–20. ISBN 978-1-58829-007-6. [Google Scholar]
- Volff, J.-N. Turning junk into gold: Domestication of transposable elements and the creation of new genes in eukaryotes. BioEssays News Rev. Mol. Cell. Dev. Biol. 2006, 28, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Piskurek, O.; Jackson, D.J. Transposable elements: From DNA parasites to architects of metazoan evolution. Genes 2012, 3, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Estecio, M.R.H.; Gallegos, J.; Dekmezian, M.; Lu, Y.; Liang, S.; Issa, J.-P.J. SINE retrotransposons cause epigenetic reprogramming of adjacent gene promoters. Mol. Cancer Res. 2012, 10, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Han, J.S.; Boeke, J.D. LINE-1 retrotransposons: Modulators of quantity and quality of mammalian gene expression? BioEssays News Rev. Mol. Cell. Dev. Biol. 2005, 27, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Peaston, A.E.; Evsikov, A.V.; Graber, J.H.; de Vries, W.N.; Holbrook, A.E.; Solter, D.; Knowles, B.B. Retrotransposons regulate host genes in mouse oocytes and preimplantation embryos. Dev. Cell 2004, 7, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [PubMed]
- De Koning, A.P.J.; Gu, W.; Castoe, T.A.; Batzer, M.A.; Pollock, D.D. Repetitive Elements May Comprise Over Two-Thirds of the Human Genome. PLoS Genet. 2011, 7, e1002384. [Google Scholar] [CrossRef] [PubMed]
- López-Flores, I.; Garrido-Ramos, M.A. The Repetitive DNA Content of Eukaryotic Genomes; Karger Publishers: Basel, Switzerland, 2012; Volume 7, ISBN 978-3-318-02150-9. [Google Scholar]
- Feschotte, C.; Pritham, E.J. DNA Transposons and the Evolution of Eukaryotic Genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Ostertag, E.M.; Kazazian, H.H., Jr. Biology of mammalian L1 retrotransposons. Annu. Rev. Genet. 2001, 35, 501–538. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Hamada, M.; Teral, Y.; Okada, N. The 3′ Ends of tRNA-Derived Short Interspersed Repetitive Elements Are Derived from the 3′ Ends of Long Interspersed Repetitive Elements. Mol. Cell. Biol. 1996, 16, 3756–3764. [Google Scholar] [CrossRef] [PubMed]
- Dewannieux, M.; Esnault, C.; Heidmann, T. LINE-mediated retrotransposition of marked Alu sequences. Nat. Genet. 2003, 35, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Okada, N.; Hamada, M.; Ogiwara, I.; Ohshima, K. SINEs and LINEs share common 3′ sequences: A review. Gene 1997, 205, 229–243. [Google Scholar] [CrossRef]
- Kajikawa, M.; Okada, N. LINEs Mobilize SINEs in the Eel through a Shared 3′ Sequence. Cell 2002, 111, 433–444. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Churakov, G.; Jurka, J.; Brosius, J.; Schmitz, J. Evolutionary history of 7SL RNA-derived SINEs in Supraprimates. Trends Genet. TIG 2007, 23, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Houck, C.M.; Rinehart, F.P.; Schmid, C.W. A ubiquitous family of repeated DNA sequences in the human genome. J. Mol. Biol. 1979, 132, 289–306. [Google Scholar] [CrossRef]
- Ullu, E.; Tschudi, C. Alu sequences are processed 7SL RNA genes. Nature 1984, 312, 171–172. [Google Scholar] [CrossRef] [PubMed]
- Quentin, Y. Origin of the Alu family: A family of Alu-like monomers gave birth to the left and the right arms of the Alu elements. Nucleic Acids Res. 1992, 20, 3397–3401. [Google Scholar] [CrossRef] [PubMed]
- Dewannieux, M.; Heidmann, T. Role of poly(A) tail length in Alu retrotransposition. Genomics 2005, 86, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Deininger, P. Alu elements: Know the SINEs. Genome Biol. 2011, 12, 236. [Google Scholar] [CrossRef] [PubMed]
- Jurka, J.; Smith, T. A fundamental division in the Alu family of repeated sequences. Proc. Natl. Acad. Sci. USA 1988, 85, 4775–4778. [Google Scholar] [CrossRef] [PubMed]
