Abstract
As part of an overall decline in biodiversity, populations of many organisms are declining and species are being lost at unprecedented rates around the world. This includes many populations and species of amphibians. Although numerous factors are affecting amphibian populations, we show potential direct and indirect effects of climate change on amphibians at the individual, population and community level. Shifts in amphibian ranges are predicted. Changes in climate may affect survival, growth, reproduction and dispersal capabilities. Moreover, climate change can alter amphibian habitats including vegetation, soil, and hydrology. Climate change can influence food availability, predator-prey relationships and competitive interactions which can alter community structure. Climate change can also alter pathogen-host dynamics and greatly influence how diseases are manifested. Changes in climate can interact with other stressors such as UV-B radiation and contaminants. The interactions among all these factors are complex and are probably driving some amphibian population declines and extinctions.
1. Introduction
Large losses in biodiversity are occurring around the world [1]. Though the exact number of species being lost is unknown, some estimate that the rate of extinction is greater than any known in the last 100,000 years [2]. This so called “biodiversity crisis” is exemplified by population declines, range reductions and extinctions of amphibian species around the world [3,4,5,6,7]. In at least some regions, amphibian losses appear to be more severe than losses in other vertebrate taxa [4,8,9]. From an evolutionary historical perspective, amphibians may be part of a sixth major extinction event [10].
There appears to be no single cause for amphibian population declines. Like other animals, amphibians are affected by numerous environmental stresses that often act in complex ways [11]. Such stressors include habitat alteration and destruction, environmental contamination, introduced exotic species, disease, increasing ultraviolet-B radiation and climate change. The causes for the decline of a given species may be different from region to region and even in different populations of the same species. There may be synergistic interactions between more than one factor. There may be interspecific differences and even differences between life stages in how amphibians react to these stresses.
The emphasis of our review is on how climate change may affect amphibians at the individual, population and even community level. There are often complex and even synergistic interactions among climatic factors affecting amphibians. Nevertheless, we attempt to categorize these factors as direct and indirect, realizing that this is often a simplistic categorization involving abiotic and biotic components. We also summarize how climate change may interact with other stressors such as contaminants, ultraviolet radiation and diseases. Because numerous declines of amphibian populations are well documented, we believe the potential effects of climate change at the population level are especially important to document and discuss.
2. Discussion
2.1. Climate Perspective
In the 1970s, environmental biologists and atmospheric scientists predicted that two significant human-induced environmental changes, global warming and ozone depletion, could potentially affect the biology of a wide array of plants, animals and microorganisms. Increased emissions of "greenhouse" gases resulting from burning fossil fuels and land conversion were projected to cause a significant rise in global temperatures in the coming decades. Moreover, it was shown that chlorofluorocarbons (CFCs) and other commonly used industrial gases, were depleting the earth's protective ozone layer, increasing the amount of cell-damaging ultraviolet-B (UV-B; 280−315 nm) radiation that reaches the Earth’s surface [12]. Scientists projected that species might respond to these global changes by altering their behavior and shifting ranges. However, if they are unable to adapt to these environmental changes, they may experience increased mortality and significant sublethal effects. Additionally, a number of scientists suggested that global warming and ozone depletion will affect entire ecological communities [13,14,15].
It is now abundantly clear that the Earth’s climate is changing in response to anthropogenic greenhouse-gas emissions. Average annual global temperatures have risen 0.7 °C over the last century [16]. The rate of increase has been greatest in the latter part of the century. For example, the rate at which annual average global temperatures increased from 1980 to 2005 was roughly double the rate of increase from 1905 to 2005. Although different parts of the globe have warmed at different rates, at least in the last 25 years, most of the Earth’s surface has warmed. Precipitation patterns have also changed over the last 100 years. These, changes, however, have been less consistent—some areas have experienced decreases in precipitation and some areas experienced increases [16].
The Earth’s climate is projected to change even more dramatically over the next century. Global average annual temperatures are projected to rise between 1.1 and 6.4 °C by 2100 [16]. The largest increases are projected for the high northern latitudes where average annual temperatures may increase more than 7.5 °C (Figure 1a). This projected pattern is much more pronounced for the months that make up the northern hemisphere’s winter (December, January, and February). In June, July, and August, the largest temperature increases are projected for the centers of the continents [16].
Although global mean precipitation is projected to increase by the end of the century, there is far less agreement across the general circulation model projections [17] as to the magnitude, and in some regions the sign of precipitation changes. In addition, there is much more spatial variability in the projections, such that some regions are projected to receive less precipitation and others are projected to receive more (Figure 1b). Total annual precipitation is projected to increase at higher latitudes in both hemispheres. At low- to mid-northern latitudes—specifically Central America, northern Africa, and Europe—and mid-southern latitudes—specifically southern Africa, and parts of Brazil, Chile, Argentina, and Australia—total annual precipitation is projected to decrease (Figure 1b). In some areas, these projected changes in precipitation vary by season. For example, central North America is projected to experience more precipitation in winter months, but less precipitation in summer months. Much of Southeast Asia is projected to see increases in precipitation in June, July, and August, and decreases in December, January, and February. It is important to note, however, that due to the greater variability in the precipitation projections generated by the general circulation models, there is more uncertainty in the precipitation projections than the temperature projections.
The latest United Nations panel on the environment discussed the importance of the link between ozone depletion (with increasing UV-B radiation) and climate [18]. Exposure of organisms to increasing UV-B radiation is often linked to fluctuations in weather and changes in climate [18,19,20,21,22]. This can occur with changes in precipitation and temperature and these changes can affect aquatic organisms, especially because changes in weather and climate can affect hydroperiods. Decreases in stratospheric ozone, climate warming and lake acidification leading to decreases in dissolved organic carbon concentrations [22] all result in increasing levels of UV-B radiation. Levels of UV-B radiation have risen significantly in modern time (especially since 1979) both in the tropics and in temperate regions [23,24,25]. UV-B radiation can cause mutations and cell death [15,21]. At the individual level, UV-B radiation can slow growth rates, cause immune dysfunction, induce sublethal damage and can cause individual mortality [15,21].
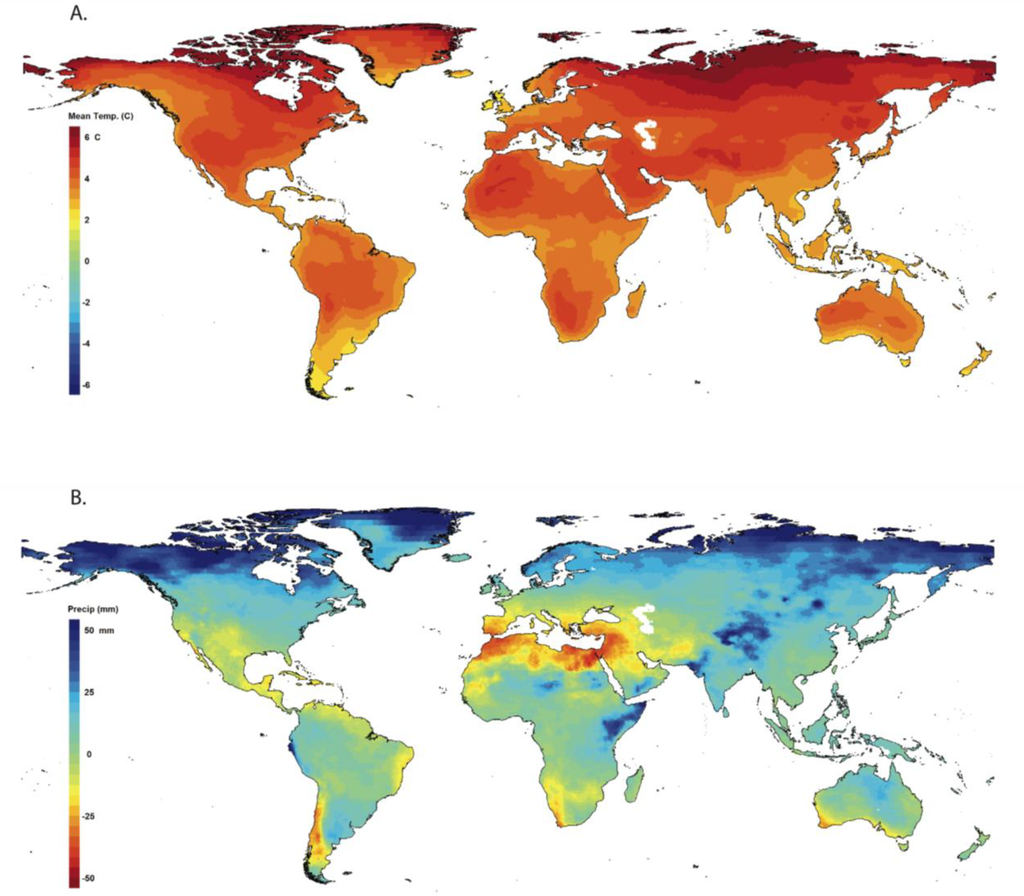
Figure 1.
Projected changes in (A) mean annual temperature, and (B) total annual precipitation. All changes are for an averaged period from 2070 to 2099 relative to an averaged period from 1961 to 1990. Mapped values are the medians of projections from 16 general circulation models run for the SRES A2 emissions scenario. All 16 climate projections were created for the World Climate Research Programme’s (WCRP’s) Coupled Model Intercomparison Project phase 3 (CMIP3) multi-model dataset and used for analyses reported in the IPCC Fourth Assessment Report. The climate projections were downscaled to a 0.5-degree resolution as described in Maurer et al. [223]. The maps were created with the ClimateWizard climate data analysis tool [224].
Figure 1.
Projected changes in (A) mean annual temperature, and (B) total annual precipitation. All changes are for an averaged period from 2070 to 2099 relative to an averaged period from 1961 to 1990. Mapped values are the medians of projections from 16 general circulation models run for the SRES A2 emissions scenario. All 16 climate projections were created for the World Climate Research Programme’s (WCRP’s) Coupled Model Intercomparison Project phase 3 (CMIP3) multi-model dataset and used for analyses reported in the IPCC Fourth Assessment Report. The climate projections were downscaled to a 0.5-degree resolution as described in Maurer et al. [223]. The maps were created with the ClimateWizard climate data analysis tool [224].
Changes in global temperature, precipitation and levels of ultraviolet radiation may contribute to amphibian population declines. Moreover, environmental stressors may act in conjunction with global climate and atmospheric changes to adversely affect amphibian populations. Finally, a number of investigators have proposed that emerging infectious diseases may be stimulated by climate change and changes in the levels of UV-B radiation reaching the earth’s surface (discussed below).
2.2. Direct Effects of Climate Change on Amphibians
2.2.1. Climate change, extreme weather patterns and amphibians
Climate change and other changes in the environment that are global in nature have been recognized as potential problems for amphibian populations by a number of investigators [19,26,27,28,29,30]. Moreover, there is some evidence that short-term changes in weather may affect amphibian population declines. Although weather conditions are more likely to be transient, localized events rather than sustained changes in climate per se, the effects of extreme weather on amphibians may give us some insight on how amphibians may respond to long-term changes in climate. Thus, McMenamin et al. [31] described how increasing temperatures and drought have contributed to loss of amphibian habitats in parts of western North America. The disappearance of several amphibian species in southeastern Brazil in the late 1970s was attributed to unusual frost [32]. Weygoldt [33] attributed population declines of Brazilian frogs in the Atlantic mountains to extremely dry winters. It was suggested that the disappearance of the golden toad (Bufo periglenes) in Costa Rica may have been due to unusual weather conditions [34].
Pounds et al. [9] illustrated the complex interrelationships among global environmental changes and amphibian population declines. They found that changes in water availability associated with changes in large-scale climate processes, such as the El Niño/Southern Oscillation (ENSO) may significantly affect amphibian, reptile and bird populations in the Monte Verde cloud forest of Costa Rica. They showed that dry periods associated with global warming are correlated with amphibian and reptile losses and changes in the bird community. In Costa Rica and potentially in other high altitude tropical sites, global warming appears to have resulted in a decrease in the amount of mist precipitation received in the forest due to increased altitude of the cloud bank.
Changes in ambient temperature may influence amphibian behaviors, including those related to reproduction. Potentially, changes in ambient temperature on a global scale could disrupt the timing of breeding, periods of hibernation, and the ability to find food [26,35].
2.2.2. Range shifts
Predicted changes in the global climate can potentially cause shifts in the geographic ranges of plants and animals. Furthermore, altitudinal shifts with changes in climate have been reported in some regions [36,37]. Recent climatic changes have already resulted in species range shifts [38]. The more extreme changes in climate projected for the coming century [16] will likely produce even larger shifts in species distributions [39,40]. The impacts of these range shifts may have profound effects on both community structure and the functioning of ecosystems [41]. Predictions of future range shifts have relied on a variety of modeling approaches. However, few studies have attempted to model the potential future impacts of climate change on amphibians [39,42]. The studies that have been conducted generally make use of bioclimatic models to predict climate-driven shifts in the potential ranges of species [17]. Bioclimatic models define the current distribution of a species as a function of current climate and then project future potential ranges based on projected future climate data [43,44,45].
Using bioclimatic models, Lawler et al. [44] assessed the relative vulnerability of amphibians to climate change in the western hemisphere. The bioclimatic model projections provide a general indication of where changes in amphibian faunas might be expected to occur over the coming century. Lawler et al. [44] mapped the distributions of 1,099 restricted-range species for which they were unable to build accurate bioclimatic models.
Lawler et al. [44] showed that some of the greatest impacts to amphibian populations will likely occur in Central America (Figure 2). Several areas in this region have high concentrations of restricted-range species and are simultaneously projected to get hotter and drier and to experience high rates of species turnover. Other portions of the hemisphere were projected to experience combinations of these factors. Although the analysis of Lawler et al. [44] included three ways in which amphibians might be vulnerable to climate change, there are several other aspects of climate vulnerability that were not addressed. For example, climate change may indirectly affect amphibians by altering the spread of or the susceptibility to disease and changing phenological relationships.
