Using Molecular-Assisted Alpha Taxonomy to Better Understand Red Algal Biodiversity in Bermuda

{kind=link}
Abstract
:1. Introduction
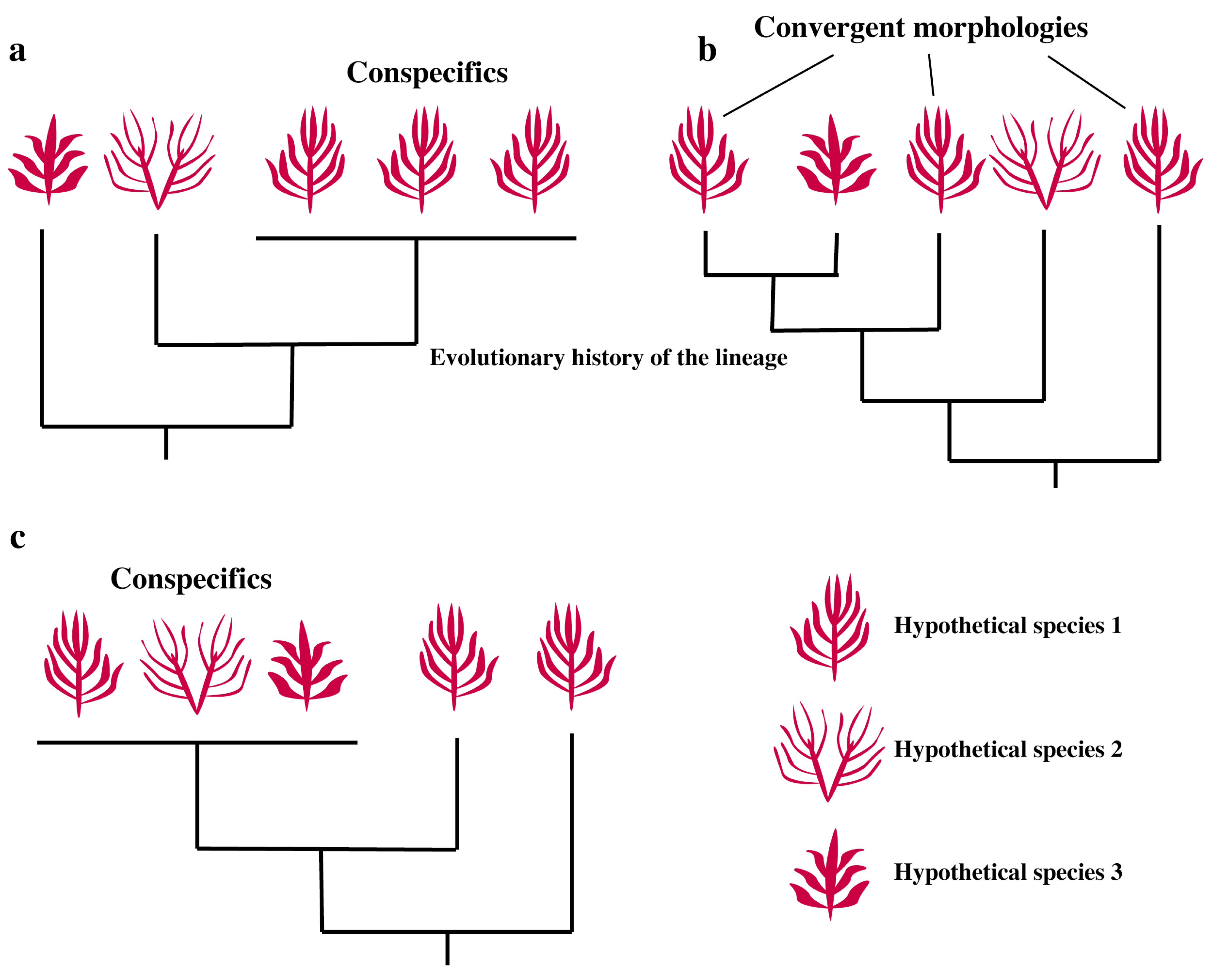
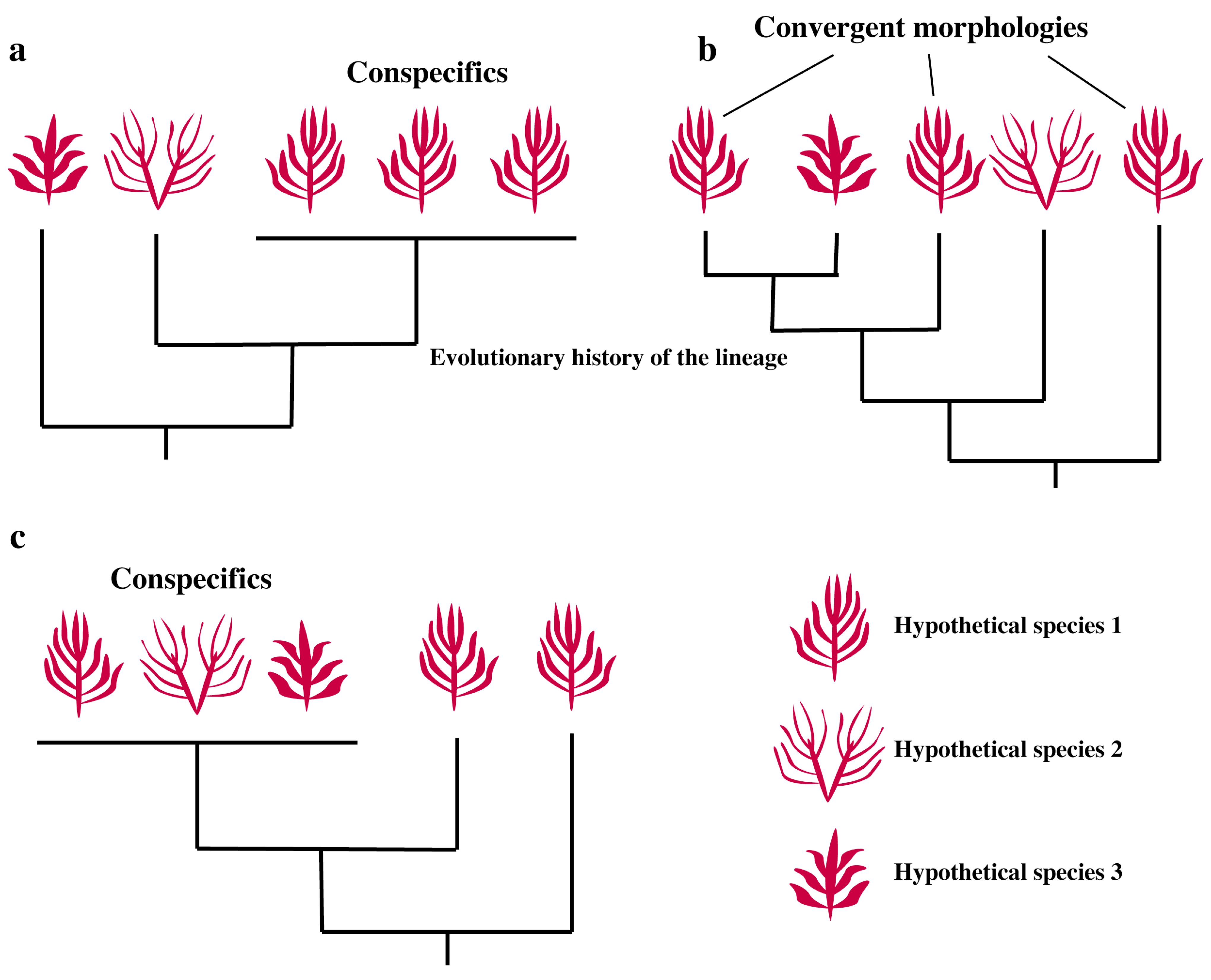
2. Convergence, Hidden Diversity and Sorting It Out
2.1. Convergence
2.2. Hidden Diversity
2.3. Morphological Plasticity
2.4. Remaining Barriers to Sorting It All Out
3. Conclusions
Acknowledgements
References
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, 2229–2238. [Google Scholar]
- Briggs, J.C. Marine Zoogeography; McGraw-Hill: New York, NY, USA, 1974; pp. 1–475. [Google Scholar]
- Lüning, K. Seaweeds. Their Environment, Biogeography, and Ecophysiology; John Wiley & Sons: New York, NY, USA, 1990; pp. 1–527. [Google Scholar]
- Ekman, S. Zoogeography of the Sea; Sidgwick & Jackson Ltd.: London, UK, 1953; pp. 1–417. [Google Scholar]
- Wynne, M.J. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: second revision. Nova Hedw. Beif. 2005, 129, 1–152. [Google Scholar]
- Searles, R.B.; Schneider, C.W. Observations on the deep water flora of Bermuda. Hydrobiologia 1987, 151-152, 261–266. [Google Scholar] [CrossRef]
- Schneider, C.W.; Searles, R.B. Notes on the marine algae of the Bermudas. 3. Avrainvillea sylvearleae, Discosporangium mesarthrocarpum and Peyssonnelia valentinii. J. Phycol. 1998, 34, 180–188. [Google Scholar]
- Schneider, C.W. An annotated checklist and bibliography of the marine macroalgae of the Bermuda islands. Nova Hedw. 2003, 76, 275–361. [Google Scholar] [CrossRef]
- Collins, F.S.; Hervey, A.B. The algae of Bermuda. Proc. Am. Acad. Arts Sci. 1917, 53, 1–195. [Google Scholar]
