Abstract
Temperature affects the morphology, physiology, and distribution of marine macroalgae, as supported by studies that used long-term data from herbaria. In the present study, sea surface temperature (SST), latitudinal distribution, and La Niña or El Niño years were correlated to the morphology of two macroalgal species of the Mexican Pacific: Ceratodictyon tenue and C. variabile. Twenty-four morphological characteristics were evaluated, and 95 samples from 1965 to 2013 in the Escuela Nacional de Ciencias Biológicas herbarium were reviewed. In 2017, 2023, and 2024, 12 specimens were sampled at three locations. Low positive correlations were found between thallus diameter and SST for C. tenue, while low positive correlations were detected for thallus length and medullary cell diameter vs. SST and medullary cell length vs. year for C. variabile. Significant relationships were found between the thallus length and cortical cell diameter of C. variabile with latitude and SST. It is concluded that SST contributes to changes in morphology, but is not the only factor that affects them. For the first time in a tropical area, the present study explores whether there is a relationship between SST, latitudinal distribution, and El Niño and La Niña years and the morphology of a genus of red algae.
1. Introduction
Sea surface temperature (SST) and its relationship with ocean currents has historically shaped the latitudinal distribution of marine species that respond by increasing in abundance and spatial distribution [1]. Temperature is related to fundamental physiological processes from the cellular level to organ functioning. Marine organisms mostly live close to their thermal tolerance limits; thus, temperature increases negatively affect their performance and survival [2]. Temperature is a key factor that determines the spatial distribution and physiological rates of marine macrophytes [3]. Climate change is occurring at an unprecedented rate, with rates up to seven times higher in the ocean than on land [4]. As the planet continues to warm up due to increasing atmospheric greenhouse gas concentrations, the ocean faces a variety of disturbances, including warming, acidification, decreasing oxygen concentrations, reduction of sea ice, and rising sea levels [5].
There is evidence that the increase in SST affects different aspects of marine macroalgae, such as metabolism (especially at the level of photosynthesis or growth), cell damage, changes in stem morphology (reduction in size, branching patterns), thalli degradation, changes in phenology, latitudinal and vertical distribution, and community structure. The worst-case scenario is the extinction of several hundred macroalgal species [6,7,8,9,10,11,12,13].
Understanding how human stressors have affected biodiversity [14] over time is crucial for understanding the biological impact [15] they have had in the past, and for making predictions on how biodiversity will respond to future global environmental changes [16]. Data from herbaria are traditionally used for taxonomic purposes, but they are also important as historical datasets for assessing recent changes in marine community assemblages [1], providing unique information for studying long-term changes [16]. Recently, the use of data from herbaria has gained relevance for the detection of morphological changes in organisms, which, on several occasions, have been related to the impact of climate change, particularly increased SST [17]. These historical data reveal patterns that would not otherwise be observable from other data sources [18,19]. In the last decade or so, there has been an increasing recognition of the value of collections in the analysis of biodiversity, with interest in their potential applications, for example, for inferring threats to conservation and ecology associated with anthropogenic changes [8,19,20,21,22].
Studies have been carried out using herbarium data to evaluate changes in the morphology and floristic composition of marine macroalgae, such as the study by Riera et al. [1], who evaluated Fucus limitaneus (formerly Fucus guiryi) (Phaeophyceae) in the Canary Islands using long-term herbarium material. They recognized that the decrease in the size of the morphological characteristics, and even the decline in F. limitaneus populations, were related to environmental variables such as temperature. Later, in the same area, Alfonso et al. [23] evaluated the changes in the morphology and reproductive aspects of Gelidium canariense (Rhodophyta) over time and found that the length of the thallus has been reduced by half over the last 40 years, coinciding with a significant increase in SST, air temperature above the sea surface, and ultraviolet radiation. Geppi and Riera [3] studied the long-term variability (38-year period, 1978–2016) of the morphological characteristics of three fucoid species, Gongolaria abies-marina, Cystoseira compressa, and C. humilis (Phaeophyceae), and recorded significant declines in thallus size, reproductive efficiency, and biomass, indicating the algae’s disappearance in some of the studied areas.
Mexico’s Pacific coast makes up 68% of the country’s coastline and comprises several different ecoregions [24] at local, regional, and global levels [25], thus covering a particularly wide range of habitats [26]. This results in substantial coastal and marine diversity. The Mexican Pacific is among the most impacted sites due to various anthropogenic pressures that significantly affect biodiversity, such as habitat modification and loss, pollution, fisheries, coastal urbanization, tourism, overexploitation, species introduction, and climate change [25,27]. The Mexican Pacific is strongly influenced by large and mesoscale processes of the Pacific Ocean that closely interact with climate conditions [24]. The warm and cold episodes of large-scale atmospheric circulation [28] and the El Niño/Southern Oscillation (ENSO) [29] are among these processes, and their variations associated with recent climate change can seriously affect the coastal zones of the Mexican Pacific [25].
