
Preliminary Insights into Summer Archaeal Communities in Eutrophic Jinhae Bay, Korea


Abstract
1. Introduction
2. Methods
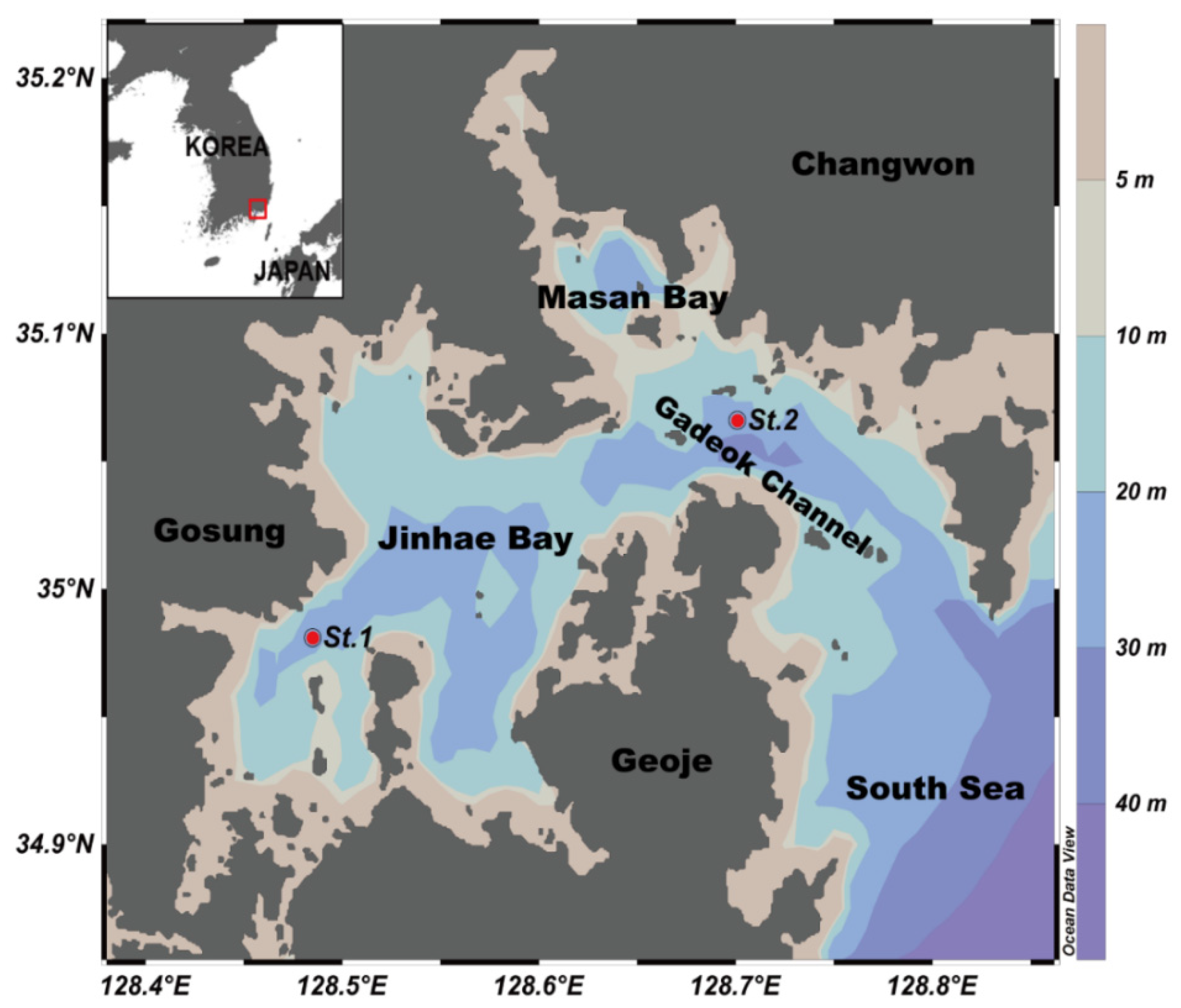
2.1. Seawater Sampling
2.2. DNA Analysis
2.3. Data Visualization and Availability
3. Results and Discussion
3.1. Physical Features of Water Columns
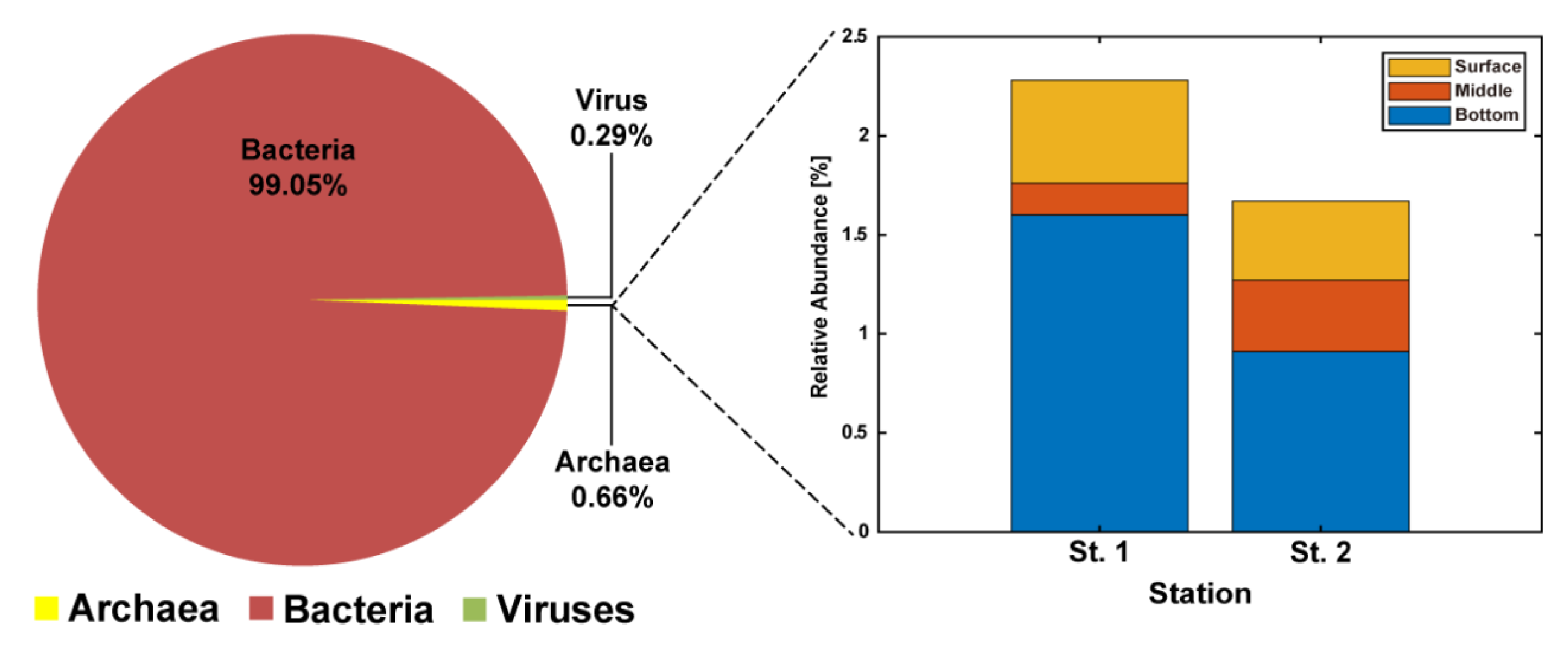
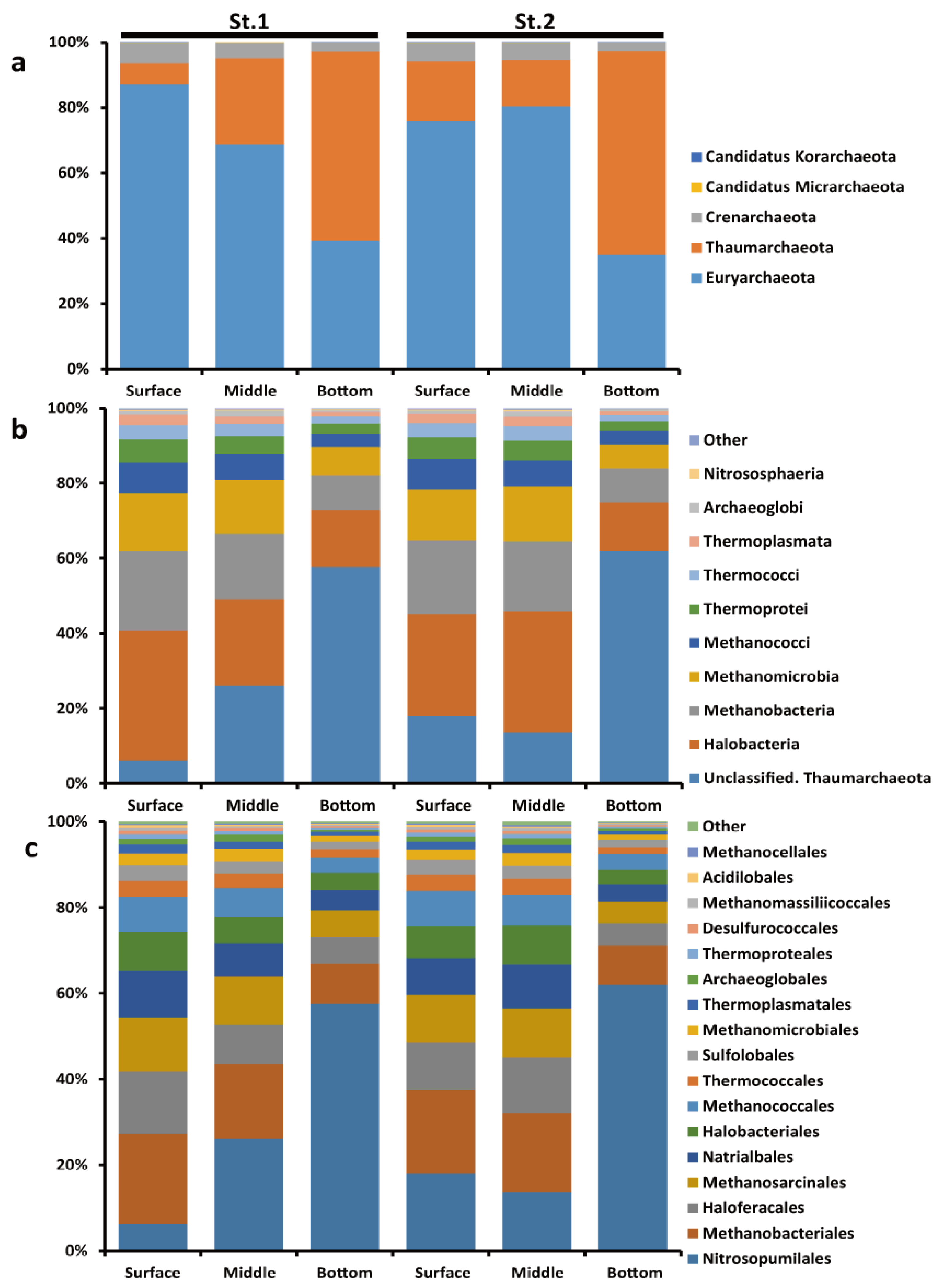
3.2. Community Structures of Archaea in Jinhae Bay and Comparison with Other Hypoxia Regions
3.3. Inferring Potential Biogeochemical Functions
4. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a Natural System of Organisms: Proposal for the Domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, L.J.; Mancinelli, R.L. Life in Extreme Environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Karner, M.B.; DeLong, E.F.; Karl, D.M. Archaeal Dominance in the Mesopelagic Zone of the Pacific Ocean. Nature 2001, 409, 507–510. [Google Scholar] [CrossRef]
- Magregor, B.J.; Moser, D.P.; Alm, E.W.; Nealson, K.H.; Stahl, D.A. Crenarchaeota in Lake Michigan Sediment. Appl. Environ. Microbiol. 1997, 63, 1178–1181. [Google Scholar] [CrossRef]
- King, G.; Smith, C.; Tolar, B.; Hollibaugh, J. Analysis of Composition and Structure of Coastal to Mesopelagic Bacterioplankton Communities in the Northern Gulf of Mexico. Front. Microbiol. 2013, 3, 438. [Google Scholar] [CrossRef]
- Gillies, L.E.; Cameron, T.J.; Sergio, D.; Rabalais, N.N.; Mason, O.U. Archaeal Enrichment in the Hypoxic Zone in the Northern Gulf of Mexico. Environ. Microbiol. 2015, 17, 3847–3856. [Google Scholar] [CrossRef]