- Howe, M.A. Algae. In Flora of Bermuda; Britton, N.L., Ed.; Charles Scribner’s Sons: New York, NY, USA, 1918; pp. 489–540. [Google Scholar]
- Taylor, W.R. Survey of the marine algae of Bermuda. Yearbook Am. Phil. Soc. 1952, 1951, 167–171. [Google Scholar]
- Taylor, W.R. Marine Algae of the Eastern Tropical and Subtropical Coasts of the Americas; University of Michigan Press: Ann Arbor, MI, USA, 1960; pp. 1–879. [Google Scholar]
- Taylor, W.R.; Bernatowicz, A.J. Distribution of marine algae about Bermuda. Berm. Biol. Sta. Res. Spec. Publ. 1969, 1, 1–42. [Google Scholar]
- Schneider, C.W. Notes on the marine algae of the Bermudas. 6. Some rare or newly reported Ceramiales (Rhodophyta), including Crouania elisiae sp. nov. Phycologia 2004, 13, 563–578. [Google Scholar] [CrossRef]
- Schneider, C.W.; Lane, C.E. Notes on the marine algae of the Bermudas. 7. Additions to the flora including Chondracanthus saundersii sp. nov. (Rhodophyta, Gigartinaceae) based on rbcL sequence analysis. Phycologia 2005, 44, 72–83. [Google Scholar] [CrossRef]
- Schneider, C.W.; Lane, C.E. Notes on the marine algae of the Bermudas. 8. Further additions to the flora, including Griffithsia aestivana sp. nov. (Ceramiaceae, Rhodophyta) and an update on the alien Cystoseira compressa (Sargassaceae, Heterokontophyta). Bot. Mar. 2007, 50, 128–140. [Google Scholar]
- Schneider, C.W.; Lane, C.E. Notes on the marine algae of the Bermudas. 9. The genus Botryocladia (Rhodophyta, Rhodymeniaceae), including B. bermudana, B. exquisita and B. flookii spp. nov. Phycologia 2008, 47, 614–629. [Google Scholar] [CrossRef]
- Schneider, C.W.; Lane, C.E.; Saunders, G.W. Crassitegula walsinghamii (Sebdeniaceae, Halymeniales), a new red algal genus and species from Bermuda based upon morphology and SSU rDNA sequence analyses. Eur. J. Phycol. 2006, 41, 115–124. [Google Scholar] [CrossRef]