The genus Ceratodictyon has 10 described species worldwide [30], of which C. variabile (J. Agardh) R.E. Norris and C. tenue (Setchell & N.L. Gardner) J.N. Norris (Rhodophyta) have been recorded throughout the Mexican Pacific since 1950 [31,32,33,34]. Additionally, both species can be easily distinguished from each other by their morphological characteristics. As they are representative, frequent species and have clearly delimited morphological characteristics, they have been used in the present study to determine whether there was a relationship between morphology, SST, and the El Niño and La Niña events over time. Herbarium samples and recently collected material from the Mexican Pacific region were used in the analysis. This is the first study of its kind conducted in a tropical area, the aim of which is to begin to evaluate the possible existence and degree of influence, if any, of factors such as SST on macroalgae.
2. Materials and Methods
2.1. Collection Site
Ceratodyction tenue and C. variabile were collected from the following locations in the Mexican Pacific: Sayulita and Nayarit in March 2017; San Agustín Bay and Santa Elena Bay, Oaxaca, in March 2023 and September 2024; and El Mechudo Point, Baja California Sur, in April 2024 (Figure 1). At each locality, we randomly collected specimens manually with a spatula in the intertidal rocky zone between the mid- and high-water marks. Samples were preserved in a mix of formaldehyde (4%) and sea water until their preparation in the laboratory. The collected macroalgae were identified at the species level using identification keys [31,32,35].
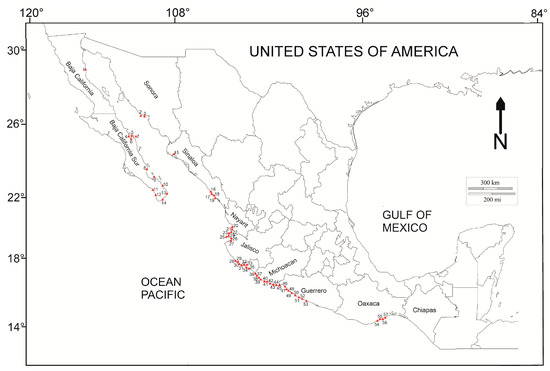
Figure 1.
Sites at which the reviewed specimens of Ceratodyction tenue and C. variabile were collected.
2.2. Herbarium Data
For this work, we used herbarium specimens taken from 55 locations in the Mexican Pacific (Figure 1) that were collected during the study period 1965–2013. These samples were obtained from specimens deposited in the phycological collection of the Herbario de la Escuela Nacional de Ciencias Biológicas of the Instituto Politécnico Nacional (ENCB-IPN) [36].
2.3. Morphological Features
Several morphological characteristics were measured for each herbarium and field-collected specimen, and 18 vegetative and six reproductive characteristics were evaluated following the methods described in other studies [32,37] (Table 1, Figure 2).

Table 1.
Morphological characteristics of Ceratodictyon species examined in herbarium and field-collected specimens. * = characteristics not included in the analysis due to lack of variation or absence in several specimens.

Figure 2.
Thalli of the Ceratodictyon species analyzed in this study. (a) Thallus of C. variabile. (b) Thallus of C. tenue. (c) Cross-section of C. variabile showing cortical cells (cc) and medullary cells (cm); the arrow indicates the reduction of medullary cells towards the center of the thallus. (d) Cross-section of C. tenue, showing cortical cells (cc) and medullary cells (cm); the arrow indicates the reduction of medullary cells towards the center of the thallus.
Transverse and longitudinal thallus sections were prepared manually using a razor blade under a LEICA stereomicroscope (Leica Microsystems AG, Wetzlar, Germany). Anatomical observations were made using a LEICA DM500 optical microscope (Leica Microsystems AG, Wetzlar, Germany).
A matrix of the data from the evaluated characteristics was created in Excel. Ranges and averages were obtained from the quantitative data, and coding was performed for the qualitative data based on the states of each characteristic.
2.4. Sea Surface Temperature
The SST data were obtained from the Marine-Coastal Information and Analysis System of the Comisión Nacional para el Conocimiento y Uso de la Biodiversidad [38]. Data for La Niña, El Niño, and neutral years were obtained from OMM [39].
