
Unravelling Metazoan and Fish Community Patterns in Yujiang River, China: Insights from Beta Diversity Partitioning and Co-Occurrence Network



Abstract
1. Introduction
2. Materials and Methods
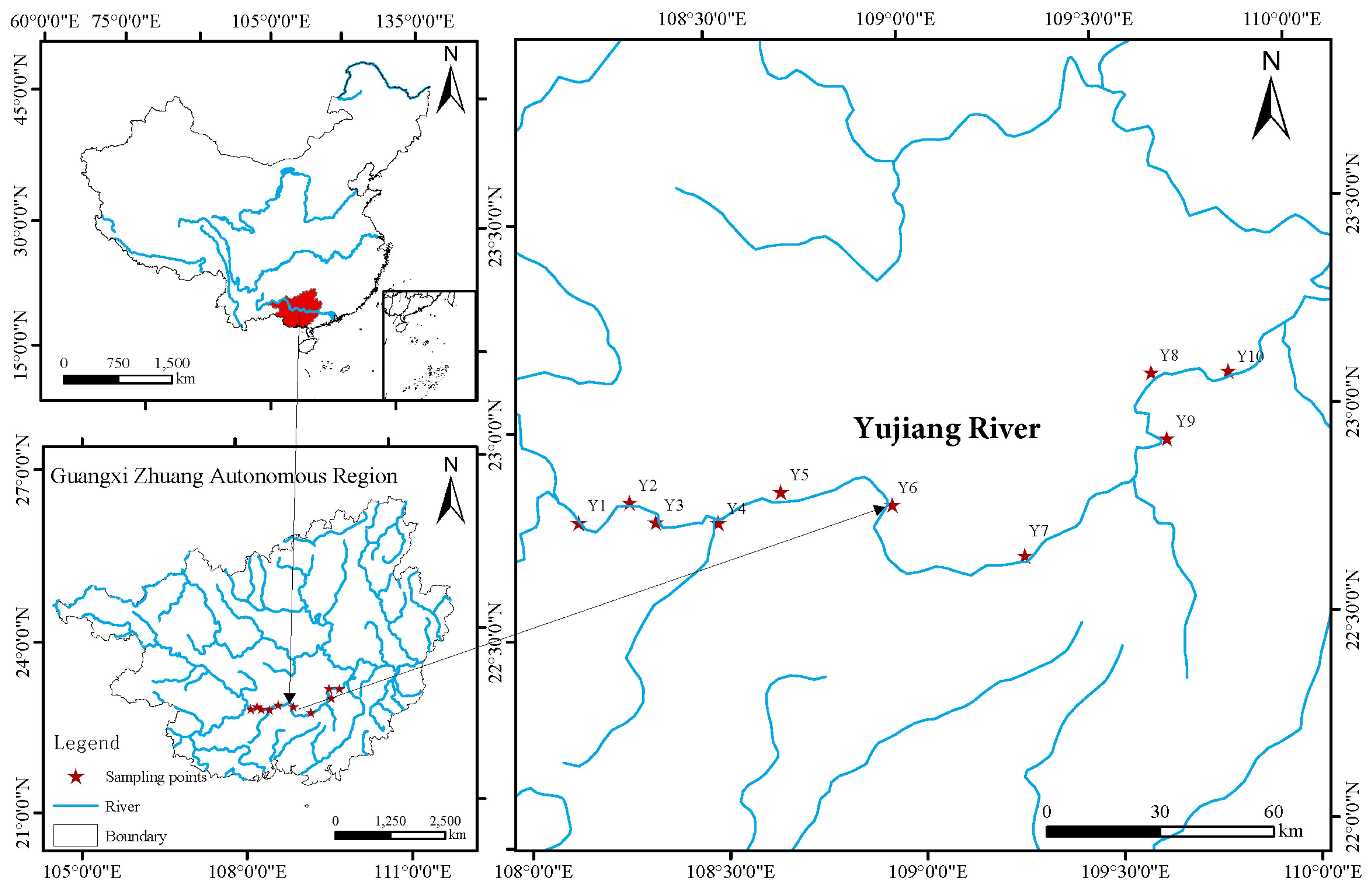
2.1. Study Area and Sample Collection
2.2. Measurements of Environmental Factors
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Data Processing
2.5. Statistical Analysis
3. Results
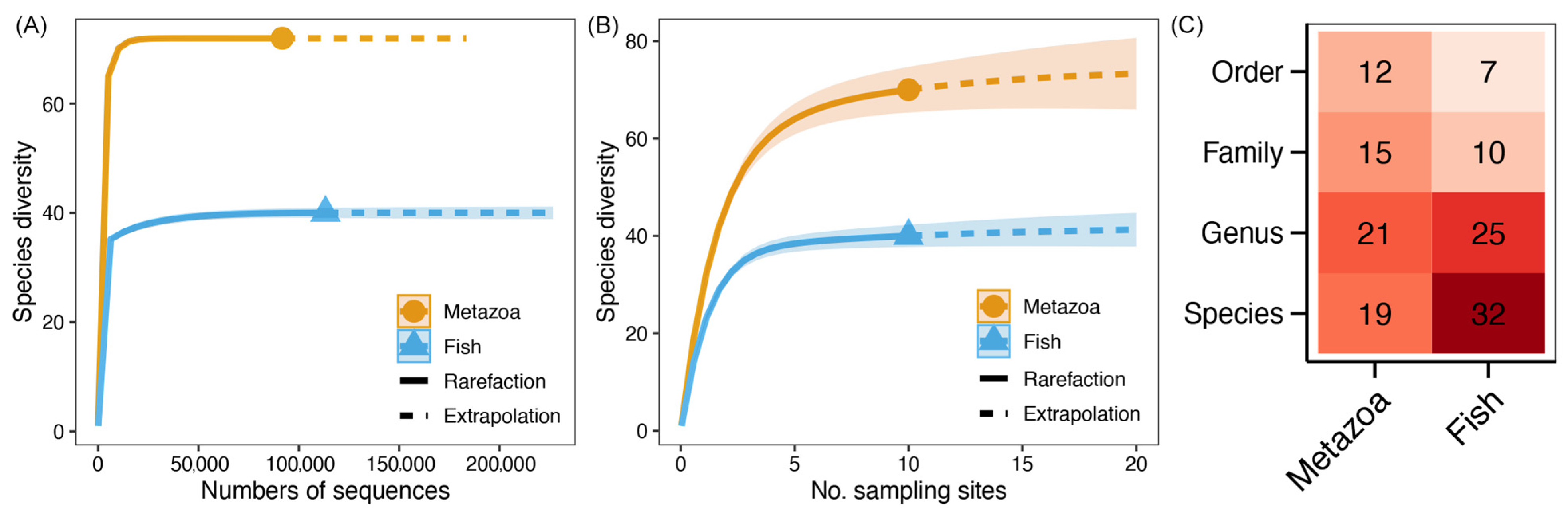
3.1. Biodiversity of Metazoan and Fish Communities
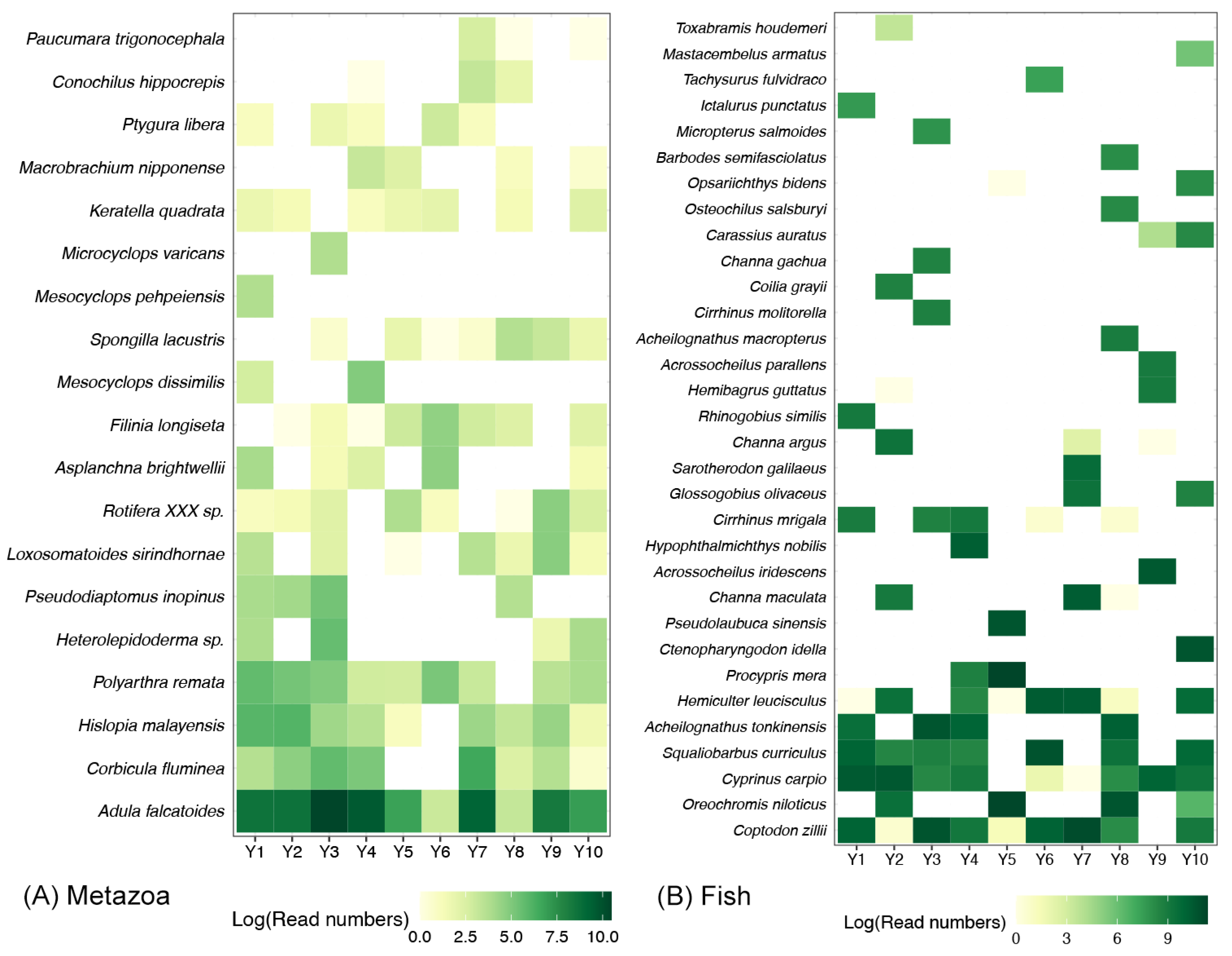
3.2. Compositions of Metazoan and Fish Communities
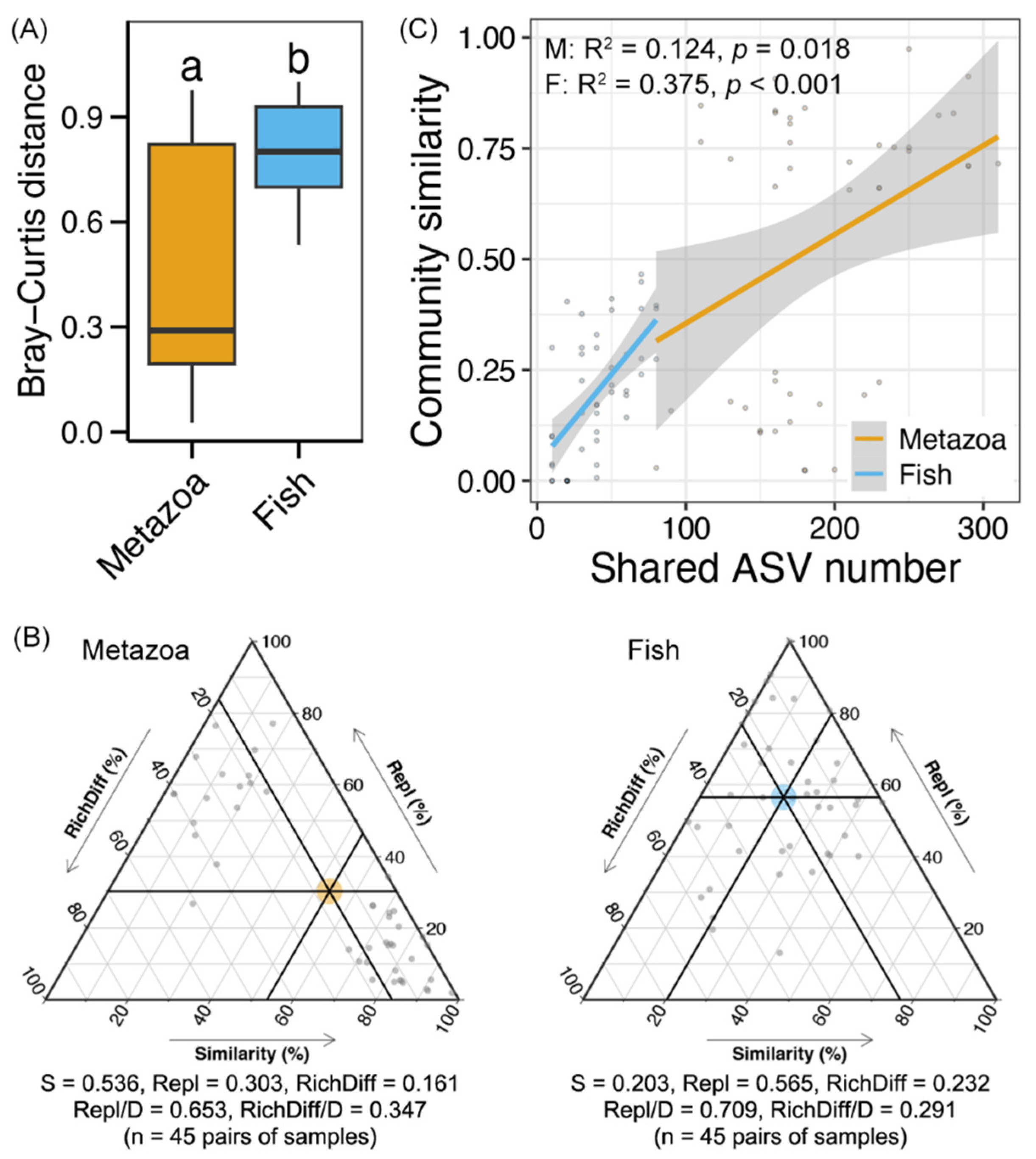
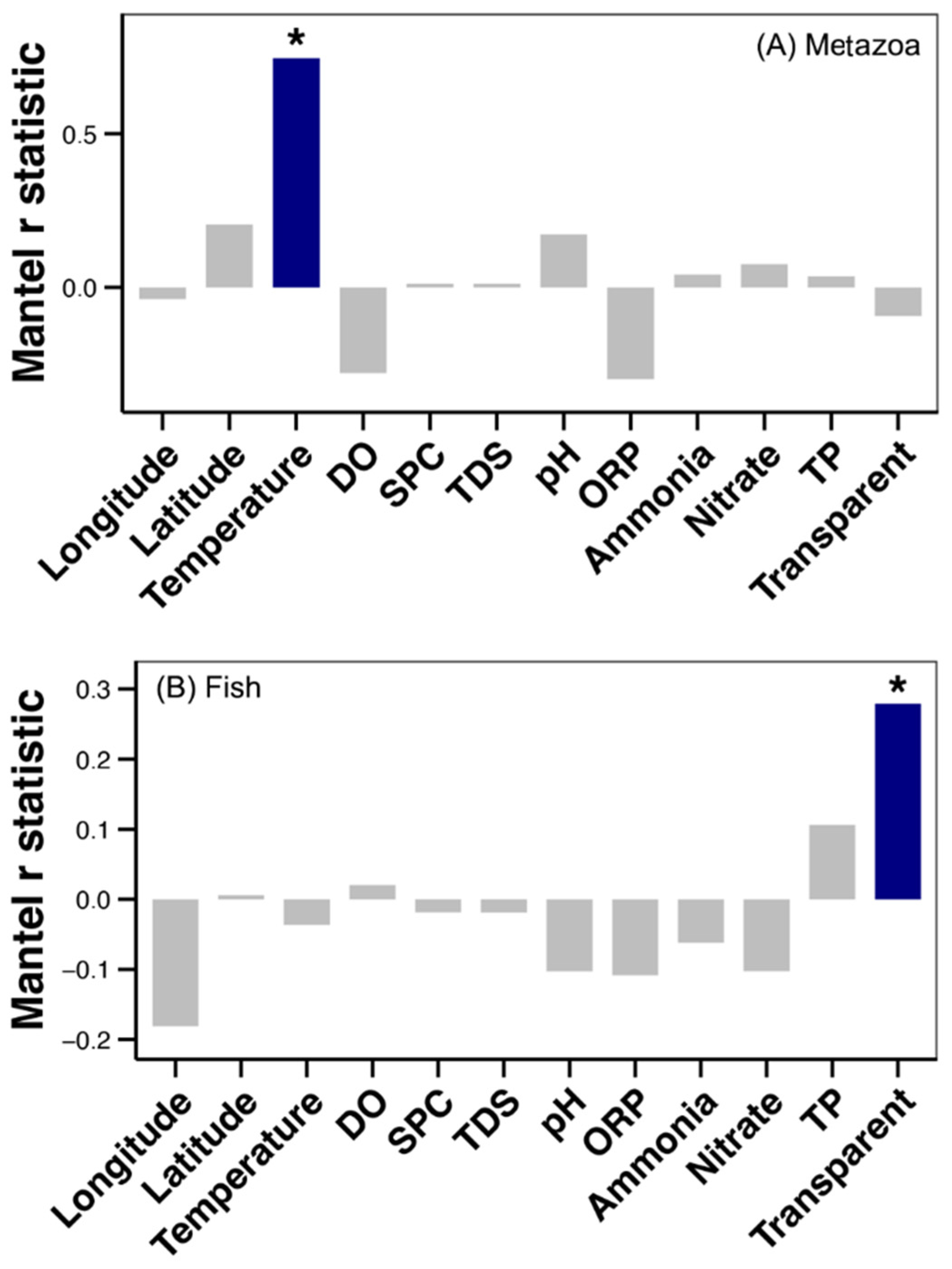
3.3. Beta Diversity Partitioning and Drivers of Metazoan and Fish Communities
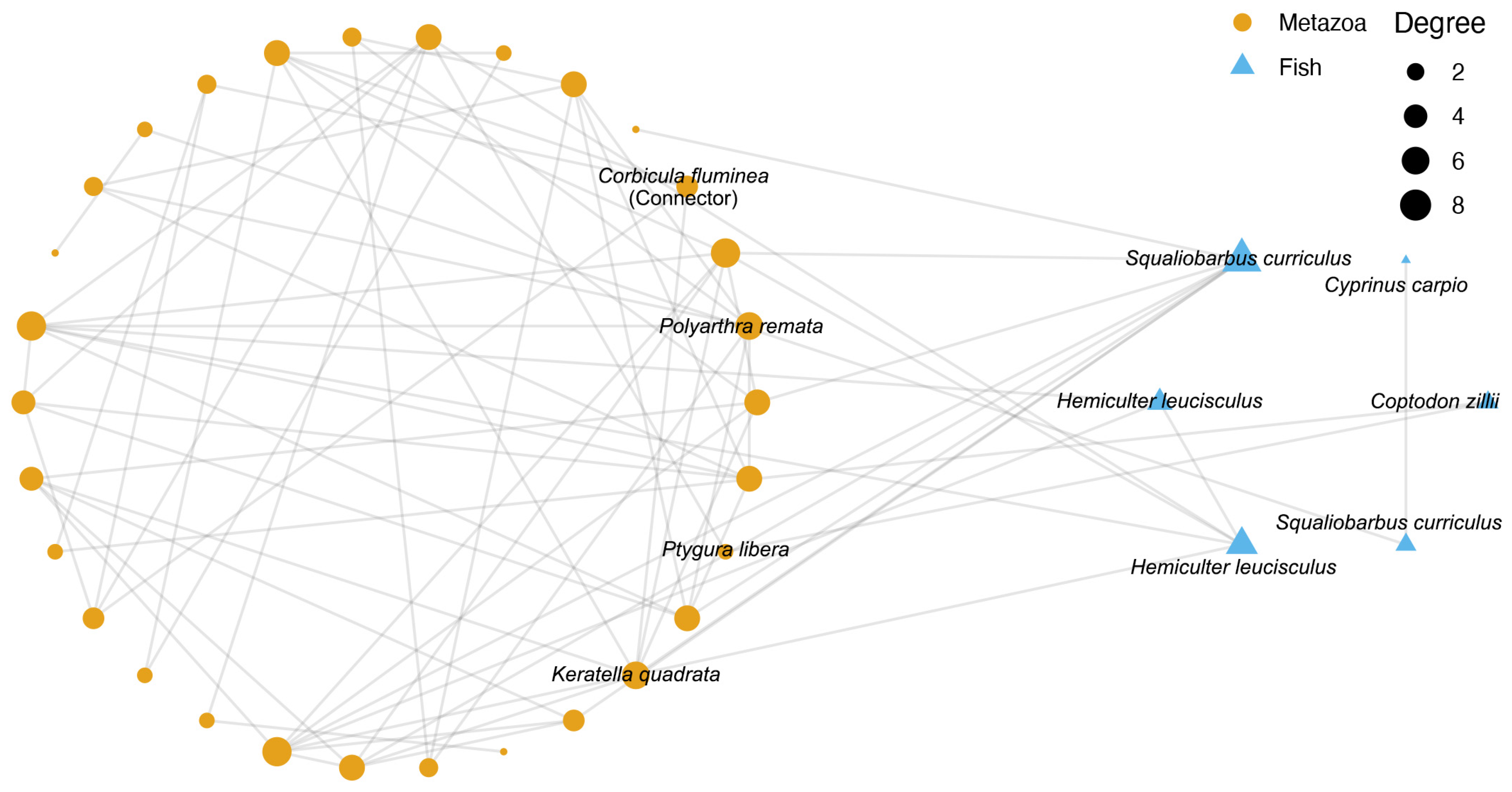
3.4. Co-Occurrence Network of Metazoa and Fish
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, P.; Li, D.; Sun, X.; Chu, Z.; Xia, T.; Zheng, B. Application of Ecological Restoration Technologies for the Improvement of Biodiversity and Ecosystem in the River. Water 2022, 14, 1402. [Google Scholar] [CrossRef]
- Blackman, R.C.; Ho, H.-C.; Walser, J.-C.; Altermatt, F. Spatio-Temporal Patterns of Multi-Trophic Biodiversity and Food-Web Characteristics Uncovered across a River Catchment Using Environmental DNA. Commun. Biol. 2022, 5, 259. [Google Scholar] [CrossRef] [PubMed]
- Pinna, M.; Zangaro, F.; Saccomanno, B.; Scalone, C.; Bozzeda, F.; Fanini, L.; Specchia, V. An Overview of Ecological Indicators of Fish to Evaluate the Anthropogenic Pressures in Aquatic Ecosystems: From Traditional to Innovative DNA-Based Approaches. Water 2023, 15, 949. [Google Scholar] [CrossRef]
- Tesfaye, G.C.; Souza, A.T.; Bartoň, D.; Blabolil, P.; Čech, M.; Draštík, V.; Frouzová, J.; Holubová, M.; Kočvara, L.; Kolařík, T.; et al. Long-Term Monitoring of Fish in a Freshwater Reservoir: Different Ways of Weighting Complex Spatial Samples. Front. Environ. Sci. 2022, 10, 1000087. [Google Scholar] [CrossRef]
- Fonseca, V.G.; Carvalho, G.R.; Sung, W.; Johnson, H.F.; Power, D.M.; Neill, S.P.; Packer, M.; Blaxter, M.L.; Lambshead, P.J.D.; Thomas, W.K.; et al. Second-Generation Environmental Sequencing Unmasks Marine Metazoan Biodiversity. Nat. Commun. 2010, 1, 98. [Google Scholar] [CrossRef] [PubMed]