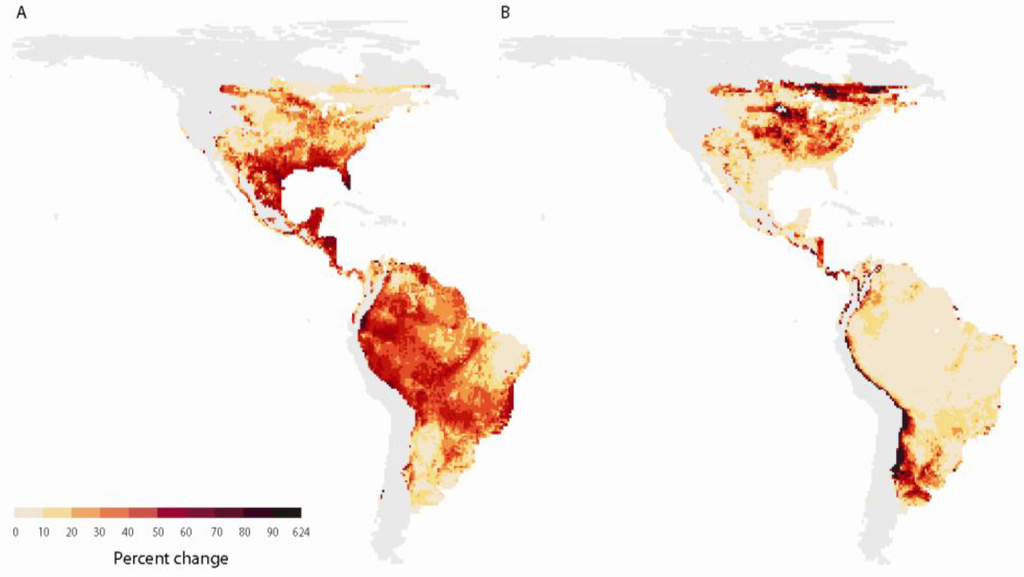
Figure 2.
Predictions of (A) losses and (B) gains in amphibian species as a result of potential climate-driven shifts in species distributions. Predictions were based on 10 different climate projections for a mid-high (A2) greenhouse-gas emission scenario. Eighty percent (8 of 10) of the climate-change projections resulted in greater percent losses (A) and gains (B) in species richness than the values in these maps. The light grey areas indicate where small sample sizes precluded reliable estimates of change. These data and analyses are described in full in Lawler et al. [17].
Figure 2.
Predictions of (A) losses and (B) gains in amphibian species as a result of potential climate-driven shifts in species distributions. Predictions were based on 10 different climate projections for a mid-high (A2) greenhouse-gas emission scenario. Eighty percent (8 of 10) of the climate-change projections resulted in greater percent losses (A) and gains (B) in species richness than the values in these maps. The light grey areas indicate where small sample sizes precluded reliable estimates of change. These data and analyses are described in full in Lawler et al. [17].
2.2.3. Effects of climate change on survival
In amphibians, temperature acts as a controlling factor for many physiological processes, including rates of oxygen uptake, heart rate, locomotion, water balance, digestion, developmental rate, sex determination, and immune function [46]. In addition, temperature can influence the concentration of dissolved oxygen in aquatic habitats, where warmer water generally has lower concentrations of dissolved oxygen. Water balance, which is essentially the interplay between water gain through osmosis and water loss through evaporation, also heavily influences amphibian physiology and behavior. Most amphibian species have highly vascularized permeable skin, which acts as an osmoregulatory and respiratory organ. With the exception of a few “watertight” species, water moves freely across the skin, resulting in high rates of evaporative water loss under dry or windy conditions [47]. Precipitation, humidity, and soil moisture can influence water availability for amphibians, and can therefore influence survival and behavior. Changes in temperature and precipitation will influence the physiology, behavior, and ecology of many amphibian species.
Although the effects of climate change on amphibians are likely to be complex, it is possible to offer some predictions about the direct effects of these changes on amphibians. Alterations to temperature and precipitation regimes may result in mortality events. Changes to these two physical factors may also result in changes in reproduction, development, feeding, dispersal, and immune function. The responses of amphibians will likely be species-specific and perhaps even specific to different populations of the same species. Temperature or desiccation thresholds for an individual will depend on both adaptation and acclimation. Changes in climate may push some individuals or populations past these thresholds, resulting in mortality or sublethal effects such as reduced growth.
The projected changes in temperature and precipitation may increase the chances of mortality for certain species or individuals. Those populations living in sensitive habitats, such as ephemeral ponds [48], coastal wetlands [49], arid and semi-arid systems [50], or alpine areas [51] are likely to see habitat loss or alteration as a result of changes in climate, which in turn may result in population decline or extirpation in these habitats. In addition, species with life history characteristics that make them more susceptible to changes in temperature or precipitation may be more likely to suffer mortality under changing climates. For example, salamanders in the family Plethodontidae lack internal lungs and rely heavily on cutaneous respiration. In general, diffusion of oxygen across the skin requires a moist surface; therefore, these salamanders may be more susceptible to changes in precipitation or temperature which increase rates of evaporative water loss across their skin. Warmer winter temperatures associated with climate change are negatively correlated with body condition, female body size, number of eggs laid, and survival of female common toads (Bufo bufo; [52]). However, with a few notable exceptions [52], evidence for such direct effects of climate change on survival of individual species or populations is rare.
The lack of data on the direct effects of climate change on survival may be due in part to the difficulty in assigning a causative agent for an observed decline. Assigning the causative agent for observed declines or extinctions is necessarily correlative in nature, and thus it is nearly impossible to identify a culprit with certainty. Some evidence exists for correlations between population declines or extinctions and climatic variability or trends ([9,53,54] but see [28,55]). However, the interaction between climate, climate change, and population trends is complex and other factors may be involved with the declines observed. Thus, climate change is suspected to play a role in declining amphibian populations. However, it is unclear whether climate change increases mortality rates directly, affects other aspects of the biology of the organism such as feeding rates, or interacts with other factors such as disease to cause declines in populations.
2.2.4. Effects of climate change on reproduction
Changes in temperature or precipitation have the potential to influence many aspects of amphibian reproduction, including timing of reproduction. Shifts in phenology due to climate change have been observed in several species. For example, Beebee [56] analyzed timing of reproduction in three species of anurans in Britain and found evidence for earlier breeding with shifting climate. Trends towards earlier breeding have been observed for several species of amphibians in Japan [57]. Newts in the genus Triturus also show trends towards earlier breeding with shifts in climate [58]. In addition, Chadwick et al. [58] found that different sexes may respond differently to changing climates. Male Triturus helveticus and T. vulgaris showed a greater degree of change in arrival date compared to females of the same species. Some species appear to breed earlier in response to changing climate, but other species do not, even within the same geographic area. In New York State, four species of anurans show trends towards earlier breeding correlated with changes in climate, while two species of anurans do not show these trends [59]. Amphibian breeding tends to peak at different times in different species. As a result, any one breeding habitat may have different species breeding at different times, with certain species present at similar stages simultaneously. However, shifts in breeding phenology due to climate change that vary among species may result in changes to amphibian assemblages in breeding habitats over time. Species that historically did not develop concurrently may suddenly inhabit the same breeding habitat at the same time, resulting in new biotic environments for the developing larvae.
Regardless of a link with previous or current changes in climate, environmental cues such as temperature and precipitation are clearly linked to onset of reproduction in many species. For example, Blaustein et al. [35] detected a relationship between timing of reproduction and temperature in three species, but not a fourth species. Temperature in February was the best predictor of onset of breeding in Rana dalmantina in Romania [60]. Rainfall events can also be associated with breeding events. In the same population of R. dalmantina, amount of precipitation in February explained 41% of the variance in number of egg masses deposited [60]. Spawning activity of the diurnal toad (Melanophryniscus rubriventris) is closely associated with heavy rainfall events [61]. The environmental cue associated with timing of reproduction may vary depending on the species’ breeding habitat or other life history characteristics. A recent study found that species using permanent ponds had a breeding peak related to temperature, while those using ephemeral ponds responded to precipitation [62]. These data suggest that timing of reproduction is closely linked to environmental cues and supports the assertion that climate change will likely affect the timing of reproduction in many species.
The effects of climate change on reproductive phenology have received the most attention, but other aspects of reproduction are likely to be affected. For example, warmer temperatures are likely to result in lower concentrations of dissolved oxygen (DO) in aquatic habitats. This reduction in oxygen may negatively affect developing embryos and larvae, particularly as increases in temperature increase the oxygen consumption rate in amphibians [46]. Chronic hypoxia can delay development, delay hatching, and result in less developed embryos at hatching in Ambystoma [63]. In addition, reduced oxygen concentrations can result in accelerated hatching in Rana, but at a smaller size [63]. Dissolved oxygen concentrations can also affect behavior of larvae. Larvae inhabiting areas with low DO move to the surface and “gulp” air more frequently than larvae in areas with high DO concentrations [64]. Frequent bobbing to the surface for air may reduce time available for foraging in these larvae, and therefore may reduce rates of growth and development. High temperatures may also affect sex determination in some species. Warm temperatures can result in either masculinization or feminization, depending on the species [65]. A shift towards more males in a population may eventually reduce reproductive rates as females become a limited resource. Changes to water temperatures due to climate change will likely have a myriad of effects on amphibians, and those effects may be complex and unexpected.
2.2.5. Effects of climate change on development
Changes in temperature due to climate change are likely to alter the developmental rates of amphibians. Generally, developmental rate increases with increasing temperature, up to a threshold which may be species- or population- specific [66]. Thermal tolerances can vary by life history stage, but the trend towards faster development with higher temperatures is common across species and life history stages [66].
Thermal environments in the embryonic stage can affect survival and development. For many species, the “tolerance band” of acceptable developmental temperature during the embryonic stage can span 10−20 ˚C. For example, Volpe [67] showed that normal development of Bufo valliceps embryos can occur between 20 and 33 ˚C; however, survival drops steeply on either side of this tolerance band, such that no gastrulation occurs at 36 ˚C. Thus, shifts in temperature that push a population past the tolerance band of normal development may result in years with extremely low or zero survival rates for embryos. Although survival of embryonic life history stages contributes very little to population dynamics in any given year [68,69], repeated years of complete reproductive failure due to changing temperatures may result in population declines. The effects of embryonic developmental temperature can carry over to later life history stages [70], suggesting that thermal environments in the embryonic life history stages can influence population dynamics indirectly.
Survival and developmental rates of larvae are also influenced by temperature. The effects of temperature on survival of larvae are likely to be complex. For example, survival of Pacific treefrog (Pseudacris regilla) larvae under high temperatures is dependent on tadpole density. When tadpole density is low, higher temperatures increase survival rates [71]. However, when tadpole density is high, higher temperatures result in reduced survival [71]. Survival of larvae can influence population dynamics to a larger degree than embryonic survival [68].
The effects of increasing temperature on larval amphibians may include a reduction in time to metamorphosis, a decrease in size at metamorphosis, or both. Moreover, the effects of temperature on development may interact with other factors such as food availability [72] and hydroperiod [73]. Exposure to higher temperatures shortens the larval period in many species [74]. For example, Scaphiopus couchii and Spea multiplicata are desert toads that show accelerated time to metamorphosis with increasing temperature [75]. This pattern of accelerated development has been observed in both anurans [72,76,77,78] and urodels [79,80]. Shorter larval periods can increase chances of survival in environments such as ephemeral ponds and streams by increasing the chance of successful emergence from a pond that is drying. For many species, however, a reduction in larval period also results in metamorphosis at a smaller size [66,74,81,82]. This pattern suggests a likely trade-off between rate of development and growth, which might be exacerbated by climate change. Trade-offs between development and other important factors related to fitness such as post-metamorphic performance [83,84,85] and immune function [86] have also been observed. As with embryonic development, crossing thermal tolerance thresholds can lead to negative effects of high temperatures. For example, exposure to extremely high temperatures (>32 ˚C) can paradoxically reduce the number of individuals successfully completing metamorphosis [87]. These effects of increasing temperature can influence survival and reproduction, thereby influencing population dynamics.
2.2.6. Effects of climate change on behavior
Climate change may also influence behaviors such as foraging and dispersal. Under stressful conditions (high temperatures and/or low humidity), amphibians may seek refugia and reduce activity [88]. Amphibians may alter their dispersal, migration, or other movements under increased temperatures and reduced humidity. For example, Rana aurora adults avoided clear cut areas when precipitation was low [89], suggesting that low humidity in the clear cuts made these areas impermeable for adult amphibians. Adult and juvenile red-spotted newts (Notophthalmus viridescens) avoid leaving refuge during hot and dry climatic conditions, and those individuals that left refuge traveled shorter distances after dry days [90]. These responses are species-specific, where individuals and species from drier habitats are more resistant to desiccation [91]. Small bodied amphibians are more vulnerable to desiccation in hot or dry environments [91], suggesting that newly metamorphosed individuals and adults of smaller body size may be more susceptible to mortality from changes in climate. A reduction in time spent foraging may in turn reduce the available resources for growth and reproduction and thus may reduce the chances of survival. Similarly, changes in migration or dispersal distances may reduce gene flow between populations and reduce the likelihood of rescue by immigration after local population crashes.
2.3. Indirect Effects of Climate Change on Amphibians
Long-term changes in temperature and precipitation will likely affect amphibians through a variety of indirect pathways (Figure 3). Potential changes include impacts on terrestrial and aquatic habitats, food webs and other community-level interactions (e.g., competition), the spread of diseases, and the interplay among these factors. These changes can influence the dynamics of species occurrence across a landscape and can ultimately lead to range shifts of entire assemblages [37].
