
Systematic Revision of the Oligocene Billfishes (Istiophoriformes: Palaeorhynchidae) from Romania


Abstract
:1. Introduction
Geological Setting
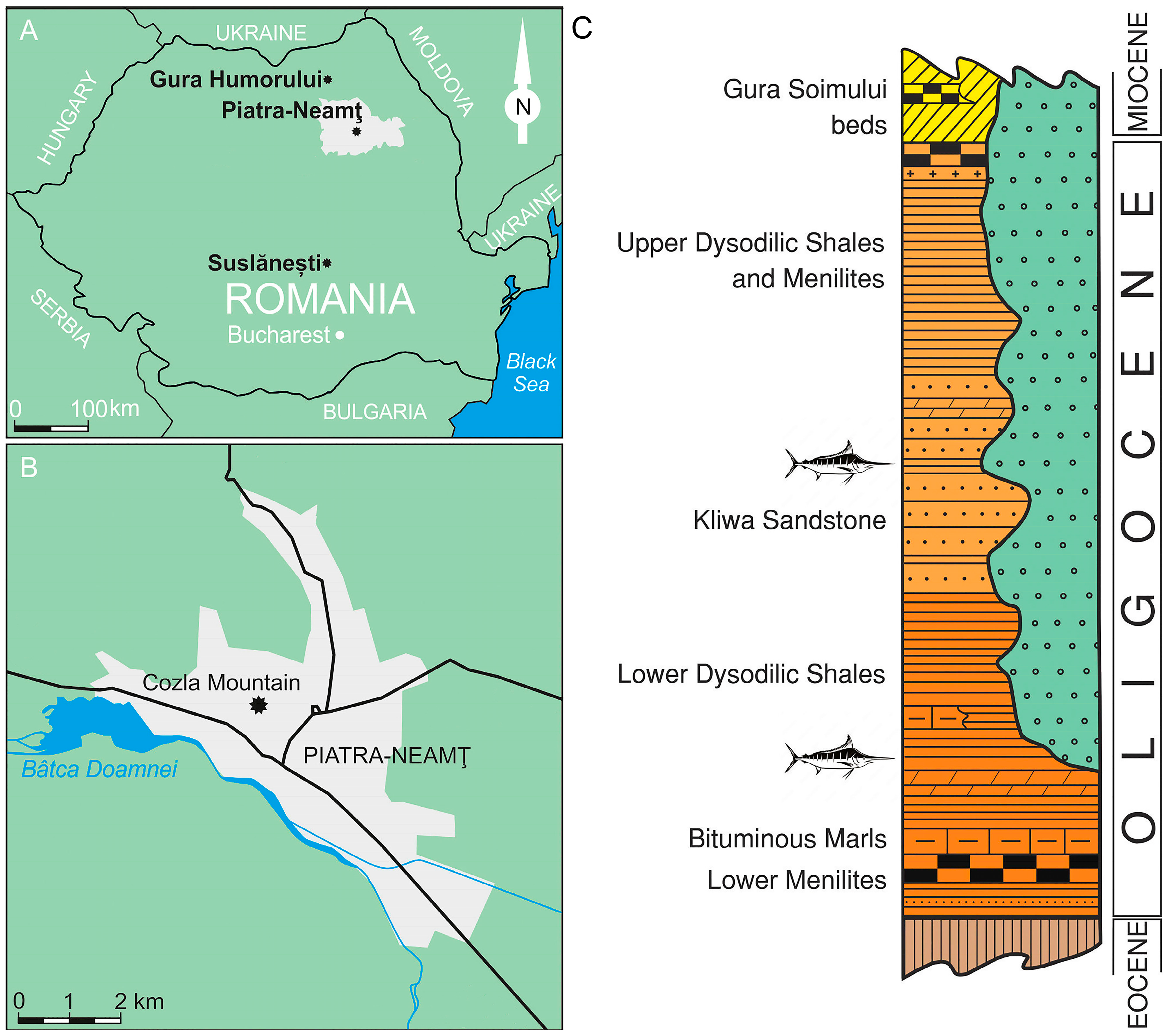
2. Materials and Methods
3. Systematic Paleontology
- Superorder Acanthopterygii Rosen and Patterson, 1969 [30]
- Order Istiophoriformes Betancur-R et al., 2013 [11]
- Diagnosis: The following is condensed from the overview of [1]. Istiophoriformes have premaxillaries and maxillaries tightly bound in a non-protrusible, elongated rostrum. The body shape is described as slender fusiform [7], which means slender and elongated but not snake-like. Vertebrae are elongated and longer than they are deep. Neural and haemal spines are not simple but have various modifications, depending on the istiophoriform family. The caudal skeleton has a high degree of fusion, with two smaller hypural plates in †Hemingwaya. At the same time, other istiophoriforms have one massive hypural plate consisting of four or all five hypurals fused into one. The caudal fin is hypurostegic.