- Talluto, L.; del Campo, R.; Estévez, E.; Altermatt, F.; Datry, T.; Singer, G. Towards (Better) Fluvial Meta-Ecosystem Ecology: A Research Perspective. NPJ Biodivers. 2024, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, Z.; Yang, G.; Feng, G. Dominated Taxonomic and Phylogenetic Turnover but Functional Nestedness of Wetland Bird Beta Diversity in North China. Land 2022, 11, 1090. [Google Scholar] [CrossRef]
- Kuehne, L.M.; Dickens, C.; Tickner, D.; Messager, M.L.; Olden, J.D.; O’Brien, G.; Lehner, B.; Eriyagama, N. The Future of Global River Health Monitoring. PLoS Water 2023, 2, e0000101. [Google Scholar] [CrossRef]
- Kaufmann, P.R.; Hughes, R.M.; Paulsen, S.G.; Peck, D.V.; Seeliger, C.W.; Weber, M.H.; Mitchell, R.M. Physical Habitat in Conterminous US Streams and Rivers, Part 1: Geoclimatic Controls and Anthropogenic Alteration. Ecol. Indic. 2022, 141, 109046. [Google Scholar] [CrossRef] [PubMed]
- Stenekes, S.; Parlee, B.; Seixas, C. Culturally Driven Monitoring: The Importance of Traditional Ecological Knowledge Indicators in Understanding Aquatic Ecosystem Change in the Northwest Territories’ Dehcho Region. Sustainability 2020, 12, 7923. [Google Scholar] [CrossRef]
- Fueyo, Á.; Sánchez, O.; Carleos, C.; Escudero, A.; Cordón, J.; Granero-Castro, J.; Borrell, Y.J. Unlocking Rivers’ Hidden Diversity and Ecological Status Using DNA Metabarcoding in Northwest Spain. Ecol. Evol. 2024, 14, e70110. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.N.; Shen, E.W.; Seemann, J.; Correa, A.M.S.; O’Donnell, J.L.; Altieri, A.H.; Knowlton, N.; Crandall, K.A.; Egan, S.P.; McMillan, W.O.; et al. Environmental DNA Survey Captures Patterns of Fish and Invertebrate Diversity across a Tropical Seascape. Sci. Rep. 2020, 10, 6729. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, D.-H.; Song, Y.-D.; Wang, T.-T.; Fan, S.-D.; Wu, E.-N.; Chen, N.-L.; Xia, W.-T.; Xu, M.N.; Chen, Z.-B.; et al. Application of Environmental DNA Metabarcoding to Identify Fish Community Characteristics in Subtropical River Systems. Ecol. Evol. 2024, 14, e11214. [Google Scholar] [CrossRef] [PubMed]