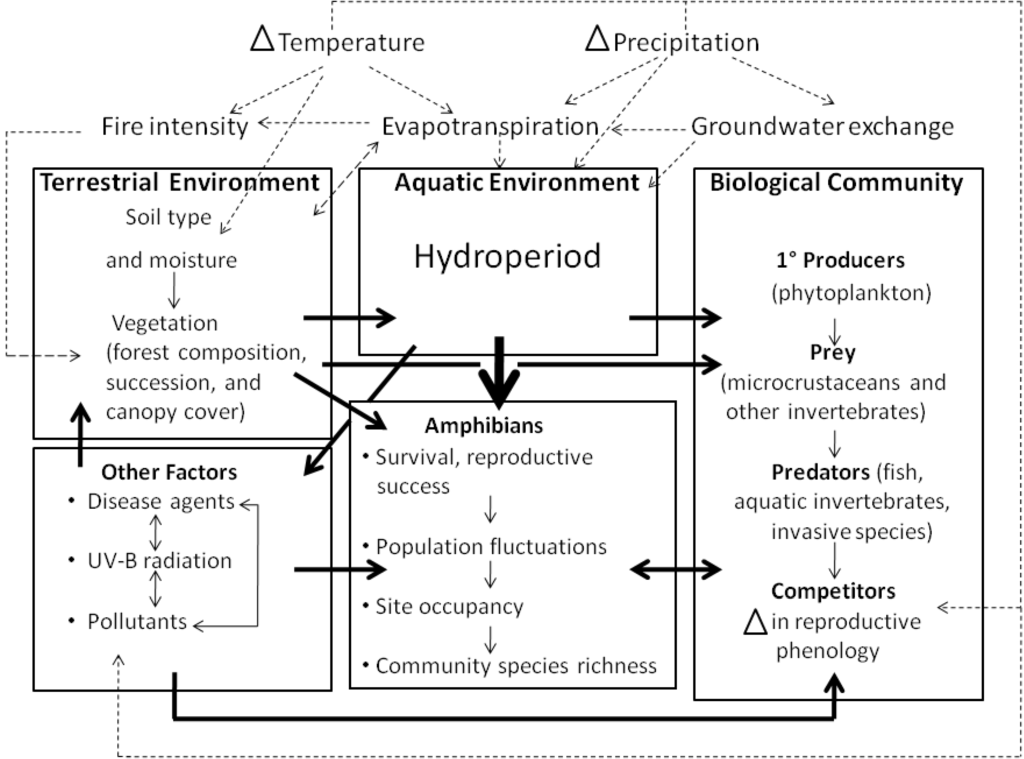
Figure 3.
Conceptual model of pathways through which changes in temperature and precipitation may impact amphibian populations, using a generalized freshwater ecosystem as an example. Changes in temperature and precipitation affect amphibians via the effects of climate on terrestrial and aquatic habitats, the biological community in which amphibians are embedded and other factors such as disease agents, UV-B radiation and pollution. Factors that influence the availability of water, such as the hydroperiod of aquatic habitats, are likely the most important constraints on the reproductive success and persistence of amphibians (indicated by heavier arrow). Dashed arrows indicate interactions among meteorological variables, and their effects on biological communities and the environments in which they occur. Heavy solid arrows indicate relationships among compartments; light solid arrows indicate relationships within compartments.
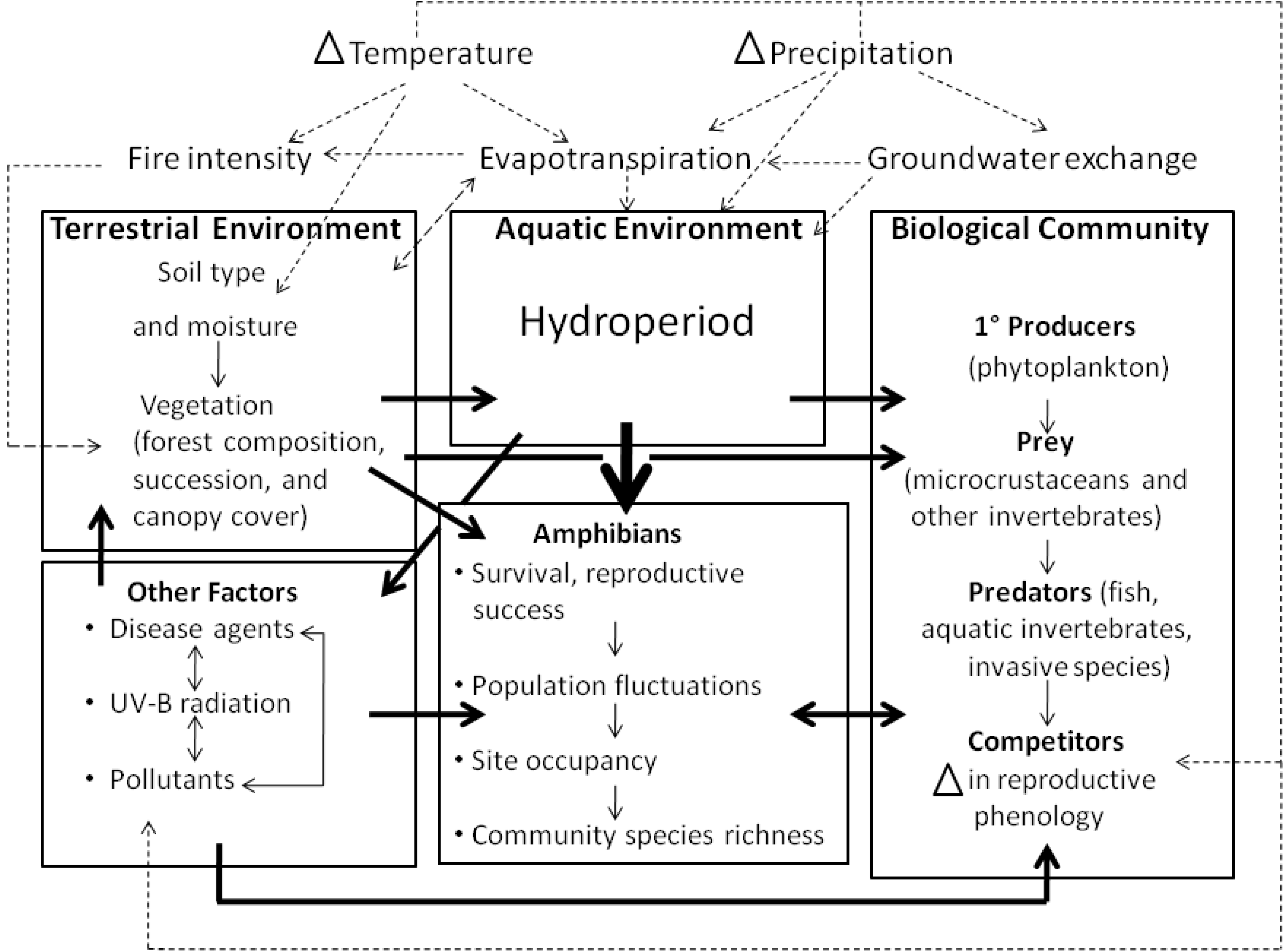
Figure 3.
Conceptual model of pathways through which changes in temperature and precipitation may impact amphibian populations, using a generalized freshwater ecosystem as an example. Changes in temperature and precipitation affect amphibians via the effects of climate on terrestrial and aquatic habitats, the biological community in which amphibians are embedded and other factors such as disease agents, UV-B radiation and pollution. Factors that influence the availability of water, such as the hydroperiod of aquatic habitats, are likely the most important constraints on the reproductive success and persistence of amphibians (indicated by heavier arrow). Dashed arrows indicate interactions among meteorological variables, and their effects on biological communities and the environments in which they occur. Heavy solid arrows indicate relationships among compartments; light solid arrows indicate relationships within compartments.
2.3.1. Habitat change
All amphibians, including species with nonaquatic eggs, depend to some extent on the availability of freshwater for successful reproduction [92]. For terrestrial-breeding species with direct development, the risk of evaporative water loss is likely the most important constraint on embryonic survival [93]. Because of their diverse reproductive modes that can involve metamorphosis from an aquatic larva to a terrestrial juvenile/adult for many species [66], amphibians are particularly vulnerable to degradation and change that affect both terrestrial and aquatic environments.
Amphibians that breed in ephemeral and often isolated bodies of water (e.g., vernal pools and intermittent headwater streams) are especially vulnerable to changes in temperature and precipitation [46]. Precipitation has increased by about 10% since 1910 across the contiguous United States [93] and is expected to continue to increase in many areas during the next century [93]. Yet, evapotranspiration losses from freshwater wetlands [95] and headwater streams [48,96] could possibly exceed precipitation during cyclical droughts, resulting in drying of aquatic sites (Figure 3). In turn, shortened wetland hydroperiods and disrupted stream flow will likely lead to an increased frequency of reproductive failure for many amphibians that breed in these habitats (Figure 3). Insufficient rainfall and shortened hydroperiods cause catastrophic reproductive failure in pond-breeding amphibians [97,98,99,100,101,102] and are likely contributors to population declines in several species ([31,54] but see [103]), some of which are federally threatened or endangered.
For coastal freshwater wetlands, global climate change is expected to affect the rate of rising sea level, along with the intensity and pattern of hurricanes and tropical storms [104]. Sea level rise contributes to the loss of coastal wetlands, and storm surge and its associated salt water intrusion during hurricanes can affect the abundance and species richness of amphibians in these habitats [105]. Moreover, storm surge and freshwater sheet flooding can introduce fishes (important amphibian predators) into normally isolated, fishless wetlands [106]. In the Coastal Plain of the southeastern U.S., these coastal freshwater wetlands are important breeding sites for a highly species-rich and abundant assemblage of amphibians [107].
Change that affects terrestrial habitats (e.g., changes in soil moisture and vegetation) can directly impact post-metamorphic adults and juvenile amphibians, as well as developing embryos of fully terrestrial species. Changes in temperature and precipitation can alter the frequency, intensity, duration and timing of fire, rates of evapotranspiration, and groundwater levels (Figure 3). Many terrestrial amphibians (e.g., salamanders of the family Plethodontidae) are desiccation-prone [108] and are well adapted to moist forest floors and cool, highly oxygenated streams that especially characterize montane regions. Warming may be a stressor for organisms that inhabit cool climates [109]; thus, these species may be particularly vulnerable to global warming and reductions in soil moisture.
The composition of the plant community in and around wetlands is a consequence of soil type and climate. Decreases in wetland hydroperiod and water depth during droughts can lead to an increase in grass, upland, and woody species, especially in shallower margins of wetlands, whereas prolonged hydroperiods promote the persistence of aquatic and emergent species [110]. The structure and composition of vegetation in the terrestrial environment can have an important influence on the successful reproduction and recruitment of amphibians in the aquatic habitat (Figure 3). Canopy openings and shifts in species composition of vegetation can alter microclimatic factors such as the duration and intensity of light, levels of temperature and moisture, as well as the accumulation, quality and decomposition of leaf litter [111]. In turn, the extent of canopy cover over wetlands [112,113,114] and the composition of leaf litter in the pond basin [115,116] may play a strong role in the reproductive success of some aquatic-breeding amphibians.
In fire-dependent ecosystems (e.g., habitats in the western and southeastern U.S.), variation in temperature and precipitation influence the frequency, intensity, duration and timing of fire. Fire suppression in pine savannas of the southeastern U.S. can lead to the establishment of hardwood and shrub species in otherwise historically pine-dominated (Pinus spp.) communities [117]. The presence of these invasive broad-leaved species in or near wetlands may increase the rate of evapotranspiration [118], thus potentially shortening wetland hydroperiod and compromising successful metamorphosis of larval amphibians.
2.3.2. Food availability
Tadpoles (larval anurans) are found in a wide array of freshwater microhabitats where they may function as “ecosystem engineers” by altering resources and influencing other consumers [119]. Tadpoles display a diversity of trophic modes, including herbivory (as grazers and scrapers of epilithon [often less accurately referred to as “periphyton”], as well as filter and suspension feeders), detritivory, carnivory (as cannibals, predators, and scavengers), and oophagy [119]. Global warming may influence the availability of autotrophic organisms that contribute to the diet of many tadpoles, as higher water temperatures are expected to increase rates of primary production and nutrient cycling [120]. Higher water temperatures can exacerbate many forms of water pollution and their associated blooms of filamentous cyanobacteria and green algae, bacteria, protozoans and small metazoans [121,122]. Such changes in the relative abundances of food items are known to influence tadpole feeding behavior and growth responses for some species [119].
Larval salamanders are gape-limited carnivores that prey on aquatic invertebrates [123]. Changes in temperature and precipitation, through their influence on wetland hydrology, can impact population densities, biomass and production of temporary wetland invertebrates ([124] Figure 3). In the northeast and other regions of the U.S., salamanders are the dominant vertebrate in intermittent, headwater forest streams [48]. In the UK, future projections suggest that spring macroinvertebrate abundance in headwater streams might decline by 21% for every 1 °C rise in water temperature, illustrating the sensitivity of headwater stream ecosystems to climate change [125]. Hydrologic variation is an important contributor to the biomass and assemblage structure of aquatic insects, benthic macroinvertebrates and macrozooplankton [126,127].
Because abundances and species richness of cladocerans and other wetland invertebrates increase seasonally [128] longer hydroperiods in a given year may translate into a larger density of microcrustaceans that enter dormancy in the soil when the pond dries. When the pond refills with winter rains in the subsequent season, calanoid, cyclopoid, and harpacticoid copepods, as well as cladocerans, hatch from resting eggs contained in the soil within hours [128], thus potentially producing a greater biomass of prey for larval salamanders. It is unclear, however, whether populations and communities of larval salamanders are limited by prey abundance ([123], but see references in [124]).