- Schneider, C.W.; Lane, C.E.; Saunders, G.W. Notes on the marine algae of the Bermudas. 11. More additions to the benthic flora and a phylogenetic assessment of Halymenia pseudofloresii Collins & M. Howe (Halymeniales, Rhodophyta) from its type locality. Phycologia 2010, 49, 154–168. [Google Scholar] [CrossRef]
- Schneider, C.W.; Wynne, M.J. Notes on the marine algae of the Bermudas. 10. Woelkerlingia sterreri sp. nov. (Rhodophyta, Ceramiaceae), a first record of the genus in the western Atlantic. Carib. J. Sci. 2009, 44, 301–310. [Google Scholar]
- Saunders, G.W.; Lane, C.E.; Schneider, C.W.; Kraft, G.T. Unraveling the Asteromenia peltata species complex with clarification of the genera Halichrysis and Drouetia (Rhodymeniaceae, Rhodophyta). Can. J. Bot. 2006, 84, 1581–1607. [Google Scholar] [CrossRef]
- Hommersand, M.H.; Guiry, M.D.; Fredericq, S.; Leister, G.L. New perspectives in the taxonomy of the Gigartinaceae (Gigartinales, Rhodophyta). Hydrobiologia 1993, 260-261, 105–120. [Google Scholar] [CrossRef]
- Hommersand, M.H.; Fredericq, S.; Freshwater, D.W.; Hughey, J. Recent developments in the systematics of the Gigartinaceae (Gigartinales, Rhodophyta) based on rbcL sequence analysis and morphological evidence. Phycol. Res. 1999, 47, 139–151. [Google Scholar] [CrossRef]
- Ballantine, D.L.; Aponte, N.E. Botryocladia bahamense sp. nov. (Rhodymeniaceae, Rhodophyta) from the Bahamas, western Atlantic. Crypt. Algol. 2002, 23, 123–130. [Google Scholar]
- Afonso-Carrillo, J.; Sobrino, C. Vegetative and reproductive morphology of Botryocladia botryoides, B. occidentalis and B. canariensis sp. nov. (Rhodymeniaceae, Rhodophyta) from the Canary Islands. Phycologia 2003, 42, 138–150. [Google Scholar] [CrossRef]