- Caffrey, J.M.; Bano, N.; Kalanetra, K.; Hollibaugh, J.T. Ammonia Oxidation and Ammonia-Oxidizing Bacteria and Archaea from Estuaries with Differing Histories of Hypoxia. ISME J. 2007, 1, 660–662. [Google Scholar] [CrossRef]
- Lee, J.; Lim, J.; Park, J.; Youn, S.; Oh, H.; Kim, J.; Kim, M.K.; Cho, H.; Yoon, J.; Kim, S.; et al. First Investigation of Microbial Community Composition in the Bridge (Gadeok Channel) between the Jinhae-Masan Bay and the South Sea of Korea. Ocean Sci. J. 2018, 53, 251–260. [Google Scholar] [CrossRef]
- Kwon, J.; Lee, J.; Kim, Y.; Lim, J.; Choi, T.; Ye, M.; Jun, J.; Kim, S. Long-Term Variations of Water Quality in Jinhae Bay. J. Korean Soc. Mar. Environ. Energy 2014, 17, 324–332. [Google Scholar] [CrossRef]
- Kim, N.S.; Kang, H.; Kwon, M.; Jang, H.; Kim, J.G. Comparison of Seawater Exchange Rate of Small Scale Inner Bays within Jinhae Bay. J. Korean Soc. Mar. Environ. Energy. 2016, 19, 74–85. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, M.; Yoon, J. Study of Formation and Development of Oxygen Deficient Water Mass, using Ecosystem Model in Jinhae, Masan Bay. J. Ocean. Eng. Technol. 2010, 24, 41–50. [Google Scholar]
- Lee, J.; Lim, J.; Park, J.; Kim, I. Temporal and Vertical Variation in Microbial Community Composition in Response to Physicochemical Characteristics in a Water Column of Highly Eutrophied Jinhae Bay, South Korea. J. Mol. Microbiol. Biotechnol. 2018, 28, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Kim, J.I.; Kim, S.T.; Lee, J.H.; Lee, J.B. Seasonal Variation in Species Composition of Catch by a Coastal Beam Trawl in Jinhae Bay and Jinju Bay, Korea. J. Korean Soc. Fish. Tech. 2012, 48, 428–444. [Google Scholar] [CrossRef]