2.5. Statistical Analysis
The morphological variables for average thallus length, diameter, average size medullar cells, average size cortical cells, and average length of cortical cells were selected for univariate linear regression to detect a correlation with latitude, longitude, sampling year, and/or average annual SST. The analyses were performed separately for each species. RStudio 2022.07.2+576 software [40] was used to perform the linear regressions with the function ‘lm’ and a formula expressing each morphological variable as the result of the effect of latitude, longitude, year, or average annual SST at the sampling site:
where morvar is the selected morphological variable; lat is latitude; long is longitude; year is the sampling year; and temp is the average annual SST at the site. The variables that were deemed insignificantly correlated were removed to obtain linear models that only considered variables with a significant correlation.
lm = morvar ~ lat + long + year + temp
Two-way permutational analyses of variance (PERMANOVA) were performed on the morphological variables for each species by employing Euclidean distances, the type of ENSO year, and the Mexican states as groups to determine whether significant differences exist among Ceratodictyon populations that might be related to the ENSO phenomenon. The type of ENSO year was selected as a group due to the different effects this phenomenon has on sea temperature, while the Mexican states were selected as groups to account for the different populations of Ceratodictyon distributed along the Mexican Pacific coast. When the two-way PERMANOVA detected significant differences attributed to ENSO years or states, a one-way PERMANOVA was carried out alongside a paired PERMANOVA as a post-hoc test to determine the groups with significant statistical differences. All PERMANOVAs were performed independently using Past v. 4.17 [41] software for each morphological variable and species.
3. Results
A total of 107 specimens of Ceratodictyon (53 C. tenue and 54 C. variabile) from the Mexican Pacific (Houston, TX, USA) were reviewed, of which 95 were deposited in the ENCB-IPN herbarium and 12 were obtained from recent collecting in the field. Samples from 57 localities were reviewed (Figure 1) (Supplementary File). Of the total number of characteristics evaluated, reproductive characteristics were not considered, since only two samples were in the tetrasporic phase, and the remainder were vegetative. Fourteen vegetative characteristics were considered to have significant variation, with thallus size and diameter and size of the medullary and cortical cells being the most strongly related to changes in SST (Table 1, Figure 2). SST has increased throughout the Mexican Pacific since 1965 (Figure 2), with an approximate increase of 0.5 °C per decade, and a positive correlation was found (R2 = 0.7226) (Figure 3).
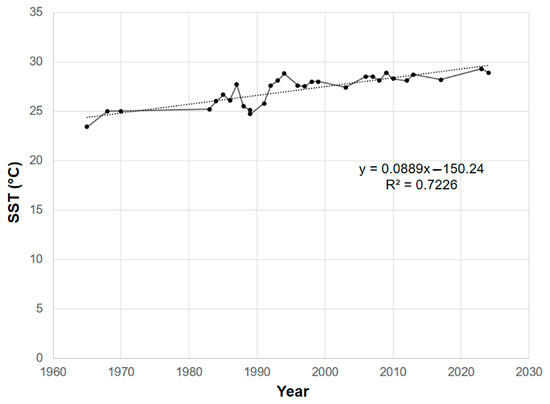
Figure 3.
Average annual sea surface temperature (SST) variation by year from 1965 to 2024.
3.1. Linear Regressions
For C. tenue, no significant differences were observed between SST and morphological characteristics except for thallus diameter. A slight tendency toward a decrease in thallus diameter with increasing temperature was observed. The linear regression of the average thallus diameter of C. tenue on the annual temperature was statistically significant (F = 5.04, df = 45, p = 0.02973), but showed a low correlation (R2 = 0.10).
The linear regression of the average medullar cell diameter of C. variabile on the annual temperature was statistically significant (F = 7.668, df = 41, p = 0.008402), but showed a low correlation (R2 = 0.15). Similarly, the correlation of the average cortical cell length with the year was statistically significant (F = 8.225, df = 41, p = 0.006498), but the correlation was weak (R2 = 0.16).
The correlations of the average cortical cells of C. variabile with latitude and longitude were statistically significant (F = 4.061, df = 40, p = 0.02479), but weak (R2 = 0.16). Moreover, the correlations of the average length of the thallus of C. variabile with latitude and annual temperature were also statistically significant (F = 5.246, DF = 40, p = 0.009479), but weak (R2 = 0.20). In the multiple linear regression, the average diameter was weakly correlated (R2 = 0.23) with annual temperature and the thallus length of C. variabile, but the regression was significant (F = 5.99, df = 40, p = 0.005301).
3.2. PERMANOVAS
A significant effect of the factor Mexican state (F = 1.9523, p = 0.0146) and the interaction between Mexican state and ENSO (F = 1.2232, p = 0.0041) were found for the average diameter of C. tenue. The ENSO factor alone did not have a significant effect (F = 0.3410, p = 0.5829). Significant effects of the factors Mexican state (F = 1.672, p = 0.0233) and ENSO (F = 4.3442, p = 0.0038) on the average number of medullary cells were observed. A significant effect of the factor ENSO (F = 3.1859, p = 0.0174) on the average number of cortical cells was found (Table 2).