- Gavio, B.; Fredericq, S. Botryocladia caraibica (Rhodymeniales, Rhodophyta), a new species from the Caribbean. Cryptogam. Algol. 2003, 24, 93–106. [Google Scholar]
- Gavio, B.; Fredericq, S. New species and new records of offshore members of the Rhodymeniales (Rhodophyta) in the northern Gulf of Mexico. Gulf Mex. Sci. 2005, 23, 58–83. [Google Scholar]
- Saunders, G.W.; Strachan, I.M.; Kraft, G.T. The families of the order Rhodymeniales (Rhodophyta): a molecular-systematic investigation with a description of Faucheaceae fam. nov. Phycologia 1999, 38, 23–40. [Google Scholar] [CrossRef]
- Huisman, J.M.; Millar, A.J.K. Asteromenia (Rhodymeniaceae, Rhodymeniales), a new red algal genus based on Fauchea peltat. J. Phycol. 1996, 32, 138–145. [Google Scholar]
- Taylor, W.R. Caribbean marine algae of the Allan Hancock Expedition, 1939. Allan Hancock Atl. Exp. Rep. 1942, 2, 1–193. [Google Scholar]
- Le Gall, L.; Dalen, J.L.; Saunders, G.W. Phylogenetic analyses of the red algal order Rhodymeniales supports recognition of the Hymenocladiaceae fam. nov., Fryeellaceae fam. nov., and Neogastroclonium gen. nov. J. Phycol. 2008, 44, 1556–1571. [Google Scholar] [CrossRef]
- Won, B.Y.; Cho, T.O.; Fredericq, S. Morphological and molecular characterization of species of the genus Centroceras (Ceramiaceae, Ceramiales), including two new species. J. Phycol. 2009, 45, 227–250. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; www electronic publication, National University of Ireland: Galway, Ireland, 2010. Available online: http://www.algaebase.org (accessed on Mar 1, 2010).