- Hwang, O.; Shin, K.; Baek, S.; Lee, W.; Kim, S.; Jang, M. Annual Variations in Community Structure of Mesozooplankton by Short-Term Sampling in Jangmok Harbor of Jinhae Bay. Ocean Polar Res. 2011, 33, 235–253. [Google Scholar] [CrossRef]
- Lim, K.; Shin, H.; Yoon, S.; Koh, C. Assessment of Benthic Environment Based on Macrobenthic Community Analysis in Jinhae Bay, Korea. Sea 2007, 12, 9–23. [Google Scholar]
- Lee, J.; Park, K.; Lim, J.; Yoon, J.; Kim, I. Hypoxia in Korean Coastal Waters: A Case Study of the Natural Jinhae Bay and Artificial Shihwa Bay. Front. Mar. Sci. 2018, 5, 70. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kim, D.; Song, L.; Breitwieser, F.P.; Salzberg, S.L. Centrifuge: Rapid and Sensitive Classification of Metagenomic Sequences. Genome Res. 2016, 26, 1721–1729. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive Metagenomic Visualization in a Web Browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Bernhard, A.E.; Landry, Z.C.; Blevins, A.; de la Torre, J.R.; Giblin, A.E.; Stahl, D.A. Abundance of Ammonia-Oxidizing Archaea and Bacteria Along an Estuarine Salinity Gradient in Relation to Potential Nitrification Rates. J. Appl. Environ. Microbiol. 2010, 76, 1285–1289. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S.; He, J.Z. Nitrification Driven by Bacteria and Not Archaea in Nitrogen-Rich Grassland Soils. Nat. Geosci. 2009, 2, 621–624. [Google Scholar] [CrossRef]
- Schleper, C. Ammonia Oxidation: Different Niches for Bacteria and Archaea? ISME J. 2010, 4, 1092–1094. [Google Scholar] [CrossRef]