- †Family Palaeorhynchidae Gunther, 1880 [31]Diagnosis: The following is taken from [32]. Palaeorhynchidae have an enlarged and downturned maxillary flange, a continuous soft dorsal fin, which can be divided into two sections: an anterior one of longer ceratotrichia and a posterior one of shorter, more tightly packed rays. A fan-shaped pterygiophore bears each ray in the anterior section with three radiating ridges. The pterygiophores of the caudal section appear to be simple, wedge-like structures tapered away from the axial skeleton. The neural and haemal spines bear a caudal, ovoid-shaped extension.
- †Homorhynchus Van Beneden, 1873 [17]Type species: †Homorhynchus bruxelliensis van Beneden, 1873 [17], p. 207, by monotypy.Composition: †Homorhynchus deshayesi (Agassiz, 1844) [33] from the Middle Eocene of France and Belgium and †Homorhynchus colei (Aggasiz, 1844) [33] from the Early Oligocene of Switzerland and Russia [1,32,34].Diagnosis from [15,32,34]: The maximum body length reaches ~60 cm; length is 8–12 times the body height; rostrum markedly exceeds lower jaw; vertebrae: 50–55 in total; two pterygiophores per interneural spaces in anterior part of dorsal fin, respectively, per interhaemal spaces of the anterior section of the anal fin. Dorsal fin-ray (D1) count ≈ 2 × vertebral count; there are no true D2 (second dorsal fin) and A2 (second anal fin), but the only continuous dorsal fin and only continuous anal fin are both divided in a long anterior section and short posterior one, surrounding the caudal peduncle. In the posterior section, there are more than two pterygiophores per interneural space or interhaemal space; hypurostegic caudal fin; pectoral and pelvic fins present.
- †Homorhynchus colei (Agassiz, 1844) [33]1833–1844 †Palaeorhynchum colei Agassiz: 85, p1.32, Figure 1 [33].1833–1844 †Palaeorhynchum microspondylum Agassiz: 85, pl. 34a, Figure 2 [33].1886 †Hemirhynchus colei (Agassiz, 1844), Wettstein, 78. [35]1960 †Homorhynchus colei Danil’chenko, 161. p1. XV, Figure 2 [15].1977 †Palaeorhynchus longirostris Agassiz, 1843, Ciobanu, p. 121, pl. XLII, Figure 1 [23].1977 †Palaeorhynchus glarisiaus Blainville, 1818, Ciobanu, p. 122, pl. XLIII, Figure 1 [23].2000b †Homorhynchus colei Ag., Monsch, p. 153, Figure 7.59 [34].2010 †Homorhynchus colei (Agassiz, 1844), Bannikov, p. 149, pl. XXVI, Figure 2 [36].2011 †Homorhynchus colei (Agassiz, 1844), Monsch and Bannikov, p. 291, Figure 42 [32].Diagnosis from [15,32]: As for the genus, plus the body length is approximately 8–10 times the maximum body depth. The head length is about 3–3.5 times the body depth; the skull is shallow: depth about two-thirds of maximum body depth [37]. A total of 54–55 (26 + 28–29) vertebrae (25 + 28 in [15]. The dorsal fin originates adjacent to the first vertebra.