For terrestrial amphibians, higher amounts of precipitation increase humidity on the forest floor and underneath moisture-retaining cover (assuming this is not counteracted by increased temperatures), thus facilitating more opportunities to forage [129,130,131]. Above average rainfall in one year may increase the terrestrial prey base for adult females, which may then translate into greater reproductive success (through more “fit” offspring) in the subsequent year. For instance, Milanovich et al. [132] documented that, in the western slimy salamander (Plethodon albagula), the amount of precipitation one year prior to oviposition was correlated with average clutch size in the subsequent year. Food availability has been shown to affect traits related to individual fitness in other species of salamanders as well (Figure 3; [133,134]).
2.3.3. Community changes
On a regional scale, climate has a strong influence on the geographic distributions of species. For parapatric populations of competing species, climate-driven extensions and contractions in range boundaries can influence the dynamics of competitive interactions in contact zones [135]. Climate-induced variation in breeding phenology provides insight into the potential consequences that climate change may have on the structure of amphibian communities (Figure 3). For aquatic-breeding species, changes in the timing of reproduction can alter temporal overlap between competitors, predators and their prey [136,137,138,139,140].
Climate change is predicted to have a significant effect on the persistence and spread of non-indigenous species which, in turn, could have profound consequences for the structure and stability of natural ecosystems [141]. As alien predators, various introduced species are known to be contributing to amphibian population declines around the world [142]. Examples of introduced species of amphibians that are of particular concern include the Cuban treefrog (Osteopilus septentrionalis), the American bullfrog (Lithobates catesbeianus), and the cane toad (Rhinella marina), all of which prey upon, and compete with, native amphibians. Projections of various climate change scenarios indicate the likelihood of expansion of the current distribution of Cuban treefrogs in North America [143], cane toads in Australia [144], and American bullfrogs in various parts of the world [145].
Climate-induced changes that influence the occurrence of keystone species within communities will potentially affect the other members of the community as well. Important examples of amphibian species that function as keystone predators include the mole salamander (Ambystoma talpoideum) and the eastern newt (Notophthalmus viridescens) [146,147,148]. Interestingly, the presence of the predatory eastern newt can reduce or eliminate the negative, competitive impact that introduced Cuban treefrog tadpoles have on tadpoles of native species [149]. Although mole salamanders and eastern newts are not yet considered species of conservation concern, they have declined significantly over 28 yr in some areas of the Coastal Plain of the southeastern U.S. [107]. Both of these species require intermediate to long hydroperiod wetlands for successful reproduction. If future climate change shortens the hydrology of breeding sites used by these species, there may be cascading impacts on the structure of amphibian communities in some freshwater wetlands.
2.4. Interactions with Other Stressors
2.4.1. Emerging diseases
Overall, the effects of climate change on amphibian diseases are hard to predict, because many mechanisms are involved. While some pathogens may increase in prevalence and severity, others may decline. Understanding the processes and trends of climate change and disease will be essential in preventing epidemics in wildlife and humans.
Climate change has the potential to alter species interactions in many ways. Of particular concern are the effects of changing climate on host-pathogen interactions which could dramatically alter disease dynamics. While some host-pathogen systems may experience a decrease in disease severity, it is predicted that most will observe an increase in epidemics [150].
2.4.2. Immunity
Intimately tied with disease is immune function and certain climate changes may affect immune systems. For example, temperature may have profound direct effects on amphibian immune function [151]. Ectotherms may use thermoregulatory behavior to modulate immune function and reduce the risk of infection [152]. Several key aspects of immunity are suppressed at low temperatures (<20 °C) in ectotherms. For example, serum complement level [153,154], activity of anti-microbial peptides [155], T and B-lymphocyte numbers and proliferation activity [154,156,157,158], esosinophil numbers [154], and antibody production [157,159,160] are reduced at low temperatures in ectotherms. However some aspects of immune activity appear to be robust to low temperature. Phagocytic activity and respiratory burst [161,162] as well as numbers of circulating neutrophils [154,163], and activity of nonspecific cytotoxic cells [160] may be relatively stable or even enhanced at low temperatures in some species. Maintenance of non-specific innate responses at low temperatures may function to offset suppression of other immune parameters, and may be especially important in a seasonal context. Few studies have addressed how various immune components are affected by elevated temperature. Increases in ambient temperature or water temperature potentially may have implications for both post-metamorphic and larval amphibians. For example, in some fish species, at high temperatures (>30 °C) there is suppression of phagocytic activity and serum killing ability [164].
Recovery of some aspects of immune responsiveness may be rapid when temperatures are raised. For example, serum complement levels may increase rapidly (as quickly as 48h) after animals are restored to warmer temperatures after being held at low temperatures [153,154]. It is unclear whether all aspects of immunity recuperate in this manner, but it is possible that cellular processes (e.g., maturation and proliferation of T and B lymphocytes) require slightly longer time to rebound from abrupt changes in temperature [158]. Therefore, to some extent, a lag exists between changes in temperature, and subsequent changes in immune parameters [163].
Clearly, there is an urgent need for studies that examine the potential for immunological changes occurring over more incremental temperatures shifts of several degree. However, the robust body of literature available on temperature-dependent immunity in ectotherms provides a springboard for initial hypothesis testing based on climate change models. In addition to directional changes in temperature predicted with climate change, it may be important to consider how variability in temperature regimes affects immunocompetence in amphibians. If amphibians modulate immune responses based on more subtle temperature differences then the energetic cost of constantly upregulating and downregulating immunological parameters based on minor but frequent temperature fluctuations may be substantial. It is also unclear if lag time between temperature-dependent aspects of immunity may serve as more vulnerable periods of time for amphibians [154]. Finally, it is unclear how temperature-dependent changes in immunity may be affected when individuals are simultaneously dealing with other physiological demands and environmental stressors.
Depending upon the model, a specific region may have more or less precipitation in the future than present and may be colder or warmer than present [17,44]. Such changes could significantly affect hydroperiods critical to amphibian life cycles. Thus, Gervasi and Foufopoulos [86] showed that wood frog (Rana sylvatica) tadpoles exposed to desiccation had shorter development times, weaker cellular immune system responses and lower total leukocyte numbers than animals from control groups. Measures of immune response showed a decrease with increasing severity of the desiccation treatment. It was unclear whether the observed depression in immune response was transient or permanent. However, even temporary periods of immune system suppression shortly after metamorphosis could lead to greater susceptibility to pathogens.
2.4.3. Emerging amphibian diseases
Amphibians are susceptible to many types of pathogens including trematodes, copepods, fungi, oomycetes, bacteria and viruses. In amphibians, infectious diseases have been implicated in numerous population declines [7,165,166,167]. Climate change has the potential to alter the dynamics of these pathogens in both direct and indirect ways to either the benefit or detriment of the amphibian host.
Climate change could alter host-pathogen interactions through changes in temperature. As global temperatures increase, most pathogens will experience faster growth and reproduction which could potentially increase the severity of infectious diseases [31,150]. Additionally, warmer winters and nighttime temperatures may reduce the cycle of pathogen die-offs that naturally occur during these colder times [168,169]. As stated above, higher water temperatures can induce eutrophication with blooms of algae, bacteria, protozoans and small metazoans [121,122]. The trematode parasite Ribeiroia ondatrae sequentially infects birds, snails, and amphibian larvae, frequently causing severe limb deformities and mortality [170]. Johnson et al. [171] showed how eutrophication promoted amphibian disease by increasing the density of infected snail hosts and by enhancing per-snail production of infectious parasites.
Climate change could also alter the host-pathogen relationship through changes in precipitation or hydrology. For example, many amphibian pathogens are aquatically transmitted (e.g., oomycetes, trematodes and certain fungi) so increased rainfall that leads to more standing water could increase rates of transmission. Also, aquatically-transmitted pathogens can be sensitive to water conditions. For example, the pathogenic “chytrid” fungus Batrachochytrium dendrobatidis (BD) dies after 3 hours desiccation [172], so loss of moisture or precipitation could affect the survival of this pathogen. Kriger [173] argues that droughts should reduce the severity of BD epidemics, but there is also some evidence that droughts actually increase outbreaks [53,174]. High temperatures impede growth and can kill BD in the laboratory [175,176].
Changes in climate could also shift the ranges the pathogen, the hosts, or the pathogen vector. As climate change alters local habitats, new areas may appear that are suitable for the host or pathogen while others disappear. Seimon et al. [177] documented upward range expansion of both amphibian hosts and BD in the Andes. As high elevation sites experience loss of glaciers, this opens up new habitat for anurans in the area. BD has been detected on amphibians at these new sites, demonstrating a shift in both host and pathogen ranges. In other scenarios, the pathogen or host could potentially shift ranges without the other following.
On a local scale, an experimental field study in Oregon by Kiesecker et al. [20] illustrates a complex interrelationship among climate change, UV-B radiation and amphibian population declines and parallels a tropical study by Pounds et al. [178]. Kiesecker et al. [20] linked ENSO events with decreased winter precipitation in the Oregon USA Cascade Range. They suggested that less winter snow pack resulted in lower water levels when western toads (Bufo boreas) breed in early spring. Toad embryos developing in shallower water are exposed to higher levels of UV-B radiation, which results in increased mortality from the pathogenic oomycete, Saprolegnia ferax.
Another example of changing hydrology affecting amphibian diseases on a local level is that of Kupferberg et al. [179]. The authors found that outbreaks of a pathogenic copepod, Lernaea cyprinacea, are more severe following unusually warm summers in northern California. The authors speculate that this could be partly caused by changes in hydrology. During the warm summer of the outbreak, water levels were reduced, forcing amphibian larvae into high densities. Reduced discharge slowed river velocities which could have allowed for easier transmission of the copepod parasite.
Pounds et al. [178] identified two dynamics that are global in nature: climate change and the potential spread of a highly virulent pathogen (discussed in Blaustein and Dobson [180]). BD is implicated as the proximate cause for Atelopus population crashes and species extinctions in tropical America. Pounds et al. [178] presented a mechanistic explanation for how climate change may influence outbreaks of BD by modifying conditions in montane areas of Central and South America where nighttime temperatures are shifting closer to the thermal optimum for BD, while increased daytime cloudiness prevents frogs from finding thermal refuges from the pathogen. Climate change and outbreaks of chytridiomycosis have been reported in several other studies. Bosch et al. [181] showed a significant association between rising temperatures and outbreaks of chytridiomycosis in Spain. D’Amen and Bombi [182] showed the emergence of BD after a climatic shift or extreme weather event in Italy. Increases in chydriomycosis were correlated with low summer temperatures in Australia [183].
Lips et al. [184] tested the robustness of the climate-linked epidemic hypothesis by reanalyzing data on declines and extinctions of Atelopus species in the same region where Pounds et al. [178] performed their research. The Lips et al. [185] analysis did not support the climate-linked epidemic hypothesis. They suggest that BD is an introduced pathogen that has been spreading throughout the American tropics since the 1970s. This spatiotemporal hypothesis suggests that BD spreads independent of climate shifts.
The assertions of Pounds et al. [178] and Lips et al. [184] have been questioned in several recent papers. Lampo et al. [185] demonstrated that some of the assumptions of the spatiotemporal hypothesis are inconsistent with data on extinctions and declines and population crashes may not be due to the arrival of BD. Parmesan and Singer [186] questioned the statistical analysis of Lips et al. [184] and suggest that both hypotheses are supported by numerous studies, are not mutually exclusive and may be interactive. Rohr et al. [187] examined the climate-linked epidemic hypothesis and the spatiotemporal spread hypothesis. Rohr et al. [187] found no support for the “chytrid-thermal optimum hypothesis” [178,180]. However, they suggest that climate change is likely to play an important role in amphibian population declines worldwide.
Alford et al. [188] stated that during the decline of amphibians in New South Wales, Australia, very few moribund specimens examined were positive for BD. Alford et al. [188] suggested that stress during growth and development may contribute to limb asymmetries in amphibians. They suggested that unusual climate conditions may be a significant stressor on amphibian growth and development. In Australia, they attributed the greater frequency of limb asymmetries in “pre-declining populations” compared with non-declining “control” populations to stress from “dramatic regional warming”. They suggest that BD may not be the only factor involved in amphibian population declines in this region and that climate conditions alone may be an important factor in declines.
Di Rosa et al. [189] provide evidence that BD may be present without causing chytridiomycosis in frogs in Italy and that other pathogens may also contribute to frog declines in this region. In agreement with Pounds et al. [190] they suggest that climate conditions may make amphibians more susceptible to a number of pathogens.
Evidence from long-term studies at the Savannah River site in South Carolina, USA, illustrates that populations of several amphibian species have been in decline [54]. At this site, the presence of BD was rare and there was no evidence of chytridiomycosis. The investigators concluded that the population declines in this region were more likely due to extreme weather such as low rainfall and shortened hydroperiod for breeding rather than a BD epidemic.
2.4.4. UV-B radiation
Changes in precipitation which affect hydroperiods and habitat alteration such as deforestation, can expose amphibians to levels of solar radiation that they have not previously experienced.
A number of investigators at various sites around the world have shown via experiments that exposure to ambient levels of UV-B radiation decreases the hatching success of many (but not all) amphibian species at natural oviposition sites in the field [35,191,192]. These studies have demonstrated that the embryos of some species are less resistant to UV-B radiation than others [191,192].
Importantly, even though hatching rates of some species may appear unaffected by ambient UV-B radiation in field experiments, an increasing number of studies illustrate a variety of sublethal effects due to UV exposure. Depending on the species, these effects have been observed in all life stages [191]. For example, when exposed to UV-B radiation, amphibians may change their behavior [193,194,195], growth and development may be slowed [196,197,198], or a number of developmental and physiological malformations may occur [199,200,201,202,203,204]. Sublethal effects may become evident even in species whose embryos appeared to be resistant in field experiments. Moreover, numerous field and laboratory experiments have also shown that UV-B radiation interacts synergistically with a variety of chemicals, low pH and pathogens [191,205,206,207].