- Yool, A.; Martin, A.P.; Fernández, C.; Clark, D.R. The Significance of Nitrification for Oceanic New Production. Nature 2007, 447, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Newell, S.E.; Babbin, A.R.; Jayakumar, A.; Ward, B.B. Ammonia Oxidation Rates and Nitrification in the Arabian Sea. Glob. Biogeochem. Cycles 2011, 25, GB4016. [Google Scholar] [CrossRef]
- Santoro, A.E.; Buchwald, C.; McIlvin, M.R.; Casciotti, K.L. Isotopic Signature of N2O Produced by Marine Ammonia-Oxidizing Archaea. Science 2011, 333, 1282–1285. [Google Scholar] [CrossRef]
- Mosier, A.C.; Allen, E.E.; Kim, M.; Ferriera, S.; Francis, C.A. Genome Sequence of “Candidatus Nitrosopumilus Salaria” BD31, an Ammonia-Oxidizing Archaeon from the San Francisco Bay Estuary. J. Bacteriol. 2012, 194, 2121–2122. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.; Jung, M.; Kim, S.; Cha, I.; Ghai, R.; MartÃn-Cuadrado, A.; RodrÃguez-Valera, F.; Rhee, S. Draft Genome Sequence of an Ammonia-Oxidizing Archaeon, “Candidatus Nitrosopumilus Sediminis” AR2, from Svalbard in the Arctic Circle. J. Bacteriol. 2012, 194, 6948–6949. [Google Scholar] [CrossRef] [PubMed]
- Stahl, D.A.; de la Torre, J.R. Physiology and Diversity of Ammonia-Oxidizing Archaea. Annu. Rev. Microbiol. 2012, 66, 83–101. [Google Scholar] [CrossRef]
- Walker, C.B.; de la Torre, J.R.; Klotz, M.G.; Urakawa, H.; Pinel, N.; Arp, D.J.; Brochier-Armanet, C.; Chain, P.S.G.; Chan, P.P.; Gollabgir, A.; et al. Nitrosopumilus Maritimus Genome Reveals Unique Mechanisms for Nitrification and Autotrophy in Globally Distributed Marine Crenarchaea. Proc. Natl. Acad. Sci. USA 2010, 107, 8818–8823. [Google Scholar] [CrossRef]
- Könneke, M.; Bernhard, A.E.; de la Torre, J.R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an Autotrophic Ammonia-Oxidizing Marine Archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef]