- Kunth, C.S. Synopsis plantarum, quas, in itinere ad plagam aequinoctialem orbis novi, collegerunt Al. de Humboldt et Am. Bonpland; F.G. Levrault: Parisiis [Paris], France, 1822; Vol. 1, pp. 1–491. [Google Scholar]
- Kützing, F.T. Tabulae phycologicae. Vol. 13; Gedruckt auf kosten des Verfassers: Nordhausen, Germany, 1863; pp. 1-31,100. [Google Scholar]
- Collins, F.S.; Howe, M.A. Notes on species of Halymenia. Bull. Torrey Bot. Club 1916, 43, 169–182. [Google Scholar]
- Taylor, W.R. Notes on three Bermudian marine algae. Hydrobiologia 1961, 18, 277–283. [Google Scholar] [CrossRef]
- Schneider, C.W. North Carolina marine algae. V. Additions to the flora of Onslow Bay, including the reassignment of Fauchea peltata Taylor to Weberella Schmitz. Brit. Phycol. 1975, 10, 129–138. [Google Scholar] [CrossRef]
- Taylor, W.R. Notes on algae from the tropical Atlantic Ocean—VII. Rev. Algol. Nouv. Sér. 1974, 11, 58–71. [Google Scholar]
- Littler, D.S.; Littler, M.M. Caribbean Reef Plants. An Identification Guide to the Reef Plants of the Caribbean, Bahamas, Florida and Gulf of Mexico; OffShore Graphics, Inc.: Washington, DC, USA, 2000; pp. 1–542. [Google Scholar]
- Kucera, H.; Saunders, G.W. Assigning morphological variants of Fucus (Fucales, Phaeophyceae) in Canadian waters to recognized species using DNA barcoding. Botany 2008, 86, 1065–1079. [Google Scholar] [CrossRef]
- Saunders, G.W. Routine DNA barcoding of Canadian Gracilariales (Rhodophyta) reveals the invasive species Gracilaria vermiculophylla in British Columbia. Mol. Ecol. Resour. 2009, 9, 140–150. [Google Scholar] [CrossRef]
- Walker, R.H.; Brodie, J.; Russell, S.; Irvine, L.M.; Orfanidis, S. Biodiversity of coralline algae in the northeastern Atlantic including Corallina caespitosa sp. nov. (Corallinoideae, Rhodophyta). J. Phycol. 2009, 45, 287–297. [Google Scholar] [CrossRef]
- Wiemers, M.; Fiedler, K. Does the DNA barcoding gap exist?—A case study in blue butterflies (Lepidoptera: Lycaenidae). Front. Zool. 2007, 4, 8. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cianciola, E.N.; Popolizio, T.R.; Schneider, C.W.; Lane, C.E. Using Molecular-Assisted Alpha Taxonomy to Better Understand Red Algal Biodiversity in Bermuda. Diversity 2010, 2, 946-958. https://doi.org/10.3390/d2060946
Cianciola EN, Popolizio TR, Schneider CW, Lane CE. Using Molecular-Assisted Alpha Taxonomy to Better Understand Red Algal Biodiversity in Bermuda. Diversity. 2010; 2(6):946-958. https://doi.org/10.3390/d2060946
Chicago/Turabian StyleCianciola, Elisabeth N., Thea R. Popolizio, Craig W. Schneider, and Christopher E. Lane. 2010. "Using Molecular-Assisted Alpha Taxonomy to Better Understand Red Algal Biodiversity in Bermuda" Diversity 2, no. 6: 946-958. https://doi.org/10.3390/d2060946
APA StyleCianciola, E. N., Popolizio, T. R., Schneider, C. W., & Lane, C. E. (2010). Using Molecular-Assisted Alpha Taxonomy to Better Understand Red Algal Biodiversity in Bermuda. Diversity, 2(6), 946-958. https://doi.org/10.3390/d2060946