- Weigand, A.M.; Macher, J.-N. A DNA Metabarcoding Protocol for Hyporheic Freshwater Meiofauna: Evaluating Highly Degenerate COI Primers and Replication Strategy. Metabarcoding Metagenomics 2018, 2, e26869. [Google Scholar] [CrossRef]
- Lim, N.K.M.; Tay, Y.C.; Srivathsan, A.; Tan, J.W.T.; Kwik, J.T.B.; Baloğlu, B.; Meier, R.; Yeo, D.C.J. Next-Generation Freshwater Bioassessment: EDNA Metabarcoding with a Conserved Metazoan Primer Reveals Species-Rich and Reservoir-Specific Communities. R. Soc. Open Sci. 2016, 3, 160635. [Google Scholar] [CrossRef] [PubMed]
- Buchner, D.; Macher, T.-H.; Beermann, A.J.; Werner, M.-T.; Leese, F. Standardized High-Throughput Biomonitoring Using DNA Metabarcoding: Strategies for the Adoption of Automated Liquid Handlers. Environ. Sci. Ecotechnol. 2021, 8, 100122. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, N.P.; Delahooke, K.M.; Barnes, N.; Rideout, B.W.T.; Kenchington, C.G.; Manica, A.; Mitchell, E.G. Morphology Shapes Community Dynamics in Early Animal Ecosystems. Nat. Ecol. Evol. 2024, 8, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Villéger, S.; Miranda, J.R.; Hernandez, D.F.; Mouillot, D. Low Functional β-Diversity despite High Taxonomic β-Diversity among Tropical Estuarine Fish Communities. PLoS ONE 2012, 7, e40679. [Google Scholar] [CrossRef] [PubMed]
- Lamy, T.; Legendre, P.; Chancerelle, Y.; Siu, G.; Claudet, J. Understanding the Spatio-Temporal Response of Coral Reef Fish Communities to Natural Disturbances: Insights from Beta-Diversity Decomposition. PLoS ONE 2015, 10, e0138696. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Fan, C.; Zhang, C.; Zhao, X.; Gadow, K. von Decomposing Spatial β-Diversity in the Temperate Forests of Northeastern China. Ecol. Evol. 2021, 11, 11362–11372. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Xu, D.; Ren, L.; Zhao, D. Using a Topological Approach to Identify Keystone Species of Fish in Eutrophic Lake Ecosystems: A Case of Zhushan Bay, Taihu Lake. Fish. Manag. Ecol. 2023, 30, 89–98. [Google Scholar] [CrossRef]
- GB 11893-89; Water Quality—Determination of Total Phosphorus—Ammonium Molybdate Spectrophotometric Method. State Bureau of Technical Supervision: Beijing, China, 1989.