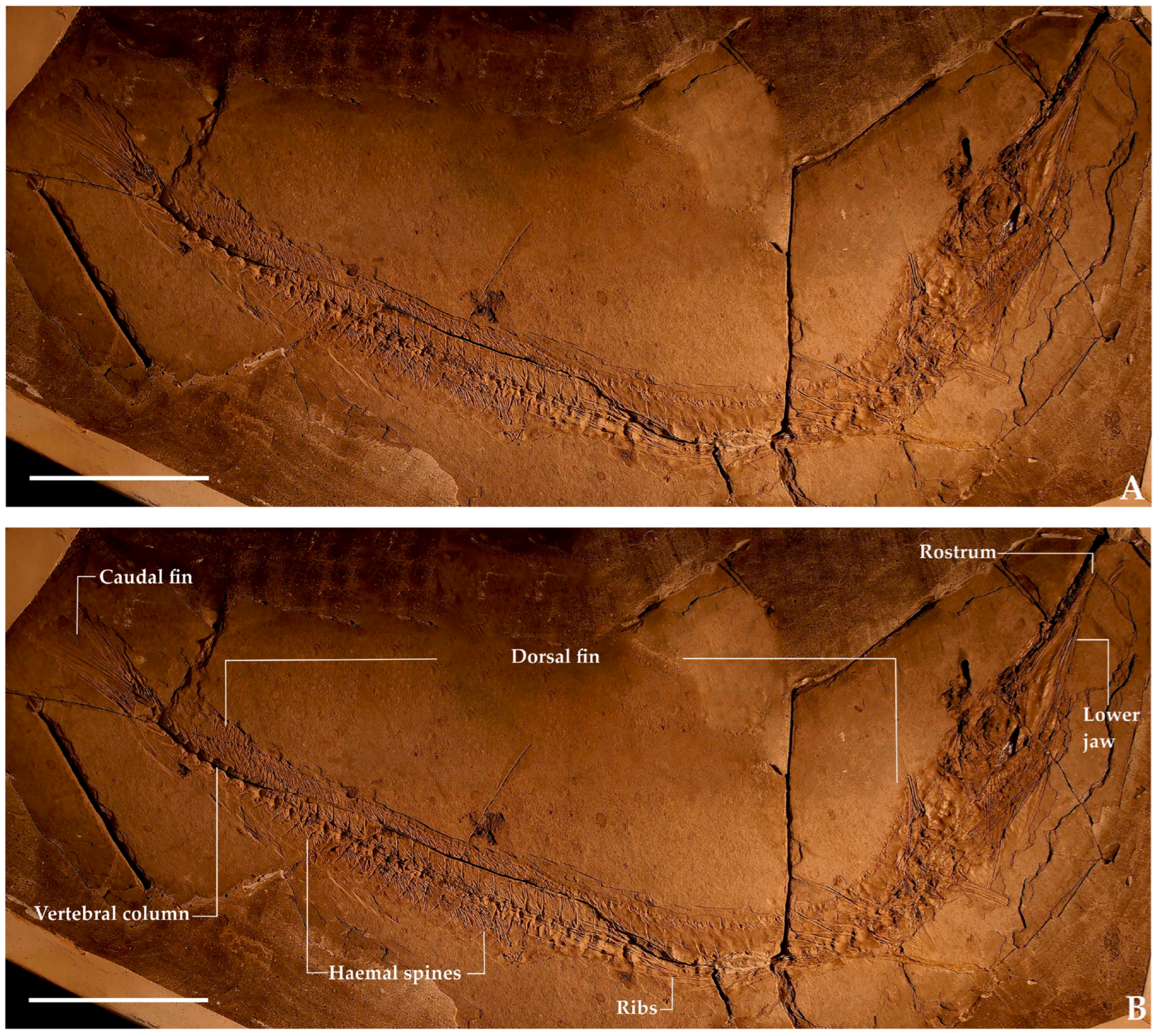
- Name in collection: †Palaeorhynchus glarisianus Blainville, 1818 [16]Number: MSNPN-PC 154 (Figure 2).Location in collection: Public exhibition.General overview: Complete fish on slab.Size: Length from distal tip of rostrum to last vertebra—375 mm.General body features: The body is incompletely preserved due to bending but appears slender and almost snake-like, tapering posteriorly, similar to modern billfish, but somewhat slimmer. The body is covered in large cycloid scales.Head: The relative head depth to body depth cannot be determined due to incomplete preservation in the anterior region. The skull roof is thick and ornamented with lines (shallow grooves). The rostrum is long and slender, though its full length is not entirely preserved. Preservation of the rostrum is poor, with much of its structure obscured by an irregular cavity filled in the restoration process. Small black structures in this area are likely artefacts from chemical reactions between the matrix and glue rather than teeth. The premaxillary flange is fragmented, with a small section overlapping the proximal dentary and angular. We infer that the original flange was considerably larger. The lower jaw is distinctly shorter than the rostrum.Vertebral column: The vertebrae are hourglass-shaped, with pronounced constrictions and stout neural and haemal spines. The total vertebral count is uncertain due to poor anterior preservation, but we estimate at least 24 precaudal vertebrae (possibly more) and 28 caudal vertebrae (including the urostyle). Precaudal vertebrae lack parapophyses, with ribs articulating directly. These ribs are thick, robust, and curve posteriorly. Neural and haemal spines exhibit leaf- or trapezium-shaped caudal lamellae, which are poorly visible. Because of this, a false first impression of forked spines is created. The haemal spines of the first caudal vertebrae are obscured by the first 21 anal-fin pterygiophores, which are compressed into the body cavity. From the 13th caudal vertebra onward, neural and haemal spines become spatula-like, transitioning to slender trapezoidal shapes with posterior-pointing apices in the most caudal spines.Unpaired fins: Both the dorsal and anal fins show two or three pterygiophores per interneural or interhaemal space. Their ceratotrichia are relatively short (shorter than maximum body depth, unlike †Palaeorhynchus). The dorsal fin preserves 71 rays, as the posterior section is missing and the anterior region is poorly preserved. The anal fin has four rays anterior to the first haemal spine, with a total of 29 rays preserved. In the caudal section, the anal fin appears compressed inward toward the vertebral column. The caudal fin complex is unclear; there may be one or two epurals, and the fifth hypural