The experimental field study in Oregon USA by Kiesecker et al. [20] discussed above illustrates the interrelationships among precipitation, climate change and UV radiation and how they may interact to harm amphibians and supports the tropical study by Pounds et al. [9]. Merilä et al. [208] present an interesting scenario combining climate change, UV radiation and amphibian breeding. They suggest that if amphibians are breeding earlier in northern ecosystems, then their annual life cycle will not only start earlier relative to the calendar date but also with regard to maximum UV-B exposure. UV-B exposure would be lower than if amphibians bred later in the spring. Thus, they suggest that global warming, which may induce amphibians to breed earlier may counteract the effects of increasing UV-B levels generated by a thinning ozone layer.
Temperature and UV-B radiation may also interact synergistically. Two recent studies [209,210] demonstrate reduced survival of amphibian larvae when cold temperatures and UV-B radiation are combined. There are several possible mechanisms by which this could occur. For example, DNA repair enzymes, such as photolyase, are essential in mitigating the damage to DNA caused by exposure to UV-B radiation. Cold temperatures can reduce the activity of photolyase and slow repair of damaged DNA. Additionally, cold temperatures could indirectly increase the threat of UV-B by altering the behavior of larval amphibians such that they are exposed to higher levels of UV-B. Any amphibian inhabiting cold habitats with high levels of UV-B could be particularly at risk. Global warming is likely to change temperatures in many habitats, which will alter the threat of UV-B for amphibians and make the risk of negative effects from UV-B exposure difficult to predict.
2.4.5. Air-borne contaminants
Besides global environmental changes, numerous agents, including air-borne pollutants, may be toxic to amphibians by themselves or interact with warming trends and UV radiation in a synergistic fashion. As worldwide agricultural demands rise there will be significant increases in pesticide use [211]. Pesticides have the potential for atmospheric transport [212,213] and deposition where they may be available for uptake by biota, especially by amphibians through their permeable skin, and may alter nutrient dynamics or increase water clarity allowing for greater penetration of ultraviolet radiation [214]. Contaminants transported atmospherically are potentially harmful to amphibians [215,216,217,218] and they may interact with UV-B radiation, other contaminants and changes in climate [190]. For example, acid deposition from the atmosphere to aquatic systems has been linked to both lethal and sublethal effects on developing amphibians, particularly those that breed in temporary ponds [218]. However, the effects of other atmospheric contaminants on amphibians have not been well documented.
Pounds and Crump [220] suggested that atmospheric scavenging of contaminants by clouds might concentrate them and release them in remote areas such as Monteverde, Costa Rica where many populations of amphibians have declined. This effect may be particularly important under unusually hot, dry conditions [9,220]. In California, atmospheric deposition of organophosphate pesticides from the highly agricultural Central Valley may be contributing to declines in frog populations [221]. Pesticides may adhere to foliage where they may threaten native species [221]. Activity of the enzyme cholinesterase in treefrogs (Hyla regilla) was impaired in areas where populations of ranid frogs were declining [221]. Cholinesterase impairment might be linked to the presence of organophosphate pesticides [221]. Using GIS analysis, Davidson et al. [212] concluded that pesticides carried upwind from the Central Valley of California may be contributing to declines in red-legged (Rana aurora) frogs in California.
3. Conclusions
Our review suggests that climate change may have both direct and indirect effects on amphibians at the individual, population and community level. Shifts in amphibian ranges are predicted under certain climate model scenarios. Changes in climate may affect survival, growth, reproduction and dispersal capabilities. Moreover, climate change can alter amphibian habitats including vegetation, soil, and hydrology. Climate change can influence food availability and overall predator-prey and competitive interactions which can alter community structure. Climate change can also alter pathogen-host dynamics and greatly influence how diseases are manifested. Changes in climate can interact with other stressors such as UV-B radiation and contaminants. The interactions between all these factors are complex and are probably driving some amphibian population declines and extinctions.
We suggest that a multidisplinary approach is essential for understanding the direct and indirect effects of climate change on amphibians. Climate modeling should be fortified with both laboratory and field experiments. Experimental studies should be bolstered with molecular work, especially using molecular tools that can provide clues as to how amphibians are affected at the physiological level [222]. Molecular biologists, ecologists and modelers working together may be able to help us understand the broad impacts of climate change on amphibians.
Acknowledgements
We thank Ezy Yoo, S. Turner, J. Lerner, L. Biederman and P. Beck for their help.
References
- Lawton, J.H.; May, R.M. Extinction Rates; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Wilson, E.O. The Diversity of Life; Harvard University Press: Cambridge, MA, USA, 1992. [Google Scholar]
- Houlahan, J.E.; Findlay, C.S.; Schmidt, B.R.; Meyer, A.H.; Kuzmin, S.L. Quantitative evidence for global amphibian population declines. Nature 2000, 404, 752–755. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischmann, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef]
- Lannoo, M. Amphibian Declines: The Conservation Status of United States Species; University California Press: Berkeley, CA, USA, 2005. [Google Scholar]
- Mendelson, J.R., III; Lips, K.R.; Gagliardo, R.W.; Rabb, G.B.; Collins, J.P.; Diffendorfer, J.E.; Daszack, P.; Ibáñez, R.; Zippel, K.C.; Lawson, D.P.; Wright, K.M.; Stuart, S.N.; Gascon, C.; Silva, H.R.; Burrowes, P.A.; Joglar, R.L.; La Marca, E.; Lötters, S.; Perez, L.H.; Weldon, C.; Hyatt, A.; Rodriguez-Mahecha, J.V.; Hunt, S.; Robertson, H.; Lock, B.; Raxworthy, C.J.; Frost, D.R.; Lacy, R.C.; Alfrod, R.A.; Campbell, J.A.; Parra-Olea, G.; Bolaños, F.; Domingo, J.J.C.; Halliday, T.; Murphy, J.B.; Wake, M.H.; Coloma, L.A.; Kuzmin, S.L.; Price, M.S.; Howell, K.M.; Lau, M.; Pethiyagoda, R.; Boone, M.; Lannoo, M.J.; Blaustein, A.R.; Dobson, A.; Griffiths, A.; Crump, M.L.; Wake, D.B.; Brodie, E.D.J. Confronting amphibian declines and extinctions. Science 2006, 313, 48. [Google Scholar] [CrossRef]
- Bielby, J.; Cooper, N.; Cunningham, A.A.; Garner, T.W.J.; Purvis, A. Predicting susceptibility to rapid declines in the world’s frogs. Conserv. Lett. 2008, 1, 82–90. [Google Scholar] [CrossRef]
- Pounds, J.A.; Fogden, M.P.L.; Savage, J.M.; Gorman, G.C. Test of null models for amphibian declines on a tropical mountain. Conserv. Biol. 1997, 11, 1307–1322. [Google Scholar] [CrossRef]
- Pounds, J.A.; Fogden, M.P.L.; Campbell, J.H. Biological responses to climate change on a tropical mountain. Nature 1999, 398, 611–615. [Google Scholar] [CrossRef]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A review from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Kiesecker, J.M. Complexity in conservation: Lessons from the global decline of amphibian populations. Ecol. Lett. 2002, 5, 597–608. [Google Scholar] [CrossRef]
- van der Leun, J.C.; Tang, X.; Tevini, M. Environmental Effects of Ozone Depletion 1998 Assessment; Elsevier: Lausanne, Switzerland, 1998. [Google Scholar]
- Peters, R.L; Lovejoy, T.E. Global Warming and Biological Diversity; Yale University Press: New Haven, CT, USA, 1992. [Google Scholar]
- Reaser, J.K.; Blaustein, A.R. Repercussions of global change. In Status and Conservation of North American Amphibians; Lannoo, M., Ed.; University of California Press: Berkeley, CA, USA, 2005; pp. 60–63. [Google Scholar]
- Cockell, C.S.; Blaustein, A.R. Ecosystems, Evolution and Ultraviolet Radiation; Springer: New York, NY, USA, 2001. [Google Scholar]
- IPCC. Climate change 2007: the physical science basis. In Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Lawler, J.J.; Shafer, S.L.; White, D.; Kareiva, P.; Maurer, E.P.; Blaustein, A.R.; Bartlein, P.J. Projected climate-induced faunal change in the western hemisphere. Ecology 2009, 90, 588–597. [Google Scholar] [CrossRef]
- Andrady, A.; Aucamp, P.J.; Bais, A.; Ballaré, C.L.; Björn, L.O.; Bornman, J.F.; Caldwell, M.; Cullen, A.P.; Erickson, D.J.; de Gruijl, F.R.; Häder, D.P.; Ilyas, M.; Kulandaivelu, G.; Kumar, H.D.; Longstreth, J.; McKenzie, R.L.; Norval, M.; Paul, N.; Redhwi, H.H.; Smith, R.C.; Solomon, K.R.; Sulzberger, B.; Takizawa, Y.; Tang, X.; Teramura, A.H.; Torikai, A.; van der Leun, J.C.; Wilson, S.R.; Worrest, R.C.; Zepp, R.G. Environmental effects of ozone depletion and its interactions with climate change: progress report, 2008. Photochem. Photobiol Sci. 2009, 8, 13–22. [Google Scholar] [CrossRef]
- Ovaska, K. Vulnerability of amphibians in Canada to global warming and increased ultraviolet radiation. Amphibians in decline: Canadian studies of a global problem, society for the study of amphibians and reptiles. Herpetol. Conserv. 1997, 1, 206–225. [Google Scholar]
- Kiesecker, J.M.; Blaustein, A.R.; Belden, L.K. Complex causes of amphibian population declines. Nature 2001, 410, 681–684. [Google Scholar] [CrossRef]
- Tevini, M. UV-B Radiation and Ozone Depletion: Effects on Humans, Animals, Plants, Microorganisms, and Materials; Lewis Publishers: Boca Raton, FL, USA, 1993. [Google Scholar]
- Schindler, D.W.; Curtis, P.J.; Parker, B.R.; Stainton, M.P. Consequences of climate warming and lake acidification for UV-B penetration in North American boreal lakes. Nature 1996, 379, 705–708. [Google Scholar] [CrossRef]
- Kerr, J.B.; McElroy, C.T. Evidence for large upward trends of ultraviolet-B radiation linked to ozone depletion. Science 1993, 262, 1032–1034. [Google Scholar]
- Herman, J.R.; Bhartia, P.K.; Ziemke, J.; Ahmad, Z.; and Larko, D. UV-B increases (1979-1992) from decreases in total ozone. Geophys. Res. Lett. 1996, 23, 2117–2120. [Google Scholar] [CrossRef]
- Middleton, E.M.; Herman, J.R.; Celarier, E.A.; Wilkinson, J.W.; Carey, C.; Rusin, R.J. Evaluating ultraviolet radiation exposure with satellite data at sites of amphibian declines in Central and South America. Conserv. Biol. 2001, 15, 914–929. [Google Scholar] [CrossRef]
- Donnelly, M.A.; Crump, M.L. Potential effects of climate change on two Neotropical amphibian assemblages. Clim. Change 1998, 39, 541–561. [Google Scholar] [CrossRef]
- Alford, R.A.; Richards, S.J. Global amphibian declines: a problem in applied ecology. Ann. Rev. Ecol. Syst. 1999, 30, 133–165. [Google Scholar] [CrossRef]
- Alexander, M.A.; Eischeid, J.K. Climate variability in regions of amphibian declines. Conserv. Biol. 2001, 15, 930–942. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Hatch, A.C.; Belden, L.K.; Scheessele, E.; Kiesecker, J.M. Global change: challenges facing amphibians. In Amphibian Conservation; Semlitsch, R.D., Ed.; Smithsonian Press: Washington, DC, USA, 2003; pp. 187–198. [Google Scholar]
- Carey, C.; Alexander, M.A. Climate change and amphibian declines: Is there a link? Divers. Distribut. 2003, 9, 111–121. [Google Scholar] [CrossRef]
- McMenamin, S.K.; Hadley, E.A.; Wright, C.K. Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proc. Natl. Acad. Sci. USA 2008, 105, 16988–16993. [Google Scholar] [CrossRef]
- Heyer, W.R.; Rand, A.S.; Goncalvez da Cruz, C.A.; Peixoto, O.L. Decimations, extinctions and colonizations of frog populations in southeast Brazil and their evolutionary implications. Biotropica 1988, 20, 230–235. [Google Scholar] [CrossRef]
- Weygoldt, P. Changes in the composition of mountain stream frog communities in the Atlantic Mountains of Brazil: Frogs as indicators of environmental deteriorations? Neotrop. Fauna & Environ. 1989, 24, 249–255. [Google Scholar]
- Crump, M.L.; Hensley, F.R.; Clark, K.L. Apparent decline of the golden toad: Underground or extinct? Copeia 1992, 1, 413–420. [Google Scholar]