- Offre, P.; Spang, A.; Schleper, C. Archaea in Biogeochemical Cycles. Annu. Rev. Microbiol. 2013, 67, 437–457. [Google Scholar] [CrossRef] [PubMed]
- Conrad, R. The Global Methane Cycle: Recent Advances in Understanding the Microbial Processes Involved. Environ. Microbiol. Rep. 2009, 1, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Knittel, K.; Boetius, A. Anaerobic Oxidation of Methane: Progress with an Unknown Process. Annu. Rev. Microbiol. 2009, 63, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Haroon, M.F.; Hu, S.; Shi, Y.; Imelfort, M.; Keller, J.; Hugenholtz, P.; Yuan, Z.; Tyson, G.W. Anaerobic Oxidation of Methane Coupled to Nitrate Reduction in a Novel Archaeal Lineage. Nature 2013, 500, 567–570. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Station | Depth (m) | T (°C) | S | DO (mg L−1) | Oxygen Conditions |
|---|---|---|---|---|---|
| 1 | 0 | 28.2 | 30.7 | 7.82 | Oxic |
| 10 | 22.3 | 32.1 | 7.61 | Oxic | |
| 21 | 17.7 | 33.3 | 1.55 | Hypoxic | |
| 2 | 0 | 24.7 | 32.3 | 8.64 | Oxic |
| 11 | 22.9 | 32.3 | 8.31 | Oxic | |
| 23 | 19.2 | 32.8 | 10.27 | Oxic |
| Genus | Species | St. 1 | St. 2 | Functions | ||||
|---|---|---|---|---|---|---|---|---|
| Surface (%) | Middle (%) | Bottom (%) | Surface (%) | Middle (%) | Bottom (%) | |||
| Candidatus Nitrosomarinus | catalina | 1.27 | 16.86 | 42.50 | 10.36 | 7.25 | 46.55 | |
| Nitrosopumilus | sp. Nsub | 0.45 | 4.17 | 8.22 | 2.27 | 1.63 | 8.72 | |
| Methanosarcina | Unclassified. Methanosarcina | 1.96 | 2.21 | 1.04 | 1.89 | 2.00 | 0.83 | |
| Methanobrevibacter | arboriphilus | 1.79 | 1.47 | 0.73 | 1.91 | 1.83 | 0.81 | |