Table 2.
Two-way permutational analysis of variance of the thallus characteristics of Ceratodictyon tenue, comparing the morphological characteristics with the greatest variation in the El Niño and La Niña events and Mexican states where the thalli were sampled. * = value statistically significant (p < 0.05).
One-way PERMANOVA was performed to evaluate the individual effects of the factors Mexican state and ENSO on some C. tenue morphological variables. Significant effects were found for Mexican state (F = 2.541, p = 0.0212) and ENSO (F = 6.225, p = 0.0048) on average medullary cell count. A significant effect of ENSO (F = 5.172, p = 0.0107) on the average cortical cell count was observed. The effect of Mexican state on average thallus diameter was not significant (F = 1.82, p = 0.1006) (Table 3).
Table 3.
One-way permutational analysis of variance of the thallus characteristics of Ceratodictyon tenue, comparing the morphological traits with the greatest variation during El Niño and La Niña events and Mexican states where the thalli were sampled. * = value statistically significant (p < 0.05).
In the paired PERMANOVA (post-hoc) for the effect of Mexican state on mean medullar cell count and the effect of ENSO on mean medullary and cortical cell counts, significant main effects were found in the one- and two-way PERMANOVAs (Table 4).
Table 4.
Paired permutational analysis of variance for the morphological characteristsics of the thallus of Ceratodictyon tenue between Mexican states.
Significant differences (p < 0.05) in the mean medullary cell count were found between the following pairs of Mexican states. Significant differences in mean medullary cell count were found between the following paired ENSO phases: La Niña–neutral (p = 0.0203) and La Niña–El Niño (p = 0.0022). A significant difference was also found in mean cortical cell count between the neutral ENSO phases and El Niño (p = 0.0020).
For C. variabile, a significant effect of the factor Mexican state (F = 5.7423, p = 0.0002) on the average thallus length was found, as well as the effect of the factor ENSO (F = 1.1874, p = 0.1782). A significant effect of Mexican state (F = 2.8235, p = 0.0013) on the average thallus diameter was observed. A significant effect of Mexican state (F = 5.5876, p = 0.0001) on the average number of cortical cells was found (Table 5). The one-way PERMANOVA showed significant effects of Mexican state on the average thallus length (F = 8.099, p = 0.0001), average thallus diameter (F = 3.905, p = 0.0017), and average cortical cell count (F = 7.153, p = 0.0001) (Table 6).
Table 5.
Two-way permutational analysis of variance of thallus characteristics of Ceratodictyon variabile, comparing the morphological characters with the greatest variation in the El Niño and La Niña events and Mexican states where the thalli were sampled. * = value statistically significant (p < 0.05).
Table 6.
One-way permutational analysis of variance of the thallus characteristics of Ceratodictyon variabile, comparing the morphological traits with the greatest variation during El Niño and La Niña events and Mexican states where the thalli were sampled. * = value statistically significant (p < 0.05).
The paired PERMANOVA (post-hoc) for the effects of Mexican state on the average thallus length and diameter and the average cortical cell were significantly different for the variables in which significant main effects were found in the one- and two-way PERMANOVAs. Significant differences (p < 0.05) were found in the average thallus length and diameter and the average cortical cell count between pairs of states (Table 7).
Table 7.
Paired permutational analysis of variance for the morphological characteristics of the thallus of Ceratodictyon variabile between Mexican states.
4. Discussion
The evaluation of Ceratodictyon thalli over a long period of time reflects the relationships between some morphological characteristics and the average annual SST and El Niño and La Niña events. In several cases, the correlations were significant but low, which may indicate that these factors are not the only ones involved. Among the morphological characteristics that exhibited the greatest change were thallus size, diameter, and medullary and cortical cell diameters, with a slight tendency for these to decrease, which is related to the gradual increase in temperature. Changes in these characteristics in populations of brown algae have been noted by other authors, particularly Fucales and Laminariales [42], such as in Fucus guiryi [43], Gongolaria abies-marina, Cystoseira compressa, and C. humilis [3], as well as in populations of red algae of the order Gelidiales, including Gelidium amansii [44], G. canariense, G. arbuscula, Pterocladiella capillacea [23,45,46], and Gelidium robustum [47]. Studies of macroalgae impacted by global warming are scarce, and those that do exist have been carried out in temperate zones. Therefore, studies in tropical zones are very important in order to recognize whether the impact is similar, lesser, or greater. Based on our observations and those of other authors, it appears that SST is not the predominant factor, as it very likely increases its effect when combined with other factors [1,43].