- Batzer, M.A.; Deininger, P.L.; Hellmann-Blumberg, U.; Jurka, J.; Labuda, D.; Rubin, C.M.; Schmid, C.W.; Ziętkiewicz, E.; Zuckerkandl, E. Standardized nomenclature for Alu repeats. J. Mol. Evol. 1996, 42, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.A.; Keller, H.; Mills, R.E.; Schmidt, S.; Moran, J.V.; Weichenrieder, O.; Devine, S.E. Active Alu retrotransposons in the human genome. Genome Res. 2008, 18, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, L.; Gonder, M.K.; Azrak, S.; Ray, D.A.; Batzer, M.A.; Tishkoff, S.A.; Liang, P. Whole genome computational comparative genomics: A fruitful approach for ascertaining Alu insertion polymorphisms. Gene 2006, 365, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Price, A.L.; Eskin, E.; Pevzner, P.A. Whole-genome analysis of Alu repeat elements reveals complex evolutionary history. Genome Res. 2004, 14, 2245–2252. [Google Scholar] [CrossRef] [PubMed]
- Van de Lagemaat, L.N.; Gagnier, L.; Medstrand, P.; Mager, D.L. Genomic deletions and precise removal of transposable elements mediated by short identical DNA segments in primates. Genome Res. 2005, 15, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Piskurek, O.; Okada, N. Poxviruses as possible vectors for horizontal transfer of retroposons from reptiles to mammals. Proc. Natl. Acad. Sci. USA 2007, 104, 12046–12051. [Google Scholar] [CrossRef] [PubMed]
- Stoneking, M.; Fontius, J.J.; Clifford, S.L.; Soodyall, H.; Arcot, S.S.; Saha, N.; Jenkins, T.; Tahir, M.A.; Deininger, P.L.; Batzer, M.A. Alu Insertion Polymorphisms and Human Evolution: Evidence for a Larger Population Size in Africa. Genome Res. 1997, 7, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, M.; Rooney, A.P.; Okada, N. Phylogenetic relationships among cetartiodactyls based on insertions of short and long interpersed elements: Hippopotamuses are the closest extant relatives of whales. Proc. Natl. Acad. Sci. USA 1999, 96, 10261–10266. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, H.; Maruyama, S.; Okada, N. Retroposon analysis and recent geological data suggest near-simultaneous divergence of the three superorders of mammals. Proc. Natl. Acad. Sci. USA 2009, 106, 5235–5240. [Google Scholar] [CrossRef] [PubMed]
- Shedlock, A.M.; Okada, N. SINE insertions: Powerful tools for molecular systematics. BioEssays News Rev. Mol. Cell. Dev. Biol. 2000, 22, 148–160. [Google Scholar] [CrossRef]
- Nikaido, M.; Piskurek, O.; Okada, N. Toothed whale monophyly reassessed by SINE insertion analysis: The absence of lineage sorting effects suggests a small population of a common ancestral species. Mol. Phylogenet. Evol. 2007, 43, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Rishishwar, L.; Wang, L.; Clayton, E.A.; Mariño-Ramírez, L.; McDonald, J.F.; Jordan, I.K. Population and clinical genetics of human transposable elements in the (post) genomic era. Mob. Genet. Elem. 2017, 7, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hancks, D.C.; Kazazian, H.H. Roles for retrotransposon insertions in human disease. Mob. DNA 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Loftus, A.; Murphy, G.; Brown, H.; Montgomery, A.; Tabak, J.; Baus, J.; Carroll, M.; Green, A.; Sikka, S.; Sinha, S. Development and validation of InnoQuant® HY, a system for quantitation and quality assessment of total human and male DNA using high copy targets. Forensic Sci. Int. Genet. 2017, 29, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.L.; Roy-Engel, A.M.; Nguyen, S.V.; Salem, A.H.; Vogel, E.; Vincent, B.; Myers, J.; Ahmad, Z.; Nguyen, L.; Sammarco, M.; et al. Large-scale analysis of the Alu Ya5 and Yb8 subfamilies and their contribution to human genomic diversity. J. Mol. Biol. 2001, 311, 17–40. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.B.; Salem, A.H.; Hedges, D.J.; Keegan, C.N.; Kimball, B.; Walker, J.A.; Watkins, W.S.; Jorde, L.B.; Batzer, M.A. Genome-wide analysis of the human Alu Yb-lineage. Hum. Genom. 2004, 1, 167–178. [Google Scholar] [CrossRef]