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.M.; Breiner, H.-W.; Richards, T.A. Multiple Marker Parallel Tag Environmental DNA Sequencing Reveals a Highly Complex Eukaryotic Community in Marine Anoxic Water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a Set of Universal PCR Primers for Metabarcoding Environmental DNA from Fishes: Detection of More than 230 Subtropical Marine Species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, J.; Zhao, Y.; He, Y.; Chen, J.; Dong, C.; Liu, L.; Wang, J.; Zhou, L. Contrasting Pollution Responses of Native and Non-Native Fish Communities in Anthropogenically Disturbed Estuaries Unveiled by eDNA Metabarcoding. J. Hazard. Mater. 2024, 480, 136323. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Wang, R.; Xu, S.; Li, Z.; Liu, L.; Li, M.; Zhou, L. Changes in Protist Communities in Drainages across the Pearl River Delta under Anthropogenic Influence. Water Res. 2021, 200, 117294. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2’s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference Database (PR2): A Catalog of Unicellular Eukaryote Small Sub-Unit rRNA Sequences with Curated Taxonomy. Nucleic Acids Res. 2013, 41, D597–D604. [Google Scholar] [CrossRef] [PubMed]
- Leray, M.; Knowlton, N.; Machida, R.J. MIDORI2: A Collection of Quality Controlled, Preformatted, and Regularly Updated Reference Databases for Taxonomic Assignment of Eukaryotic Mitochondrial Sequences. Environ. DNA 2022, 4, 894–907. [Google Scholar] [CrossRef]
- Wan, W.; Grossart, H.; He, D.; Liu, W.; Wang, S.; Yang, Y. Differentiation Strategies for Planktonic Bacteria and Eukaryotes in Response to Aggravated Algal Blooms in Urban Lakes. Imeta 2023, 2, e84. [Google Scholar] [CrossRef] [PubMed]
- Steinhauser, D.; Krall, L.; Müssig, C.; Büssis, D.; Usadel, B. Correlation Networks. In Analysis of Biological Networks; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008; pp. 305–333. ISBN 978-0-470-25348-9. [Google Scholar]
- Zhao, Z.; Li, H.; Sun, Y.; Zhan, A.; Lan, W.; Woo, S.P.; Shau-Hwai, A.T.; Fan, J. Bacteria versus Fungi for Predicting Anthropogenic Pollution in Subtropical Coastal Sediments: Assembly Process and Environmental Response. Ecol. Indic. 2022, 134, 108484. [Google Scholar] [CrossRef]