Morphological changes in response to environmental variables are very common among macroalgae, and SST has been identified as one of the main drivers of the changes in macroalgal development, abundance, and biomass [3,43,47]. Previous studies have indicated a significant relationship between temperature increases and drastic reductions in the size and reproductive capacity of organisms [23,43,48]. When under thermal stress, some macroalgal species undergo a process of metabolic acclimation, mainly with regard to photosynthesis and cellular respiration. However, this limits their growth, resulting in reduced thalli [3]. Increased ocean temperature can have a stressful effect on species that modify their morphological structure due to physiological changes caused by changes in energy budgets [11,42].
Regarding the relationship with SST, significant differences were observed in the diameters of the medullary and cortical cells of C. tenue during El Niño, La Niña, and neutral years. Larger diameters were observed in La Niña and neutral years compared to El Niño years. El Niño and La Niña events affect the morphology, abundance, and composition of macroalgal populations [47,49,50]. Hernández-Guerrero et al. [47] showed the impact of these meteorological events directly on the development and morphological characteristics of red algae. Important changes in the size, branching pattern, and reproduction of Gelidium robustum occurred from 1980 to 1990, during which there were El Niño and La Niña events. The increase in temperature in an El Niño year had a greater effect on the thalli, reducing the size and branching patterns and altering the life cycle of this alga. During the La Niña event, larger thalli were present, which coincides with the results of our study, in which larger diameters occurred in the medullary and cortical cells during La Niña years, followed by neutral years, with the smallest sizes in El Niño years. This may be due to lower temperatures occurring during La Niña years, which favors the presence of nutrients and even upwellings in many places [47,51]. This type of thallus alteration caused by rising temperatures has been documented in various genera of macroalgae, mainly in temperate zones or for algae of commercial interest, leaving a significant information gap for tropical species and those that have no direct economic use [47].
Furthermore, a significant relationship was observed between specimen distribution and morphology, which is linked to the mosaic of local climatic conditions, with important implications for the thermal sensitivity of organisms. Climatic variability can influence the range of thermal tolerance and is likely an important component of locally adapted responses to warming [52]. This has been observed in other algae, such as Fucus, in which significant changes in the morphology of nearby populations have been observed. The value of areas where environmental conditions represent a refuge for macroalgae is highlighted [43], as this may occur in some sites in Mexico for Ceratodictyon. Sites such as Michoacán have considerably larger thalli despite being exposed to high temperature conditions.
The low correlations observed may be related to the data used, since there are some biases regarding the number of specimens per location and per year, in addition to a significant period of time without information ranging from 2013 to 2017 and then from 2017 to 2023. Therefore, consecutive samplings should be carried out to determine significant changes in macroalgal populations, especially in tropical areas where climate change is not considered to have such a drastic effect, but is undocumented.
Although C. tenue and C. variabile are important components of rocky shorelines, much remains unknown regarding their physiology. However, our study presents evidence that distribution, El Niño and La Niña events, and SST have moderate to slight effects on these organisms. Long-term studies are needed to correlate these factors with other factors to detect the influence each may have on morphology.
5. Conclusions
In this study, we report the relationship between SST, El Niño, and La Niña events, as well as the latitudinal distribution and morphology of two red algal species of the genus Ceratodictyon from 1965 to 2024. This is the first time this type of study has been conducted with red algae in a tropical region, such as the Mexican Pacific. It is necessary to further explore the factors influencing these morphological changes and whether they can be attributed to climate change, as has occurred in other parts of the world.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d17080523/s1, Table S1. Data for the specimens reviewed in this work, annual average temperature, and climatic phenomena by year.
Author Contributions
Conceptualization, N.Q.-G. and L.E.M.-C.; Methodology, N.Q.-G., L.G.A.-E., O.O.-R.; Software, B.C.-C. and L.G.A.-E.; Validation, N.Q.-G., L.E.M.-C. and A.C.M.-G.; Formal Analysis, N.Q.-G., L.G.A.-E. and B.C.-C.; Investigation, N.Q.-G. and L.G.A.-E.; Resources, A.C.M.-G. and L.E.M.-C.; Data Curation, N.Q.-G. and L.G.A.-E.; Writing—Original Draft Preparation, N.Q.-G., L.G.A.-E., L.E.M.-C. and A.C.M.-G.; Writing—Review & Editing, N.Q.-G., L.G.A.-E., L.E.M.-C. and A.C.M.-G.; Visualization, N.Q.-G., L.G.A.-E., L.E.M.-C. and A.C.M.-G.; Supervision, L.E.M.-C. and A.C.M.-G.; Project Administration, N.Q.-G., L.E.M.-C. and A.C.M.-G.; Funding Acquisition, N.Q.-G., L.E.M.-C. and A.C.M.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors upon request.