- Otieno, A.C.; Carter, A.B.; Hedges, D.J.; Walker, J.A.; Ray, D.A.; Garber, R.K.; Anders, B.A.; Stoilova, N.; Laborde, M.E.; Fowlkes, J.D.; et al. Analysis of the human Alu Ya-lineage. J. Mol. Biol. 2004, 342, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Rishishwar, L.; Tellez Villa, C.E.; Jordan, I.K. Transposable element polymorphisms recapitulate human evolution. Mob. DNA 2015, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Sajjadian, S.; Eichler, E.E. Limitations of next-generation genome sequence assembly. Nat. Methods 2011, 8, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Tucker, B.A.; Scheetz, T.E.; Mullins, R.F.; DeLuca, A.P.; Hoffmann, J.M.; Johnston, R.M.; Jacobson, S.G.; Sheffield, V.C.; Stone, E.M. Exome sequencing and analysis of induced pluripotent stem cells identify the cilia-related gene male germ cell-associated kinase (MAK) as a cause of retinitis pigmentosa. Proc. Natl. Acad. Sci. USA 2011, 108, E569–E576. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Salzberg, S.L. Repetitive DNA and next-generation sequencing: Computational challenges and solutions. Nat. Rev. Genet. 2011, 13, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Huddleston, J.; Ranade, S.; Malig, M.; Antonacci, F.; Chaisson, M.; Hon, L.; Sudmant, P.H.; Graves, T.A.; Alkan, C.; Dennis, M.Y.; et al. Reconstructing complex regions of genomes using long-read sequencing technology. Genome Res. 2014, 24, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Hofreiter, M.; Paijmans, J.L.A.; Goodchild, H.; Speller, C.F.; Barlow, A.; Fortes, G.G.; Thomas, J.A.; Ludwig, A.; Collins, M.J. The future of ancient DNA: Technical advances and conceptual shifts. BioEssays News Rev. Mol. Cell. Dev. Biol. 2015, 37, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, T.; Shojo, H.; Tanaka, M.; Nambiar, P.; Minaguchi, K.; Umetsu, K.; Adachi, N. Multiplex APLP System for High-Resolution Haplogrouping of Extremely Degraded East-Asian Mitochondrial DNAs. PLoS ONE 2016, 11, e0158463. [Google Scholar] [CrossRef] [PubMed]
- Seidenberg, V.; Schilz, F.; Pfister, D.; Georges, L.; Fehren-Schmitz, L.; Hummel, S. A new miniSTR heptaplex system for genetic fingerprinting of ancient DNA from archaeological human bone. J. Archaeol. Sci. 2012, 39, 3224–3229. [Google Scholar] [CrossRef]
- Singh, G.; Talwar, I.; Sandhu, H.S.; Matharoo, K.; Bhanwer, A.J.S. Genetic dissection of five ethnic groups from Punjab, North-West India—A study based on Autosomal Markers. Leg. Med. 2017, 26, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kothe, M.; Seidenberg, V.; Hummel, S.; Piskurek, O. Alu SINE analyses of 3000-year-old human skeletal remains: A pilot study. Mob. DNA 2016, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Flindt, S. Die Lichtensteinhöhle bei Osterode, Landkreis Osterode am Harz: Eine Opferhöhle der jüngeren Bronzezeit im Gipskarst des südwestlichen Harzrandes: Forschungsgeschite und erste Grabungsergebnisse. Die Kunde N. F. 1996, 47, 435–466. [Google Scholar]
- Flindt, S.; Hummel, S. Die Lichtensteinhöhle: Bestattungsplatz einer Großfamilie aus der Bronzezeit; HöhlenErlebnisZentrum: Bad Grund, Germany, 2014. [Google Scholar]
- Höss, M.; Jaruga, P.; Zastawny, T.H.; Dizdaroglu, M.; Paabo, S. DNA Damage and DNA Sequence Retrieval from Ancient Tissues. Nucleic Acids Res. 1996, 24, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.; Hummel, S.; Hermann, B.; Henke, W. DNA preservation: A microsatellite-DNA study on ancient skeletal remains. Electrophoresis 1999, 20, 1722–1728. [Google Scholar] [CrossRef]
- Seidenberg, V. Ein Bronzezeitlicher Familienclan als Genetisches Archiv—Morphologisch-Paläogenetische Bearbeitung des Skelettkollektivs aus der Lichtensteinhöhle. Ph.D. Thesis, Georg-August-Universität, Göttingen, Germany, 2016. [Google Scholar]
- Frischalowski, M. Examinaton of DNA Preservation in the Skeletal Remains from a Historic Burial Site in Eldagsen/Lower Saxony. Master’s Thesis, Georg-August-Universität, Göttingen, Germany, 2014. [Google Scholar]
- Pepperl, J. Immungenetische Marker im Wandel der Zeit. Ph.D. Dissertation, Georg-August-Universität, Göttingen, Germany, 2008. [Google Scholar]
- Smit, A.F.A.; Hubley, R.; Green, P. Repeatmasker. 2013. Available online: http://www.repeatmasker.org (accessed on 27 July 2016).