- Blaustein, A.R.; Belden, L.K.; Olson, D.H.; Green, D.L.; Root, T.L.; Kiesecker, K.M. Amphibian breeding and climate change. Conserv. Biol. 2001, 15, 1804–1809. [Google Scholar] [CrossRef]
- Bustamante, M.R.; Ron, S.R.; Coloma, L.A. Cambios en la diversidad en siete communidades de anuros en los Andes de Ecuador. Biotropica 2005, 37, 180–189. [Google Scholar] [CrossRef]
- Raxworthy, C.J.; Pearson, R.G.; Rabibisoa, N.; Rakotondrazafy, A.M.; Ramanamanjato, J.-B.; Raselimanana, A.P.; Wu, S.; Nussbaum, R.A.; Stone, D.A. Extinction vulnerability of tropical montane endemism from warming and upslope displacement: a preliminary appraisal for the highest massif in Madagascar. Glob. Change Biol. 2008, 14, 1703–1720. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Ann. Rev. of Ecol. and Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar]
- Lovejoy, T.E.; Hannah, L. Climate Change and biodiversity; Yale University Press: New Haven, CT, USA, 2005. [Google Scholar]
- Araújo, M. B.; Thuiller, W.; Pearson, R.G. Climate warming and the decline of amphibians and reptiles in Europe. J. Biogeo. 2006, 33, 1712–1728. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: are climate envelope models useful? Glob. Ecol. Biogeo. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Lawler, J.J.; White, D.; Neilson, R.P.; Blaustein, A.R. Predicting climate-induced range shifts: model differences and model reliability. Glob. Change Biol. 2006, 12, 1568–1584. [Google Scholar] [CrossRef]
- Lawler, J.J.; Shafer, S.L.; Bancroft, B.A.; Blaustein, A.R. Projected climate impacts for the amphibians of the western hemisphere. Conserv. Biol. 2010, 24, 38–50. [Google Scholar] [CrossRef]
- Rome, L.C.; Stevens, E.D.; John-Alder, H.B. Temperature and thermal acclimation and physiological function. In Environmental Physiology of the Amphibia; Feder, M.E., Burggren, W.W., Eds.; University of Chicago Press: London, UK, 1992; pp. 183–205. [Google Scholar]
- Hillyard, S.D. Behavioral, molecular and integrative mechanisms of amphibian osmoregulation. J. Exper. Zool. 1999, 283, 662–674. [Google Scholar] [CrossRef]
- Brooks, R.T. Potential impacts of global climate change on the hydrology and ecology of ephemeral freshwater systems of the forests of the northeastern United States. Clim. Change 2009, 95, 469–483. [Google Scholar] [CrossRef]
- Rios-López, N. Effects of increased salinity on tadpoles of two anurans from a Caribbean coastal wetland in relation to their natural abundance. Amphibia-Reptilia 2008, 29, 7–18. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W.; Mata, L.J.; Arnell, N.W.; Döll, P.; Kabat, P.; Jiménez, B.; Miller, K.A. Freshwater Resources and Their Management. Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth. In Assessment Report of the Intergovernmental Panel on Climate Change; Oki, T., Sen, Z., Shiklomanov, I.A., Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- IPCC. Assessment of Observed Changes and Responses in Natural and Managed Systems. In Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Rosenzweig, C., Casassa, G., Karoly, D.J., Imeson, A., Liu, C., Menzel, A., Rawlins, S., Root, T.L., Seguin, B., Tryjanowski, P., Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Reading, C.J. Linking global warming to amphibian declines through its effect on female body condition and survivorship. Oecologia 2007, 151, 125–131. [Google Scholar] [CrossRef]
- Burrowes, P.A.; Joglar, R.L.; Green, D.L. Potential causes for amphibian declines in Puerto Rico. Herpetologica 2004, 60, 141–154. [Google Scholar] [CrossRef]
- Daszak, P.; Scott, D.E.; Kilpatrick, A.M.; Faggioni, C.; Gibbons, J.W.; Porter, D. Amphibian population declines at Savannah River Site are linked to climate, not chytridiomycosis. Ecology 2005, 86, 3232–3237. [Google Scholar] [CrossRef]
- Carey, C.; Heyer, W.R.; Wilkinson, J.; Alford, R.A.; Arntzen, J.W.; Halliday, T.; Hungerford, L.; Lips, K.R.; Middelton, E.M.; Orchard, S.A.; Rand, A.S. Amphibian declines and environmental change: Use of remote-sensing data to identify environmental correlates. Conser. Biol. 2001, 15, 903–913. [Google Scholar] [CrossRef]
- Beebee, T.J.C. Amphibian breeding and climate. Nature 1995, 374, 219–220. [Google Scholar] [CrossRef]
- Kusano, T.; Inoue, M. Long-term trends toward earlier breeding of Japanese amphibians. J. Herp. 2008, 42, 608–614. [Google Scholar] [CrossRef]
- Chadwick, E.A.; Slater, F.M.; Ormerod, S.J. Inter- and intraspecific differences in climatically mediated phenological change in coexisting Triturus species. Glob. Change Biol. 2006, 12, 1069–1078. [Google Scholar] [CrossRef]
- Gibbs, J.P.; Breisch, A.R. Climate warming and calling phenology of frogs near Ithaca, New York, 1900−1999. Conserv. Biol. 2001, 15, 1175–1178. [Google Scholar] [CrossRef]
- Hartel, T. Weather conditions, breeding date and population fluctuation in Rana dalmantina from central Romania. Herpetol. J. 2008, 18, 1–5. [Google Scholar] [CrossRef]
- Vaira, M. Annual variation of breeding patterns of the toad, Melanophryniscus rubriventris (Vellard, 1947). Amphibia-Reptilia 2005, 26, 193–199. [Google Scholar] [CrossRef]
- Richter-Boix, A.; Llorente, G.A.; Montori, A. Breeding phenology of an amphibian community in a Mediterranean area. Amphibia-Reptilia 2006, 27, 549–559. [Google Scholar] [CrossRef]
- Mills, N.E.; Barnhart, M.C. Effects of hypoxia on embryonic development in two Ambystoma and two Rana species. Physiol. Biochem. Zool. 1999, 72, 179–188. [Google Scholar] [CrossRef]
- Wassersug, R.J.; Seibert, E.A. Behavioral responses of amphibian larvae to variation in dissolved oxygen. Copeia 1975, 1975, 86–103. [Google Scholar] [CrossRef]
- Eggert, C. Sex determination: the amphibian models. Reprod. Nutr. Dev. 2004, 44, 539–549. [Google Scholar] [CrossRef]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; Johns Hopkins University Press: Baltimore, MD, USA, 1986. [Google Scholar]
- Volpe, E.P. Embryonic temperature tolerance and rate of development of Bufo valliceps. Physiol. Zool. 1957, 30, 164–175. [Google Scholar]
- Vonesh, J.R.; De la Cruz, O. Complex life cycles and density dependence: assessing the contribution of egg mortality to amphibian declines. Oecologia 2002, 133, 325–333. [Google Scholar] [CrossRef]
- Biek, R.; Funk, W.C.; Maxell, B.A.; Mills, L.S. What is missing in amphibian decline research: Insights from ecological sensitivity analysis. Conserv. Biol. 2002, 16, 728–734. [Google Scholar] [CrossRef]
- Broomhall, S.D. Egg temperature modifies predator avoidance and the effects of the insecticide endosulfan on tadpoles of an Australian frog. J. Appl. Ecol. 2004, 41, 105–113. [Google Scholar] [CrossRef]
- Govindarajulu, P.P.; Anholt, B.R. Interaction between biotic and abiotic factors determines tadpole survival rate under natural conditions. Ecoscience 2006, 13, 413–421. [Google Scholar] [CrossRef]
- Álvarez, D.; Nicieza, A.G. Effects of temperature and food quality on anuran larval growth and metamorphosis. Funct. Ecol. 2002, 16, 640–648. [Google Scholar] [CrossRef]
- Loman, J. Temperature genetic and hydroperiod effects on metamorphosis of brown frogs Rana arvalis and R. temporaria in the field. J. Zool. 2002, 258, 115–129. [Google Scholar] [CrossRef]
- Morand, A.; Joly, P.; Grolet, O. Phenotypic variation in metamorphosis in five anuran species along a gradient of stream influence. C.R. Acad. Sci. Paris Life Sciences 1997, 320, 645–652. [Google Scholar]
- Buchholz, D.R.; Hayes, T.B. Larval period comparison for the spadefoot toads Scaphiopus couchii and Spea multiplicata (Pelobatidae: Anura). Herpetologica 2000, 56, 455–468. [Google Scholar]
- Voss, S.R. Relationship between stream order and length of larval period in the salamander Eurycea wilderae. Copeia 1993, 1993, 736–742. [Google Scholar] [CrossRef]
- Browne, R.K.; Edwards, D.L. The effect of temperature on the growth and development of the endangered green and golden bell frog (Litoria aurea). J. Thermal Biol. 2003, 28, 295–299. [Google Scholar] [CrossRef]
- Berven, K.A.; Gill, D.E.; Smith-Gill, S.J. Countergradient selection in the Green Frog, Rana clamitans. Evolution 1979, 33, 609–623. [Google Scholar] [CrossRef]
- Beachy, C.K. Effects of larval growth history on metamorphosis in a stream-dwelling salamander (Desmognathus ochrophaeus). J. Herpetol. 1995, 29, 375–382. [Google Scholar] [CrossRef]
- Hickerson, C.M.; Barker, E.L.; Beachy, C.K. Determinants of metamorphic timing in the black-bellied salamander, Desmognathus quadramaculatus. Southeast. Nature. 2005, 4, 33–50. [Google Scholar] [CrossRef]
- Wilbur, H.M.; Collins, J.P. Ecological aspects of amphibian metamorphosis. Science 1973, 182, 1305–1314. [Google Scholar]
- Werner, E.E. Amphibian metamorphosis: growth rate, predation risk, and the optimal size at transformation. Amer. Nat. 1986, 128, 319–341. [Google Scholar]
- John-Alder, H.B.; Morin, P.J. Effects of larval density on jumping ability and stamina in newly metamorphosed Bufo woodhousii fowleri. Copeia 1990, 1, 856–860. [Google Scholar] [CrossRef]
- Goater, C.P.; Semlitsch, R.D.; Bernasconi, M.V. Effects of body size and parasite infection on the locomotory performance of juvenile toads. Bufo bufo. Oikos 1993, 66, 129–136. [Google Scholar] [CrossRef]
- Beck, C.W.; Congdon, J.D. Effects of age and size at metamorphosis on performance and metabolic rates of southern toad, Bufo terrestris, metamorphs. Funct. Ecol. 2000, 14, 32–38. [Google Scholar] [CrossRef]
- Gervasi, S.S.; Foufopoulos, J. Costs of plasticity: responses to desiccation decrease post-metamorphic immune function in a pond-breeding amphibian. Funct. Ecol. 2008, 22, 100–108. [Google Scholar]
- Downie, J.R.; Bryce, R.; Smith, J. Metamorphic duration: an under-studied variable in frog life histories. Biol. J. Linn. Soc. 2004, 83, 261–272. [Google Scholar] [CrossRef]
- Pough, F.H.; Taigen, T.L.; Stewart, M.L.; Brussard, P.F. Behavioral modification of evaporative water loss by a Puerto Rican frog. Ecology 1983, 64, 244–252. [Google Scholar] [CrossRef]
- Chan-McLeod, A.C.A. Factors affecting the permeability of clearcuts to red-legged frogs. J. Wild. Manag. 2003, 67, 663–671. [Google Scholar] [CrossRef]
- Roe, A.W.; Grayson, K.L. Terrestrial movements and habitat use of juvenile and emigrating adult Eastern red-spotted newts, Notophthalmus viridescens. J. Herpetol. 2008, 42, 22–30. [Google Scholar] [CrossRef]
- Ray, C. Vital limits and rates of desiccation in salamanders. Ecology 1958, 39, 75–83. [Google Scholar] [CrossRef]
- Wells, K.D. The Ecology and Behavior of Amphibians; The University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Karl, T.R.; Knight, R.W. Secular trends of precipitation amount, frequency, and intensity in the United States. Bull. Am. Meteorol. Soc. 1998, 79, 231–241. [Google Scholar] [CrossRef]
- National Assessment Synthesis Team. Climate Change Impacts on the United States: The Potential Consequences of Climate Variability and Change; U.S. Global Change Research Program: Washington, DC, USA, 2000. [Google Scholar]
- Brooks, R.T. Weather-related effects on woodland vernal pool hydrology and hydroperiod. Wetlands 2004, 24, 104–114. [Google Scholar] [CrossRef]
- Lake, P.S. Ecological effects of perturbation by drought in flowing waters. Freshwater Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Relationship of pond drying to the reproductive success of the salamander Ambystoma talpoideum. Copeia 1987, 1, 61–69. [Google Scholar] [CrossRef]
- Dodd, C.K., Jr. Cost of living in an unpredictable environment: the ecology of striped newts Notophthalmus perstriatus during a prolonged drought. Copeia 1993, 1, 605–614. [Google Scholar] [CrossRef]
- Dodd, C.K., Jr. The effects of drought on population structure, activity, and orientation of toads (Bufo quercicus and B. terrestris) at a temporary pond. Ethol. Ecol. Evol. 1994, 6, 331–349. [Google Scholar] [CrossRef]
- Richter, S.C.; Young, J.E.; Johnson, G.N.; Seigel, R.A. Stochastic variation in reproductive success of a rare frog, Rana sevosa: implications for conservation and for monitoring amphibian populations. Biol. Conserv. 2003, 111, 171–177. [Google Scholar] [CrossRef]
- Palis, J.G.; Aresco, M.J.; Kilpatrick, S. Breeding biology of a Florida population of Ambystoma cingulatum (Flatwoods salamander) during a drought. Southeast. Nat. 2006, 5, 1–8. [Google Scholar]