Acknowledgments
We thank Fernando Acosta Ruiz for his support in processing the samples and Julio A. Acosta-Calderón for his support with the collection.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviation
The following abbreviations are used in this manuscript:
| SST | Sea Surface Temperature |
References
- Riera, R.; Sangil, C.; Sansón, M. Long-term herbarium data reveal the decline of a temperate-water algae at its southern range. Estuar. Coast. Shelf Sci. 2015, 165, 159–165. [Google Scholar] [CrossRef]
- Harley, C.D.; Randall Hughes, A.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.; Thornber, C.S.; Williams, S.L. The impacts of climate change in coastal marine systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef]
- Geppi, E.F.; Riera, R. Responses of intertidal seaweeds to warming: A 38-year time series shows differences of sizes. Estuar. Coast. Shelf Sci. 2022, 270, 107841. [Google Scholar] [CrossRef]
- Schultz, L.; Wessely, J.; Dullinger, S.; Albano, P.G. The climate crisis affects Mediterranean marine molluscs of conservation concern. Divers. Distrib. 2023, 30, e13805. [Google Scholar] [CrossRef]
- Hu, N.; Yu, Z.; Huang, Y.; Liu, D.; Wang, F.; Zhang, T. Elevated temperatures increase growth and enhance foraging performances of a marine gastropod. Aquac. Environ. Interact. 2021, 13, 177–188. [Google Scholar] [CrossRef]
- Davison, I.; Pearson, G. Stress tolerance in intertidal seaweeds. J. Psychol. 1996, 32, 197–211. [Google Scholar] [CrossRef]
- Rilov, G.; Treves, H. Climate change effects on marine ecological communities. In Role of Seaweeds in Future Globally Changing Environments. Cell Origin; Seckbach, J., Einav, R., Israel, A., Eds.; Springer: Berlin, Germany, 2010. [Google Scholar] [CrossRef]
- Wernberg, T.; Russell, B.D.; Thomsen, M.S.; Gurgel, C.F.D.; Bradshaw, C.J.A.; Poloczanska, E.S.; Connell, S.D. Seaweed communities in retreat from ocean warming. Curr. Biol. 2011, 21, 1828–1832. [Google Scholar] [CrossRef]
- Eggert, A. Seaweed Responses to Temperature. In Seaweed Biology. Ecological Studies; Wiencke, C., Bischof, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; 219p. [Google Scholar] [CrossRef]
- Clarke, A. Costs and consequences of evolutionary temperature adaptation. Trends Ecol. Evol. 2003, 18, 573–581. [Google Scholar] [CrossRef]
- Ji, Y.; Gao, K. Effects of climate change factors on marine macroalgae: A review. Adv. Mar. Biol. 2021, 88, 91–136. [Google Scholar] [CrossRef]
- Monteiro, C.; Pereira, J.; Seabra, R.; Lima, F.R. Fine-scale survey of intertidal macroalgae reveals recent changes in a cold-water biogeographic stronghold. Front. Mar. Sci. 2022, 9, 880074. [Google Scholar] [CrossRef]
- Tegner, M.; Dayton, P. El Nino effects on southern California kelp forest communities. Adv. Ecol. Res. 1987, 17, 243–279. [Google Scholar] [CrossRef]
- Pan, J.; Marcoval, M.A.; Bazzini, S.M.; Vallina, M.V.; Marco, S. Coastal marine biodiversity: Challenges and threats. In Marine Ecology in a Changing World; CRC Press: Boca Raton, FL, USA, 2013; pp. 43–67. [Google Scholar] [CrossRef]
- Gissi, E.; Manea, E.; Mazaris, A.D.; Fraschetti, S.; Almpanidou, V.; Bevilacqua, S.; Katsanevakis, S. A review of the combined effects of climate change and other local human stressors on the marine environment. Sci. Total Environ. 2021, 755, 142564. [Google Scholar] [CrossRef] [PubMed]
- Mannino, A.M.; Armeli Minicante, S.; Rodríguez-Prieto, C. Phycological Herbaria as a useful tool to monitor long-term changes of macroalgae diversity: Some case studies from the Mediterranean Sea. Diversity 2020, 12, 309. [Google Scholar] [CrossRef]
- Lang, P.L.; Willems, F.M.; Scheepens, J.F.; Burbano, H.A.; Bossdorf, O. Using herbaria to study global environmental change. New Phytol. 2019, 221, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Meineke, E.K.; Davis, C.C.; Davies, T.J. The unrealized potential of herbaria for global change biology. Ecol. Monogr. 2018, 88, 505–525. [Google Scholar] [CrossRef]