- Chen, W.; Liu, L.; Wang, J.; Zhou, L. Threatened Freshwater Fish Need Protection. Science 2021, 374, 164. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, P.; Xu, C.; Sun, Y.; Shi, L.; Zhou, L.; Jeppesen, E.; Chen, J.; Xie, P. Can the “10-Year Fishing Ban” Rescue Biodiversity of the Yangtze River? Innovation 2022, 3, 100235. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, J.; Wang, P.; Jeppesen, E.; Xie, P. How to Manage Fish within and after the 10-Year Fishing Ban. Innovation 2024, 5, 100694. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Jin, Z.; Yuan, Z.; Liu, Q.; Bowler, P.A. Biodiversity and Conservation of Freshwater Fishes in the Yujiang River, China. Pak. J. Zool. 2024, 56, 2501–3000. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, X.; Ao, X.; Qin, J.; Wu, X.; Ouyang, S. Fish Diversity in the Middle and Lower Reaches of the Ganjiang River of China: Threats and Conservation. PLoS ONE 2018, 13, e0205116. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, Y.; He, J.; Hou, Y.; Wang, F. The Native Fish Diversity with Environmental Influencing Factors in the Daqing River Basin, China. Front. Environ. Sci. 2024, 12, 1450903. [Google Scholar] [CrossRef]
- Wang, C.; Shao, J.; Ma, B.; Xie, J.; Li, D.; Liu, X.; Huo, B. Longitudinal Patterns in Fish Assemblages after Long-Term Ecological Rehabilitation in the Taizi River, Northeastern China. Sustainability 2022, 14, 14973. [Google Scholar] [CrossRef]
- Li, S.; Huang, Y.; Li, F.; Liu, Y.; Ma, H.; Zhang, X.; Wang, X.; Chen, W.; Cui, G.; Wang, T. Functional Alpha and Beta Diversity of Fish Communities and Their Relationship with Environmental Factors in the Huanghe River (Yellow River) Estuary and Adjacent Seas, China. Fishes 2024, 9, 222. [Google Scholar] [CrossRef]
- Zhou, L.; Guo, D.; Zeng, L.; Xu, P.; Tang, Q.; Chen, Z.; Zhu, Q.; Wang, G.; Chen, Q.; Chen, L.; et al. The Structuring Role of Artificial Structure on Fish Assemblages in a Dammed River of the Pearl River in China. Aquat. Living Resour. 2018, 31, 15. [Google Scholar] [CrossRef]