- Hummel, S. Ancient DNA Typing: Methods, Strategies and Applications; Springer: Berlin, Germany, 2003; ISBN 3-662-05050-1. [Google Scholar]
- Nelson, D.L.; Ledbetter, S.A.; Corbo, L.; Victoria, M.F.; Ramírez-Solis, R.; Webster, T.D.; Ledbetter, D.H.; Caskey, C.T. Alu polymerase chain reaction: A method for rapid isolation of human-specific sequences from complex DNA sources. Proc. Natl. Acad. Sci. USA 1989, 86, 6686–6690. [Google Scholar] [CrossRef] [PubMed]
- Qiagen. Qiagen QIAquick Gel Extraction Kit. QIAquick® Spin Handbook; Qiagen: Hilden, Germany, 2015. [Google Scholar]
- New England Biolabs® Inc. Monarch® DNA Gel Extraction Kit: Instruction Manual; New England Biolabs® Inc.: Ipswich, MA, USA, 2017. [Google Scholar]
- Macherey & Nagel. Macherey-Nagel NucleoSEQ®: Clean-Up of Sequencing Reactions User Manual; Macherey & Nagel: Düren, Germany, 2017. [Google Scholar]
- Hall BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98.
- Bénit, L.; Lallemand, J.-B.; Casella, J.-F.; Philippe, H.; Heidmann, T. ERV-L Elements: A Family of Endogenous Retrovirus-Like Elements Active throughout the Evolution of Mammals. J. Virol. 1999, 73, 3301–3308. [Google Scholar] [PubMed]
- Churakov, G.; Grundmann, N.; Kuritzin, A.; Brosius, J.; Makalowski, W.; Schmitz, J. A novel web-based TinT application and the chronology of the Primate Alu retroposon activity. BMC Evol. Biol. 2010, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Hummel, S.; Herrmann, B. Evidence of contamination in PCR laboratory disposables. Naturwissenschaften 1995, 82, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.; Whitaker, J.; Flaxman, C.; Brown, N.; Buckleton, J. An investigation of the rigor of interpretation rules for STRs derived from less than 100 pg of DNA. Forensic Sci. Int. 2000, 112, 17–40. [Google Scholar] [CrossRef]
- Butler, J. Advanced Topics in Forensic DNA Typing: Interpretation; Elsevier: Boston, MA, USA, 2014; ISBN 978-0-12-405213-0. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Alu | + | + (Int) | − | Forward Primer 5′→3′ | Reverse Primer 5′→3′ |
|---|---|---|---|---|---|
| Alu duplex 1 | |||||
| 9 | 460 | 161 | 133 | CTAACAGGAATCACTGCTAGTCAGA | CAAGGACATAGTCACAGAACAGATG |
| 34 | 437 | 179 | 101 | CTCCAACAGTCAAAYTCTTCAAATCT | GGGCAATGGGAAAATAAGACAG |
| Alu duplex 2 | |||||
| 19 | 448 | 155 | 134 | GGGTGGAGAGAAAGGTCATTAT | ATGTCAGTAGCTTTCACGAACAA |
| 35 | 438 | 186 | 116 | GGATATGTTTACTTGCTGACTGTCAAT | TCAGACAAGCAAGAATCATAACAGAA |
| Internal Alu primer | AGTAGAGACGGGGTTTCACCGTT | ||||
| Alu Duplex | Alu Element | DO 1911 | DO 3756 | DO 905.01 | DO 2255.03 | DO 58.08 | DO 1548.01 | DO 66.02 | DO 1471 | DO 58.13 | DO 187 | DO 902.01 | DO 1076 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Duplex 1 | Alu 9 | +/− | +/− | +/− | + | + | + | +/− | (+) | n.b. | n.b. | + | +/− |
| Alu 34 | +/− | +/− | + | +/− | +/− | +/− | + | +/− | +/− | +/− | + | + | |
| Duplex 2 | Alu 19 | + | + | + | + | n.b. | + | + | + * | (+) | n.b. | + | + |
| Alu 35 | + | + | + | + | n.b. | + | + | + * | n.b. | n.b. | + | + |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haß, F.; Hummel, S.; Piskurek, O. Duplex Alu Screening for Degraded DNA of Skeletal Human Remains. Diversity 2017, 9, 48. https://doi.org/10.3390/d9040048
Haß F, Hummel S, Piskurek O. Duplex Alu Screening for Degraded DNA of Skeletal Human Remains. Diversity. 2017; 9(4):48. https://doi.org/10.3390/d9040048
Chicago/Turabian StyleHaß, Fabian, Susanne Hummel, and Oliver Piskurek. 2017. "Duplex Alu Screening for Degraded DNA of Skeletal Human Remains" Diversity 9, no. 4: 48. https://doi.org/10.3390/d9040048
APA StyleHaß, F., Hummel, S., & Piskurek, O. (2017). Duplex Alu Screening for Degraded DNA of Skeletal Human Remains. Diversity, 9(4), 48. https://doi.org/10.3390/d9040048