- Meineke, E.K.; Davies, T.J.; Daru, B.H.; Davis, C.C. Biological collections for understanding biodiversity in the Anthropocene. Philos. Trans. R. Soc. B 2019, 374, 20170386. [Google Scholar] [CrossRef]
- Lavoie, C. Biological collections in an ever changing world: Herbaria as tools for biogeographical and environmental studies. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 68–76. [Google Scholar] [CrossRef]
- Nelson, W.A.; Dalen, J.; Neill, K.F. Insights from natural history collections: Analysing the New Zealand macroalgal flora using herbarium data. PhytoKeys 2013, 30, 1. [Google Scholar] [CrossRef]
- Heberling, J.M.; Isaac, B.L. Herbarium specimens as exaptations: New uses for old collections. Am. J. Bot. 2017, 104, 963–965. [Google Scholar] [CrossRef]
- Alfonso, B.; Sansón, M.; Sangil, C.; Expósito, F.J.; Díaz, J.P.; Hernández, J.C. Herbarium macroalgae specimens reveal a rapid reduction of thallus size and reproductive effort related with climate change. Mar. Environ. Res. 2022, 174, 105546. [Google Scholar] [CrossRef]
- Wilkinson, T.; Wiken, E.; Bezaury, C.J.; Hourigan, T.; Agardy, H.; Herrmann, L.; Janishevski, C.; Madden, L.; Morgan Padilla, M. Ecorregiones Marinas de América del Norte; Comisión para la Cooperación Ambiental: Montreal, QC, Canada, 2009; 200p. [Google Scholar]
- Chiappa-Carrara, X.; Enríquez, C.; Papiol, V.; Mariño-Tapia, I.; Reyes-Hernández, C. Pacific coast of Mexico. In World Seas: An Environmental Evaluation; Academic Press: Cambridge, MA, USA, 2019; pp. 655–671. [Google Scholar]
- Lara-Lara, J.R.; Arenas Fuentes, V.; Bazán Guzmán, C.; Díaz Castañeda, V.; Escobar Briones, E.; García Abad, M.C.; Gaxiola-Castro, G.; Robles-Jarero, G.; Sosa-Ávalos, R.; Soto-González, L.A.; et al. Los ecosistemas marinos. In Capital Natural de México, Vol. I: Conocimiento Actual de la Biodiversidad; Sarukhan, J., Ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, Mexico, 2008; pp. 135–159. [Google Scholar]
- Botello, A.V.; Páez-Osuna, F.; Mendez-Rodríguez, L.; Betancourt-Lozano, M.; Álvarez-Borrego, S.; Lara-Lara, R. Pacífico Mexicano Contaminación e Impacto Ambiental. Diagnóstico y Tendencias; UAC, UNAM-ICMYL, CIAD-MAZATLÁN, CIBNOR, CICESE: Ciudad de México, Mexico, 2014; p. 930. [Google Scholar]
- Deser, C.; Wallace, J.M. Large-scale atmospheric circulation features of warm and cold episodes in the tropical Pacific. J. Clim. 1990, 3, 1254–1281. [Google Scholar] [CrossRef]
- Cai, W.; Santoso, A.; Wang, G.; Yeh, S.W.; An, S.I.; Cobb, K.M.; Wu, L. ENSO and greenhouse warming. Nat. Clim. Change 2015, 5, 849–859. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. In Worldwide Electronic Publication; National University of Ireland: Galway, Ireland, 2025; Available online: http://www.algaebase.org (accessed on 1 March 2025).
- Dawson, E.Y. Marine red algae of Pacific Mexico. Part 1. Bangiales to Corallinaceae subf. Corallinoidae. Allan Hancock Pac. Exped. 1953, 17, 1–33. [Google Scholar]
- Norris, J.N. Marine Algae of the Northern Gulf of California II: Rhodophyta; Smithsonian Contributions to Botany; Smithsonian Institution Scholary Press: Washington, DC, USA, 2014; Volume 96, 555p. [Google Scholar]
- Mateo-Cid, L.E.; Mendoza-González, A.C. Algas marinas bentónicas de la costa noroccidental de Guerrero, México. Rev. Mex. Biodivers. 2012, 83, 905–928. [Google Scholar] [CrossRef]
- Mateo-Cid, L.E.; Mendoza-González, A.C.; Alvarado-Villanueva, R.; Ortega-Murillo, M.; Ceballos-Corona, J.G.A.; Sánchez-Heredia, J.D.; Méndez-Guzmán, I. Lista actualizada y nuevos registros de las algas rojas (Rhodophyta) del litoral de Michoacán, México. Rev. Mex. Biodivers. 2020, 91, e913353. [Google Scholar] [CrossRef]
- Abbott, I.A.; Hollenberg, G.J. Marine Algae of California; Standford University Press: Stanford, CA, USA, 1976. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden: Bronx, NY, USA, 2025; Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 1 March 2025).