- Taylor, B.E.; Scott, D.E.; Gibbons, J.W. Catastrophic reproductive failure, terrestrial survival, and persistence of the marbled salamander. Conserv. Biol. 2006, 20, 792–801. [Google Scholar] [CrossRef]
- Patla, D.A.; Peterson, C.R.; Corn, P.S. Amphibian decline in Yellowstone National Park. Proc. Natl. Acad. Sci. USA 2009, 106, 22. [Google Scholar] [CrossRef]
- Michener, W.K.; Blood, E.R.; Bildstein, K.L.; Brinson, M.M.; Gardner, L.R. Climate change, hurricanes and tropical storms, and rising sea level in coastal wetlands. Ecol. Apps 1997, 7, 770–801. [Google Scholar] [CrossRef]
- Schriever, T.A.; Ramspott, J.; Crother, B.I.; Fontenot, C.L., Jr. Effects of hurricanes Ivan, Katrina, and Rita on a southeastern Louisiana herpetofauna. Wetlands 2009, 29, 112–122. [Google Scholar] [CrossRef]
- Palis, J.G. Element stewardship abstract: flatwoods salamander (Ambystoma cingulatum Cope). Nat. Areas J. 1996, 16, 49–54. [Google Scholar]
- Dodd, C.K.; Barichivich, W.J.; Johnson, S.A.; Staiger, J.S. Changes in a northwestern Florida gulf coast herpetofaunal community over a 28-y period. Am. Midl. Nat. 2007, 158, 29–48. [Google Scholar] [CrossRef]
- Spotila, J.R. Role of temperature and water in the ecology of lungless salamanders. Ecol. Monogr. 1972, 42, 95–124. [Google Scholar] [CrossRef]
- Bernardo, J.; Spotila, J.R. Physiological constraints on organismal response to global warming: mechanistic insights from clinally varying populations and implications for assessing endangerment. Biol. Lett. 2006, 2, 135–139. [Google Scholar] [CrossRef]
- Stroh, C.L.; DeSteven, D.; Guntenspergen, G.R. Effect of climate fluctuations on long-terms vegetation dynamics in Carolina Bay wetlands. Wetlands 2008, 28, 17–27. [Google Scholar] [CrossRef]
- Kloeppel, B.D.; Clinton, B.D.; Vose, J.M.; Cooper, A.R. Drought impacts on tree growth and mortality of southern Appalachian forests. In Climate Variability and Ecosystem Response at Long-Term Ecological Research Sites; Greenland, D., Goodin, D.G., Smith, R.C., Eds.; Oxford University Press: New York, NY, USA, 2003. [Google Scholar]
- Halverson, M.A.; Skelly, D.K.; Kiesecker, J.M.; Freidenburg, L.K. Forest mediated light regime linked to amphibian distribution and performance. Oecologia 2003, 134, 360–364. [Google Scholar]
- Binckley, C.A.; Resetarits, W.J., Jr. Effects of forest canopy on habitat selection in treefrogs and aquatic insects: implications for communities and metacommunities. Oecologia 2007, 153, 951–958. [Google Scholar] [CrossRef]
- Roznik, E.A.; Johnson, S.A. Canopy closure and emigration by juvenile gopher frogs. J. Wildlife Manag. 2009, 73, 260–268. [Google Scholar] [CrossRef]
- Rubbo, M.J.; Kiesecker, J.M. Leaf litter composition and community structure: translating regional species changes into local dynamics. Ecology 2004, 85, 2519–2525. [Google Scholar] [CrossRef]
- Williams, B.K.; Rittenhouse, T.A.G.; Semlitsch, R.D. Leaf litter input mediates tadpole performance across forest canopy treatments. Oecologia 2008, 155, 377–384. [Google Scholar] [CrossRef]
- Kirkman, L.K. Impacts of fire and hydrological regimes on vegetation in depressional wetlands of southeastern USA. In Fire in Wetlands: A Management Perspective. Proceedings of the Tall Timbers Fire Ecology Conference, No. 19; Cerulean, S.I., Engstrom, R.T., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 1995. [Google Scholar]
- Huxman, T.E.; Wilcox, B.P.; Breshears, D.D.; Scott, R.L.; Snyder, K.A.; Small, E.E.; Hultine, K.; Pockman, W.T.; Jackson, R.B. Ecohydrological implications of woody plant encroachment. Ecology 2005, 86, 308–319. [Google Scholar] [CrossRef]
- Altig, R.; Whiles, M.R.; Taylor, C.L. What do tadpoles really eat? Assessing the trophic status of an understudied and imperiled group of consumers in freshwater habitats. Freshwater Biol. 2007, 52, 386–395. [Google Scholar] [CrossRef]
- Meyer, J.L.; Sale, M.J.; Mulholland, P.K.; Hoff, N.L. Impacts of climate change on aquatic ecosystem functioning and health. J. Am. Water Res. Assoc. 1999, 35, 1373–1386. [Google Scholar] [CrossRef]
- Schindler, D.W. Widespread effects of climatic warming on freshwater ecosystems in North America. Hydrol. Process. 1997, 11, 1043–1067. [Google Scholar] [CrossRef]
- IPCC. Climate change and water. In Technical Paper of the Intergovernmental Panel on Climate Change; Bates, B.C., Kundzewicz, Z.W., Wu, S., Palutikof, J.P., Eds.; Secretariat: Geneva, Switzerland, 2008. [Google Scholar]
- Taylor, B.E.; Estes, R.A.; Pechmann, J.H.K.; Semlitsch, R.D. Trophic relations in a temporary pond: larval salamanders and their microinvertebrate prey. Can. J. Zool. 1988, 66, 2191–2198. [Google Scholar] [CrossRef]
- Leeper, D.A.; Taylor, B.E. Abundance, biomass and production of aquatic invertebrates in Rainbow Bay, a temporary wetland in South Carolina, USA. Arch. Hydrobiol. 1998, 143, 335–362. [Google Scholar]
- Durance, I.; Ormerod, S.J. Climate change effects on upland stream macroinvertebrates over a 25-year period. Glob. Change Biol. 2007, 13, 942–957. [Google Scholar] [CrossRef]
- Batzer, D.P.; Wissinger, S.A. Ecology of insect communities in nontidal wetlands. Ann. Rev. Entomol. 1996, 41, 75–100. [Google Scholar] [CrossRef]
- Brooks, R.T. Annual and seasonal variation and the effects of hydroperiod on benthic macroinvertebrates of seasonal forest (“vernal”) ponds in Central Massachusetts, USA. Wetlands 2000, 20, 707–715. [Google Scholar] [CrossRef]
- Taylor, B.E.; Leeper, D.A.; McClure, M.A.; DeBiase, A.E. Carolina Bays: Ecology of aquatic invertebrates and perspectives on conservation. In Invertebrates in Freshwater Wetlands of North America: Ecology and Management; Batzer, D.P., Rader, R.B., Wissinger, S.A., Eds.; John Wiley & Sons: New York, NY, USA, 1999; pp. 167–197. [Google Scholar]
- Jaeger, R.G. Fluctuations in prey availability and food limitation for a terrestrial salamander. Oecologia 1980, 44, 335–341. [Google Scholar] [CrossRef]
- Jaeger, R.G.; Wicknick, J.A.; Griffis, M.R.; Anthony, C.D. Socioecology of a terrestrial salamander: juveniles enter adult territories during stressful foraging periods. Ecology 1995, 76, 533–543. [Google Scholar]
- Walton, B.M. Salamanders in forest-floor food webs: environmental heterogeneity affects the strength of top-down effects. Pedobiologia 2005, 49, 381–393. [Google Scholar] [CrossRef]
- Milanovich, J.R.; Trauth, S.E.; Saugey, D.A.; Jordan, R.R. Fecundity, reproductive ecology, and influence of precipitation on clutch size in the Western Slimy Salamander (Plethodon albagula). Herpetologica 2006, 62, 292–301. [Google Scholar] [CrossRef]
- Scott, D.E.; Fore, M.R. The effect of food limitation on lipid levels, growth, and reproduction in the marbled salamander, Ambystoma opacum. Herpetologica 1995, 51, 462–471. [Google Scholar]
- Harris, R.N.; Ludwig, P.M. Resource level and reproductive frequency in female four-toed salamanders, Hemidactylium scutatum. Ecology 2004, 85, 1585–1590. [Google Scholar] [CrossRef]
- Cunningham, H.R.; Rissler, L.J.; Apodaca, J.J. Competition at the range boundary in the slimy salamander: using reciprocal transplants for studies on the role of biotic interactions in spatical distributions. J. Anim. Ecol. 2009, 78, 52–62. [Google Scholar] [CrossRef]
- Alford, R.A.; Wilbur, H.M. Priority effects in experimental pond communities: competition between Bufo and Rana. Ecology 1985, 66, 1097–1105. [Google Scholar] [CrossRef]
- Wilbur, H.M.; Alford, R.A. Priority effects in experimental pond communities: responses of Hyla to Bufo and Rana. Ecology 1985, 66, 1106–1114. [Google Scholar] [CrossRef]
- Alford, R.A. Variation in predator phenology affects predator performance and prey community composition. Ecology 1989, 70, 206–219. [Google Scholar] [CrossRef]
- Morin, P.J.; Lawler, S.P.; Johnson, E.A. Ecology and breeding phenology of larval Hyla andersonii: the disadvantages of breeding late. Ecology 1990, 71, 1590–1598. [Google Scholar] [CrossRef]
- Lawler, S.P.; Morin, P.J. Temporal overlap, competition, and priority effects in larval anurans. Ecology 1993, 74, 174–182. [Google Scholar] [CrossRef]
- Simberloff, D. Global climate change and introduced species in United States forests. Sci. Total Environ. 2000, 262, 253–261. [Google Scholar] [CrossRef]
- Kats, L.B.; Ferrer, R.P. Alien predators and amphibian declines: review of two decades of science and the transition to conservation. Divers. Distrib. 2003, 9, 99–110. [Google Scholar] [CrossRef]
- Rödder, D.; Weinsheimer, F. Will future anthropogenic climate change increase the potential distribution of the alien invasive Cuban treefrog (Anura: Hylidae)? J. Nat. Hist. 2009, 43, 1207–1217. [Google Scholar] [CrossRef]
- Kearney, M.; Phillips, B.L.; Tracy, C.R.; Christian, K.R.; Betts, G.; Porter, W.P. Modeling species distributions without using species distributions: the cane toad in Australia under current and future climates. Ecography 2008, 31, 423–434. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of a problematic alien invasive species—the American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Morin, P.J. Predatory salamanders reverse the outcome of competition among three species of anuran tadpoles. Science 1981, 212, 1284–1286. [Google Scholar]
- Fauth, J.E. Identifying potential keystone species from field data—an example from temporary ponds. Ecol. Lett. 1999, 2, 36–43. [Google Scholar]
- Fauth, J.E.; Resetarits, W.J., Jr. Interactions between the salamander Siren intermedia and the keystone predator Notophthalmus viridescens. Ecology 1991, 72, 827–838. [Google Scholar] [CrossRef]
- Smith, K.G. Keystone predators (eastern newts, Notophthalmus viridescens) reduce the impacts of an aquatic invasive species. Oecologia 2006, 148, 342–349. [Google Scholar] [CrossRef]
- Harvell, C.D.; Mitchell, C.; Ward, J.; Altizer, S.; Dobson, A.; Ostfeld, R.; Samuel, M. Climate warming and disease risks for terrestrial and marine biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef]
- Kilpatric, A.M.; Briggs, C.J.; Daszak, P. The ecology and impact of chytridiomycosis: an emerging disease of amphibians. Trends in Ecol. Evol. 2010, 25, 109–118. [Google Scholar] [CrossRef]
- Richards-Zawacki, C.L. Thermoregulatory behavior affects prevalence of chytrid fungal infection in a wild population of Panamanian golden frogs. Proc. Roy. Soc. B. 2010, 277, 519–528. [Google Scholar] [CrossRef]
- Green, G.; Cohen, N. Effect of temperature on serum complement levels in the leopard frog, Rana pipiens. Develop. Comp. Immunol. 1977, 1, 59–64. [Google Scholar] [CrossRef]
- Maniero, G.D.; Carey, C. Changes in selected aspects of immune function in the leopard frog, Rana pipiens, associated with exposure to cold. J. Comp. Physiol. B 1997, 167, 256–263. [Google Scholar] [CrossRef]
- Matutte, B.; Storey, K.B.; Knoop, F.C.; Conlon, J.M. Induction of synthesis of an antimicrobial peptide in the skin of the freeze-tolerant frog, Rana sylvatica, in response to environmental stimuli. FEBS Lett. 2000, 483, 135–138. [Google Scholar] [CrossRef]
- Wright, R.K.; Cooper, E.L. Temperature effects of ectotherm immune response. Develop. Comp. Immunol. 1991, 5, 117–122. [Google Scholar]
- Cooper, E.L.; Wright, R.K.; Klembau, A.E.; Smith, C.T. Hibernation alters the frog’s immune system. Cryobiology 1992, 29, 616–631. [Google Scholar] [CrossRef]
- Bly, J.E.; Clem, L.W. Temperature-mediated processes in teleost immunity: in vitro immunosuppression induced by in vivo low temperature in channel catfish. Vet. Immunol. Immunopathol. 1991, 28, 365–377. [Google Scholar] [CrossRef]
- Cone, R.E.; Marchalonis, J.J. Cellular and humoral aspects of the influence of environmental temperature on the immune response of poikilothermic vertebrates. J. Immunol. 1972, 108, 952–957. [Google Scholar]
- Le Morvan, C.; Deschaux, P.; Troutaud, D. Effects and mechanisms of environmental temperature on carp (cyprinus carpio) anti-DNP antibody response and non-specific cytotoxic cell activity: A kinetic study. Develop. Comp. Immunol. 1996, 20, 331–340. [Google Scholar] [CrossRef]
- Hardie, L.J.; Fletcher, T.C.; Secombes, C.J. Effect of temperature on macrophage activation and the production of macrophage activating factor by rainbow trout (Oncorhynchus mykiss) leucocytes. Develop. Comp. Immunol. 1994, 18, 57–66. [Google Scholar]
- Le Morvan, C.; Clerton, P.; Deschaux, P.; Troutaud, D. Effects of environmental temperature on macrophage activities in carp. Fish Shellfish Immunol. 1997, 7, 209–212. [Google Scholar] [CrossRef]