- Guo, D.; Zhou, L.; Wang, G.; Lai, H.; Bi, S.; Chen, X.; Zhao, X.; Liu, S.; Luo, Y.; Li, G. Use of Artificial Structures to Enhance Fish Diversity in the Youjiang River, a Dammed River of the Pearl River in China. Ecol. Evol. 2020, 10, 13439–13450. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.S.; Batool, A.I.; Rehman, M.F.U.; Naz, S. Comparative Analysis of the Haemato-Biochemical Parameters and Growth Characteristics of Oreochromis Niloticus (Nile tilapia) Cultured under Different Feed and Habitats (Biofloc Technology and Earthen Pond System). Aquac. Res. 2022, 53, 6184–6192. [Google Scholar] [CrossRef]
- Geletu, T.T.; Tang, S.; Xing, Y.; Zhao, J. Ecological Niche and Life-History Traits of Redbelly Tilapia (Coptodon Zillii, Gervais 1848) in Its Native and Introduced Ranges. Aquat. Living Resour. 2024, 37, 2. [Google Scholar] [CrossRef]
- Radkhah, A.R.; Eagderi, S. Ecological Consequences of Tilapia Species on Fish Biodiversity of Iran and Challenges Arising from Their Introduction. Iran. J. Ichthyol. 2021, 8, 342–350. [Google Scholar] [CrossRef]
- Feng, S.; Pan, X.; Wang, J.; Liu, W.; Hui, Y.; Wang, G.; Liu, K.; Li, J.; Xu, H.; Lin, L.; et al. Risk Screening of the Non-Native Fish in the Jiulong River Basin of Southeast China. Animals 2025, 15, 461. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Xing, Y.; Geletu, T.T.; Zhao, J. Trophic Plasticity of the Invasive Redbelly Tilapia (Coptodon zillii) in China Inferred from DNA Metabarcoding Analysis. Ecol. Evol. 2025, 15, e71118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mei, X.; Tang, Y.; Razlutskij, V.; Peterka, J.; Taylor, W.D.; Naselli-Flores, L.; Liu, Z.; Tong, C.; Zhang, X. Effects of Nile Tilapia (Oreochromis niloticus) on Phytoplankton Community Structure and Water Quality: A Short-Term Mesocosm Study. Knowl. Manag. Aquat. Ecosyst. 2022, 423, 11. [Google Scholar] [CrossRef]
- Chaianunporn, T.; Panthum, T.; Singchat, W.; Chaianunporn, K.; Suksavate, W.; Chaiyes, A.; Muangmai, N.; Marod, D.; Duengkae, P.; Srikulnath, K. Sustainable Ecosystem Management Strategies for Tackling the Invasion of Blackchin Tilapia (Sarotherodon melanotheron) in Thailand: Guidelines and Considerations. Animals 2024, 14, 3292. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Zhang, S.; Xu, S.; Xiong, P.; Cao, Y.; Chen, Z.; Li, M. Comparison of Environmental DNA Metabarcoding and Bottom Trawling for Detecting Seasonal Fish Communities and Habitat Preference in a Highly Disturbed Estuary. Ecol. Indic. 2023, 146, 109754. [Google Scholar] [CrossRef]