- dos Santos, C.C.; de Castro Nunes, J.M.; do Nascimento Santos, G.; dos Santos Pestana, E.M.; Cassano, V.; de Mattos Lyra, G. Taxonomic changes in the Lomentariaceae (Rhodymeniales, Rhodophyta): Yendoa gen. nov. and Ceratodictyon sanctae-crucis sp. nov. Phycologia 2023, 62, 19–28. [Google Scholar] [CrossRef]
- Cerdeira-Estrada, S.; Martell-Dubois, R.; Valdez-Chavarin, J.; Rosique-de la Cruz, J.L.; Caballero-Aragón, H.; Santamaria-del-Angel, E.; López-Perea, J.; Perera-Valderrama, S.; Ressl, R. Marine-Coastal Information and Analysis System (SIMAR). 2014, CONABIO. Available online: https://simar.conabio.gob.mx (accessed on 23 July 2024).
- OMM. El Niño/Oscilación del Sur; No. 1145; Organización Meteorológica Mundial: Genève, Switzerland, 2014; 12p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 10 March 2025).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Alestra, T.; Schiel, D.R. Impacts of local and global stressors in intertidal habitats: Influence of altered nutrient, sediment and temperatura levels on the early life history of three habitat-forming macroalgae. J. Exp. Mar. Biol. Ecol. 2015, 468, 29–36. [Google Scholar] [CrossRef]
- Álvarez-Canali, D.; Sangil, C.; Reyes, J.; Sansón, M. Local variations in environmental parameters govern 50 years of the decline of Fucus guiryi populations in the Canary Islands (Eastern Atlantic). J. Sea Res. 2019, 155, 101823. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Zhang, L.; Pang, T.; Qin, R. Effects of temperature on the photosynthetic performance in mature thalli of the red alga Gelidium amansii (Gelidiaceae). Aquaculture 2019, 512, 734320. [Google Scholar] [CrossRef]
- Alfonso, B.; Sangil, C.; Sansón, M. Gelidiales (Rhodophyta) en las islas Canarias: Estudios previos y perspectivas futuras. Sci. Insul.-Isl. Sci. 2019, 2, 153–181. [Google Scholar]
- Alfonso, B.; Hernández, J.C.; Sangil, C.; Martín, L.; Expósito, F.J.; Díaz, J.P.; Sansón, M. Fast climatic changes place an endemic Canary Island macroalgae at extinction risk. Reg. Environ. Change 2021, 21, 113. [Google Scholar] [CrossRef]
- Hernández-Guerrero, C.J.; Casas-Valdez, M.; Ortega-García, S.; Hernández-Vazquez, S. Effect of climatic variation on the relative abundance of the red alga Gelidium robustum in Baja California Sur, Mexico. J. Appl. Phycol. 2000, 12, 177–183. [Google Scholar] [CrossRef]
- Sangil, C.; Sansón, M.; Afonso-Carrillo, J. Spatio-temporal variation and recruitment of Sargasssum flavifolium Kützing in sublittoral cobble bottoms: Relationships with environmental variables. J. Appl. Phycol. 2015, 27, 455–467. [Google Scholar] [CrossRef]
- Carballo, J.L.; Olabarria, C.; Garza, O.T. Analysis of four macroalgal assemblages along the Pacific Mexican coast during and after the 1997-98 el Niño. Ecosystems 2002, 5, 749–760. [Google Scholar] [CrossRef]
- Chávez-Sánchez, T.; Piñón-Gimate, A.; Casas-Valdez, M. Normal, the Blob, and El Niño conditions: Effects on macroalgal blooms in a subtropical zone of the Gulf of California. Estuar. Coast. Shelf Sci. 2022, 268, 107787. [Google Scholar]
- Marinovic, B.B.; Croll, D.A.; Gong, N.; Benson, S.R.; Chavez, F.P. Effects of the 1997–1999 El Niño and La Niña events on zooplankton abundance and euphausiid community composition within the Monterey Bay coastal upwelling system. Prog. Oceanogr. 2002, 54, 265–277. [Google Scholar] [CrossRef]
- Bennett, S.; Wernberg, T.; Arackal Joy, B. Central and rear-edge populations can be equally vulnerable to warming. Nat. Commun. 2015, 6, 10280. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).