- Raffel, T.R.; Rohr, J.R.; Kiesecker, J.M.; Hudson, P.J. Negative effects of changing temperature on amphibian immunity under field conditions. Funct. Ecol. 2006, 20, 819–828. [Google Scholar] [CrossRef]
- Collazos, M.E.; Barriga, C.; Ortega, E. Effect of high summer temperatures upon granulocyte phagocytic function of the tench (Tinca Tinca, L.). Comp. Immunol. Microbiol. Infect. Dis. 1995, 18, 115–121. [Google Scholar] [CrossRef]
- Daszak, P.A.; Cunningham, A.; Hyatt, A.D. Infectious disease and amphibian population declines. Divers. Distrib. 2003, 9, 141–150. [Google Scholar]
- Skerratt, L.F.; Berger, L.; Speare, R.; Cashins, S.; McDonald, K.R.; Phillott, A.D.; Hines, H.B.; Kenyon, N. Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. EcoHealth 2007, 4, 125–134. [Google Scholar] [CrossRef]
- Rödder, D.; Kielgast, J.; Bielby, J.; Schmidtlein, S.; Bosch, J.; Garner, T.W.K.; Veith, M.; Walker, S.; Fisher, M.C.; Lötters, S. Global amphibian extinction risk assessment for the panzootic chytrid fungus. Diversity 2009, 1, 52–66. [Google Scholar]
- Burdon, J.; Elmqvist, T. Selective sieves in the epidemiology of Melampsora lini. Plant Pathol. 1996, 45, 933–943. [Google Scholar] [CrossRef]
- Dwyer, G.; Elkinton, J.S. Using simple models to predict virus epizootics in gypsy moth populations. J. Anim. Ecol. 1993, 62, 1–11. [Google Scholar]
- Blaustein, A.R.; Johnson, P.T.J. Prevalence, causes, and implications of amphibian deformities. Front. Ecol. Environ. 2003, 1, 87–94. [Google Scholar]
- Johnson, P.T.J.; Chase, J.M.; Dosch, K.L.; Hartson, R.B.; Gross, J.A.; Larson, D.J.; Sutherland, D.R.; Carpenter, S.R. Aquatic eutrophication promotes pathogenic infection in amphibians. Proc. Natl. Acad. Sci. 2007, 104, 15781–15786. [Google Scholar] [CrossRef]
- Johnson, M.L.; Berger, L.; Philips, L.; Speare, R. Fungicidal effects of chemical disinfectants, UV light, desiccation and heat on the amphibian chytrid Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2003, 57, 255–260. [Google Scholar] [CrossRef]
- Kriger, K.M. Lack of evidence for the drought-linked chytridiomycosis hypothesis. J. Wild. Dis. 2009, 45, 537–541. [Google Scholar] [CrossRef]
- Lampo, M.; Rodríguez-contreras, A.; La Marca, E.; Daszak, P. A chytridiomycosis epidemic and a severe dry season precede the disappearance of Atelopus species from the Venezuelan Andes. Herpetol. J. 2006, 16, 395–402. [Google Scholar]
- Longcore, J.E.; Pessier, A.P.; Nichols, D.K. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia 1999, 91, 219–227. [Google Scholar] [CrossRef]
- Piotrowski, J.S.; Annis, S.L.; Longcore, J.E. Physiology of Batrachochytrium dendrobatidis, a chytrid pathogen of amphibians. Mycologia 2004, 96, 9–15. [Google Scholar] [CrossRef]
- Seimon, T.A.; Seimon, A.; Daszak, P.; Halloys, S.R.P.; Schloegel, L.M.; Aguilar, C.A.; Sowell, P.; Hyatt, A.D.; Konecky, B.; Simmons, J.E. Upward range extension of Andean anurans and chytridiomycosis to extreme elevations in response to tropical deglaciation. Glob. Change Biol. 2007, 13, 288–299. [Google Scholar] [CrossRef]
- Pounds, A.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.M.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; Ron, S.R.; Sánchez-Azofeifa, G.A.; Still, C.J.; Young, B.E. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef]
- Kupferberg, S.J.; Catenazzi, A.; Lunde, K.; Lind, A.J.; Palen, W.J. Parasitic Copepod (Lernaea cyprinacea) Outbreaks in Foothill Yellow-legged Frogs (Rana boylii) Linked to Unusually Warm Summers and Amphibian Malformations in Northern California. Copeia 2009, 1, 529–537. [Google Scholar]
- Blaustein, A.R.; Dobson, A. Extinction: A message from the frogs. Nature 2006, 439, 143–144. [Google Scholar] [CrossRef]
- Bosch, J.; Carrascal, L.M.; Duran, L.; Walker, S.; Fisher, M.C. Climate change and outbreaks of amphibian chytridiomycosis in a montane area of Central Spain; is there a link? Proc. Roy. Soc. (B) 2006, 274, 253–260. [Google Scholar]
- D’Amen, M.; Bombi, P. Global warming and biodiversity: evidence of climate-linked amphibian declines in Italy. Biol. Conserv. 2009, 142, 3060–3067. [Google Scholar] [CrossRef]
- Drew, A.; Allen, E.J.; Allen, L.J.S. Analysis of climatic and geographic factors affecting the presence of chytridiomycosis in Australia. Dis. Aquat. Organ. 2006, 68, 245–250. [Google Scholar] [CrossRef]
- Lips, K.R.; Diffendorfer, J.; Mendelson, J.R., III; Sears, M.W. Riding the wave: Reconciling the roles of disease and climate change in amphibian declines. Plos Biol. 2008, 6, 441–454. [Google Scholar]
- Lampo, M.; Sánchez, D.; Nava-González, F.; García, C.Z.; Acevedo, A. Wavelike epidemics in Venezuela? Plos Bio. 2008. [Google Scholar]
- Parmesan, C.; Singer, M.C. Amphibian extinctions: Disease not the whole story. Plos Biol. 2008. [Google Scholar]
- Rohr, J.R.; Raffel, T.R.; Romansic, J.M.; McCallum, H.; Hudson, P.J. Evaluating the links between climate, disease spread, and amphibian declines. Proc. Natl. Acad. Sci. USA 2008, 105, 17436–17441. [Google Scholar]
- Alford, R.A.; Bradfield, K.S.; Richards, S.J. Global warming and amphibian losses. Nature 2007, 447, E3–E4. [Google Scholar] [CrossRef]
- Di Rosa, I.; Simoncelli, F.; Fagotti, A.; Pascolini, R. The proximate cause of frog declines? Nature 2007, 447, E4–E5. [Google Scholar] [CrossRef]
- Pounds, J.A.; Bustamante, M.R.; Coloma, L.A. Pounds et al. reply. Nature 2007, 447, E5–E6. [Google Scholar] [CrossRef]
- Bancroft, B.A.; Baker, N.J.; Blaustein, A.R. A meta-analysis of the effects of ultraviolet B radiation and other stressors on survival in amphibians. Conser Biol. 2008, 22, 987–996. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Kiesecker, J.M.; Chivers, D.P.; Hokit, D.G.; Marco, A.; Belden, L.K.; Hatch, A. Effects of ultraviolet radiation on amphibians: Field experiments. Am. Zool. 1998, 38, 799–812. [Google Scholar]
- Nagl, A.M.; Hofer, R. Effects of ultraviolet radiation on early larval stages of the Alpine newt, Triturus alpestris, under natural and laboratory conditions. Oecologia 1997, 110, 514–519. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Chivers, D.P.; Kats, L.B.; Kiesecker, J.M. Effects of ultraviolet radiation on locomotion and orientation in roughskin newts (Taricha granulosa). Ethology 2000, 108, 227–234. [Google Scholar]
- Kats, L.B.; Kiesecker, J.M.; Chivers, D.P.; Blaustein, A.R. Effects of UV-B on antipredator behavior in three species of amphibians. Ethology 2000, 106, 921–932. [Google Scholar] [CrossRef]
- Belden, L.K.; Wildy, E.L.; Blaustein, A.R. Growth, survival, and behaviour of larval long-toed salamanders (Ambystoma macrodactylum) exposed to ambient levels of UV-B radiation. J. Zool. (London) 2000, 251, 473–479. [Google Scholar] [CrossRef]
- Pahkala, M.; Laurila, A.; Merilä, J. Ambient ultraviolet-B radiation reduces hatchling size in the common frog Rana temporaria. Ecography 2000, 23, 531–538. [Google Scholar] [CrossRef]
- Pahkala, M.; Laurila, A.; Merilä, J. Carry-over effects of ultraviolet-B radiation on larval fitness in Rana temporaria. Proc. Roy. Soc. London B 2001, 268, 1699–1706. [Google Scholar] [CrossRef]
- Smith, G.R.; Waters, M.A.; Rettig, J.E. Consequences of embryonic UV-B exposure for embryos and tadpoles of the plains leopard frog. Conserv. Biol. 2000, 14, 1903–1907. [Google Scholar] [CrossRef]
- Worrest, R.D.; Kimeldorf, D.J. Distortions in amphibian development induced by ultraviolet-B enhancement (290−310 nm) of a simulated solar spectrum. Photochem. Photobiol. 1976, 24, 377–382. [Google Scholar] [CrossRef]
- Hays, J.B.; Blaustein, A.R.; Kiesecker, J.M.; Hoffman, P.D.; Pandelova, I.; Coyle, I.C.; Richardson, T. Developmental responses of amphibians to solar and artificial UV-B sources: A comparative study. Photochem. Photobiol. 1996, 64, 449–456. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Kiesecker, J.M.; Chivers, D.P.; Anthony, R.G. Ambient UV-B radiation causes deformities in amphibian embryos. Proc. Natl. Acad. Sci. USA 1997, 94, 13735–13737. [Google Scholar] [CrossRef]
- Bruggeman, D.J.; Bantle, J.A.; Goad, C. Linking teratogenesis, growth, and DNA photodamage to artificial ultraviolet-B radiation in Xenopus laevis larvae. Environ. Toxicol. Chem. 1998, 17, 2114–2121. [Google Scholar] [CrossRef]
- Fite, K.V.; Blaustein, A.R.; Bengston, L.; Hewitt, H.E. Evidence of retinal light damage in Rana cascadae: a declining amphibian species. Copeia 1998, 1, 906–914. [Google Scholar]
- Croteau, M.C.; Davidson, M.A.; Lean, D.R.S.; Trudeau, V.L. Global increases in ultraviolet B radiation: Potential impacts on amphibian development and metamorphosis. Physiol. Biochem. Zool. 2008, 81, 743–761. [Google Scholar] [CrossRef]
- Kiesecker, J.M.; Blaustein, A.R. Synergism between UV-B radiation and a pathogen magnifies amphibian embryo mortality in nature. Proc. Natl. Acad. Sci. USA 1995, 92, 11049–11052. [Google Scholar] [CrossRef]
- Long, L.E.; Saylor, L.S.; Soule, M.E. A pH/UV-B synergism in amphibians. Conserv. Biol. 1995, 9, 1301–1303. [Google Scholar]
- Merilä, J.; Pahkala, M.; Johanson, U. Increased ultraviolet-B radiation, climate change and latitudinal adaptation-a frog perspective. Annales Zool. Fenn. 2000, 37, 129–134. [Google Scholar]
- Van Uitregt, V.O.; Wilson, R.S.; Franklin, C.E. Cooler temperatures increase sensitivity to ultraviolet B radiation in embryos and larvae of the frog Limnodynastes peronii. Glob. Change Biol. 2007, 13, 1114–1121. [Google Scholar] [CrossRef]
- Searle, C.L.; Belden, L.K.; Bancroft, B. A.; Han, B.A.; Biga, L.M.; Blaustein, A.R. Experimental examination of the effects of ultraviolet-B radiation in combination with other stressors on frog larvae. Oecologia 2009, 162, 237–245. [Google Scholar]
- Tilman, D.; Fargione, J.; Wolff, B. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef]
- Davidson, C.; Shaffer, H.B.; Jennings, M.R. Declines of the California red-legged frog: Climate, UV-B, habitat, and pesticides hypotheses. Ecol. App. 2001, 11, 464–479. [Google Scholar] [CrossRef]
- Davidson, C.; Knapp, R. Multiple stressors and amphibian declines: dual impacts of pesticides and fish on yellow-legged frogs. Ecol. App. 2007, 17, 587–597. [Google Scholar] [CrossRef]
- Wright, R.F.; Schindler, D.W. Interaction of acid rain and global changes: effects on terrestrial and aquatic ecosystems. Water Air Soil Poll. 1995, 85, 89–99. [Google Scholar] [CrossRef]
- Zabik, J.M.; Seiber, J.N. Atmospheric transport of organophosphate pesticides from California’s Central Valley to the Sierra Nevada mountains. J. Environ. Qual. 1993, 22, 80–90. [Google Scholar] [CrossRef]
- Aston, L.S.; Seiber, J.N. Fate of summertime airborne organophosphate pesticide residues in the Sierra Nevada Mountains. J. Environ. Qual. 1997, 26, 1483–1492. [Google Scholar] [CrossRef]
- Davidson, C. Declining downwind: Amphibian population declines in California and historical pesticide use. Ecological Applications 2004, 14, 1892–1902. [Google Scholar] [CrossRef]
- Hageman, K.J.; Simonich, S.L.; Campbell, D.H.; Wilson, G.R.; Landers, D.H.; Dunson, W.A.; Wyman, R.L. Atmospheric deposition of current-use and historic-use pesticides in snow at national parks in the western United States. Environ. Sci. Technol. 2006, 40, 3174–3180. [Google Scholar] [CrossRef]
- Kiesecker, J.M. pH induced growth reduction and its effects on predator-prey interactions between Ambystoma tigrinum and Pseudacris triseriata. Ecol. App. 1996, 6, 1325–1331. [Google Scholar]
- Pounds, J.A.; Crump, M.L. Amphibian declines and climate disturbance: the case of the golden toad and the harlequin frog. Conserv. Biol. 1994, 8, 72–85. [Google Scholar]
- Sparling, D.W.; Fellers, G.M.; McConnell, L.L. Pesticides and amphibian population declines in California, USA. Environ. Tox. Chem. 2001, 20, 1591–1595. [Google Scholar]
- Blaustein, A.R.; Alford, R.A.; Harris, R.N. The value of well designed experiments in studying diseases with special reference to amphibians. EcoHealth 2009, in press. [Google Scholar]
- Maurer, E.P.; Adam, J.C.; Wood, A.W. Climate model based consensus on the hydrologic impacts of climate change to the Rio Lempa basin of Central America. Hydrol. Earth Sys. Sci. 2009, 13, 183–194. [Google Scholar] [CrossRef]
- Girvetz, E.; Zganjar, C.; Raber, G.; Shafer, S.L.; Maurer, E.P.; Kareiva, P.; Lawler, J.J. Applied climate-change analysis. PLoS ONE 2009, 4, e8320. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).