| Methanococcus | maripaludis | 1.94 | 1.35 | 0.80 | 1.82 | 1.69 | 0.74 | |
| Methanobrevibacter | smithii | 1.78 | 1.37 | 0.81 | 1.66 | 1.55 | 0.82 | |
| Methanobrevibacter | oralis | 1.45 | 1.65 | 0.61 | 1.21 | 1.22 | 0.62 | |
| Methanosarcina | barkeri | 1.46 | 0.89 | 0.64 | 1.25 | 1.25 | 0.54 | |
| Candidatus Nitrosopelagicus | brevis | 0.44 | 0.28 | 1.84 | 0.88 | 0.78 | 1.66 | |
| Methanobrevibacter | Unclassified. Methanobrevibacter | 1.31 | 0.83 | 0.59 | 1.24 | 1.03 | 0.60 | |
| Nitrosopumilus | Unclassified. Nitrosopumilus | 0.56 | 0.89 | 1.28 | 0.80 | 0.67 | 1.36 | AOA |
| Methanobacterium | Unclassified. Methanobacterium | 1.07 | 0.84 | 0.64 | 1.13 | 1.28 | 0.55 | |
| Halorubrum | Unclassified. Halorubrum | 1.44 | 0.76 | 0.56 | 0.91 | 1.18 | 0.49 | |
| Methanosarcina | mazei | 1.03 | 1.13 | 0.52 | 0.90 | 0.90 | 0.45 | |
| Methanobrevibacter | olleyae | 0.91 | 0.80 | 0.47 | 0.79 | 0.85 | 0.40 | |
| Haloferax | Unclassified. Haloferax | 0.87 | 0.76 | 0.40 | 0.71 | 0.79 | 0.35 | |
| Sulfolobus | islandicus | 0.86 | 0.80 | 0.34 | 0.80 | 0.59 | 0.37 | |
| Nitrosopumilus | Candidatus Nitrosopumilus adriaticus | 0.59 | 0.57 | 0.64 | 0.60 | 0.54 | 0.69 | AOA |
| Methanocaldococcus | villosus | 0.88 | 0.59 | 0.37 | 0.76 | 0.65 | 0.36 | |
| Methanobrevibacter | curvatus | 0.67 | 0.56 | 0.38 | 0.71 | 0.80 | 0.36 | |
| Unclassified. Natrialbaceae | 0.90 | 0.60 | 0.36 | 0.62 | 0.66 | 0.28 | ||
| Methanobacterium | formicicum | 0.71 | 0.59 | 0.32 | 0.77 | 0.59 | 0.29 | |
| Methanobrevibacter | cuticularis | 0.75 | 0.61 | 0.28 | 0.71 | 0.54 | 0.31 | |
| Methanobrevibacter | sp. 87.7 | 0.58 | 0.85 | 0.24 | 0.56 | 0.59 | 0.35 | |