- Erős, T.; Funk, A.; Pont, D.; Hein, T.; Meulenbroek, P.; Preiszner, B.; Valentini, A.; Czeglédi, I. eDNA Metabarcoding Reveals the Role of Habitat Specialization and Spatial and Environmental Variability in Shaping Diversity Patterns of Fish Metacommunities. PLoS ONE 2024, 19, e0296310. [Google Scholar] [CrossRef] [PubMed]
- Victorero, L.; Robert, K.; Robinson, L.F.; Taylor, M.L.; Huvenne, V.A.I. Species Replacement Dominates Megabenthos Beta Diversity in a Remote Seamount Setting. Sci. Rep. 2018, 8, 4152. [Google Scholar] [CrossRef] [PubMed]
- Ohira, M.; Fukuda, S. Exploring Functional Flow Heterogeneity in Regulated Flow Regime: Fish Species Turnover along Hydraulic Gradients in an Artificial Waterway Network. River Res. Appl. 2023, 39, 1961–1971. [Google Scholar] [CrossRef]
- Borges, S.H.; D’Aquino, D.D.; da Cruz, M.V.; de Souza, R.F. Turnover in Fish Species Composition Is Related to Water Colour of Amazonian Rivers. J. Trop. Ecol. 2023, 39, e7. [Google Scholar] [CrossRef]
- Rutt, C.L.; Cooper, W.J.; Andretti, C.B.; Costa, T.V.V.; Stouffer, P.C.; Vargas, C.F.; Luther, D.A.; Cohn-Haft, M. Low Species Turnover of Upland Amazonian Birds in the Absence of Physical Barriers. Divers. Distrib. 2023, 29, 466–477. [Google Scholar] [CrossRef]
- Hong, Z.; Chen, X.; Hu, J.; Chang, X.; Qian, Y. Adverse Effects of Microcystis Aeruginosa Exudates on the Filtration, Digestion, and Reproduction Organs of Benthic Bivalve Corbicula fluminea. Sci. Rep. 2024, 14, 10934. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Damborenea, C.; Penchaszadeh, P.E.; Darrigran, G. Gonadal Cycle of Corbicula fluminea (Bivalvia: Corbiculidae) in Pampean Streams (Southern Neotropical Region). PLoS ONE 2017, 12, e0186850. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feng, C.; Pang, W.; Tian, C.; Zhao, Y. Nanoplastic-Induced Genotoxicity and Intestinal Damage in Freshwater Benthic Clams (Corbicula fluminea): Comparison with Microplastics. ACS Nano 2021, 15, 9469–9481. [Google Scholar] [CrossRef] [PubMed]