| Methanobacterium | paludis | 0.51 | 1.28 | 0.16 | 0.41 | 0.57 | 0.19 | |
| Methanobrevibacter | filiformis | 0.71 | 0.43 | 0.34 | 0.60 | 0.57 | 0.34 | |
| Nitrosopumilus | Candidatus Nitrosopumilus salaria | 0.37 | 0.55 | 0.56 | 0.49 | 0.44 | 0.52 | AOA |
| Candidatus Nitrosoarchaeum | limnia | 0.64 | 0.54 | 0.39 | 0.50 | 0.39 | 0.41 | |
| Candidatus Nitrosoarchaeum | koreensis | 0.48 | 0.54 | 0.40 | 0.50 | 0.47 | 0.40 | |
| Methanobrevibacter | millerae | 0.60 | 0.49 | 0.37 | 0.60 | 0.48 | 0.23 | |
| Methanobrevibacter | wolinii | 0.49 | 0.46 | 0.28 | 0.54 | 0.60 | 0.27 | |
| Methanosphaera | sp. WGK6 | 0.68 | 0.40 | 0.27 | 0.54 | 0.45 | 0.25 | |
| Methanocaldococcus | bathoardescens | 0.42 | 0.61 | 0.20 | 0.54 | 0.42 | 0.20 | |
| Methanocaldococcus | fervens | 0.43 | 0.69 | 0.20 | 0.37 | 0.46 | 0.21 | |
| Candidatus Methanoperedens | nitroreducens | 0.43 | 0.50 | 0.25 | 0.43 | 0.40 | 0.21 | MOA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.-H.; Yoon, S.-P. Preliminary Insights into Summer Archaeal Communities in Eutrophic Jinhae Bay, Korea. Diversity 2025, 17, 429. https://doi.org/10.3390/d17060429
Lim J-H, Yoon S-P. Preliminary Insights into Summer Archaeal Communities in Eutrophic Jinhae Bay, Korea. Diversity. 2025; 17(6):429. https://doi.org/10.3390/d17060429
Chicago/Turabian StyleLim, Jae-Hyun, and Sang-Pil Yoon. 2025. "Preliminary Insights into Summer Archaeal Communities in Eutrophic Jinhae Bay, Korea" Diversity 17, no. 6: 429. https://doi.org/10.3390/d17060429
APA StyleLim, J.-H., & Yoon, S.-P. (2025). Preliminary Insights into Summer Archaeal Communities in Eutrophic Jinhae Bay, Korea. Diversity, 17(6), 429. https://doi.org/10.3390/d17060429