- Yuan, N.; Ding, J.; Wu, J.; Bao, E.; Chu, Y.; Hu, F. A Multibiomarker Approach to Assess the Ecotoxicological Effects of Diclofenac on Asian Clam Corbicula fluminea (O. F. Müller, 1774). Environ. Sci. Pollut. Res. 2023, 30, 88598–88611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yin, J.; Tang, S.; Li, D.; Gu, X.; Zhang, S.; Suo, W.; Liu, X.; Liu, Y.; Jiang, Q.; et al. Dissecting the Chromosome-Level Genome of the Asian Clam (Corbicula fluminea). Sci. Rep. 2021, 11, 15021. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Index | Metazoa | Fish |
|---|---|---|
| Chao1 | 35.64 ± 8.72 | 9.80 ± 2.49 |
| Pd faith | 4.21 ± 1.13 | 2.44 ± 0.51 |
| Shannon | 1.28 ± 0.86 | 1.56 ± 0.22 |
| Pielou J | 0.38 ± 0.26 | 0.70 ± 0.05 |
| Chao (gamma) | 89.64 ± 12.99 | 133.75 ± 66.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, D.; Huang, Y.; Shi, J.; Wu, W.; Yuan, C.; Nong, S.; Guo, C.; Chen, W.; Zhou, L. Unravelling Metazoan and Fish Community Patterns in Yujiang River, China: Insights from Beta Diversity Partitioning and Co-Occurrence Network. Diversity 2025, 17, 488. https://doi.org/10.3390/d17070488
Li Y, Wang D, Huang Y, Shi J, Wu W, Yuan C, Nong S, Guo C, Chen W, Zhou L. Unravelling Metazoan and Fish Community Patterns in Yujiang River, China: Insights from Beta Diversity Partitioning and Co-Occurrence Network. Diversity. 2025; 17(7):488. https://doi.org/10.3390/d17070488
Chicago/Turabian StyleLi, Yusen, Dapeng Wang, Yuying Huang, Jun Shi, Weijun Wu, Chang Yuan, Shiqiong Nong, Chuanbo Guo, Wenjian Chen, and Lei Zhou. 2025. "Unravelling Metazoan and Fish Community Patterns in Yujiang River, China: Insights from Beta Diversity Partitioning and Co-Occurrence Network" Diversity 17, no. 7: 488. https://doi.org/10.3390/d17070488
APA StyleLi, Y., Wang, D., Huang, Y., Shi, J., Wu, W., Yuan, C., Nong, S., Guo, C., Chen, W., & Zhou, L. (2025). Unravelling Metazoan and Fish Community Patterns in Yujiang River, China: Insights from Beta Diversity Partitioning and Co-Occurrence Network. Diversity, 17(7), 488. https://doi.org/10.3390/d17070488