could be either autogenous or fused. Much of the hypural plate is concealed by the caudal fin. The caudal fin morphology is unclear due to displaced lepidotrichia, but we estimate ~23 rays.Pectoral fin and girdle: The shoulder girdle is twisted and visible ventrally, with two partial pectoral fins (the best-preserved fin has 11 lepidotrichia, though the original number was likely higher).The pelvic fin is not preserved.Remarks: Ciobanu [23] mentioned the following meristics for this specimen: vertebrae: 52–54 (25 + 29); dorsal fin: 60?; anal fin: 40/40; pectoral fin: 12; pelvic fin: seven; caudal fin: 11/11.Our meristic counts differ from Ciobanu’s but correspond well on most points. In our opinion, the differences are caused by the difficulty of investigating poorly preserved specimens, which we overcame by examining the specimens by all of us and consulting with one another. We have not seen the pelvic fins with seven rays that Ciobanu reported. The meristic counts and other diagnostic characters still fit the diagnostic range of this species (see also Table 1), and the geographical and geological provenance correspond. Although preservation is comparatively poor, we have no reason to believe this is another species other than the above-mentioned †Homorhynchus colei.
- Name in collection: †Palaeorhynchus longirostris Agassiz, 1843 [33]Number: MSNPN-PC 155 (Figure 3).Location in the collection: Depository.General overview description: Complete fish on slab with caudal fin missing.Size: Length from distal tip of rostrum to last preserved (50th) vertebra—610 mm.General body features: The specimen is slender, elongated, and covered in large cycloid scales. Its rostrum is elongated, constituting approximately 64% of the total head length. A prominent premaxillary flange extends over the proximal and angular regions of the dentary. The lower jaw is significantly shorter than the rostrum, reaching only about 43% of the rostrum’s proximal length.Head: The head (incorporating branchiostegal rays) displays a depth of roughly two-thirds of the body’s maximum depth (66%). The skull roof is robust but lacks surface ornamentation due to preservation limitations.Vertebral column: The vertebrae are hourglass-shaped, featuring pronounced constrictions and sturdy neural and haemal spines. While poor preservation obscures the anterior vertebral column, 28 precaudal vertebrae are inferred. Of the caudal vertebrae, 22 remain intact, though the original count was likely greater. Precaudal vertebrae lack parapophyses, with ribs directly articulating to the centra. These ribs are thickened, robust, and posteriorly curved. Neural spines exhibit faint leaf-shaped caudal lamellae, weakly visible from the third caudal neural spine onward, superficially resembling forked spines. This is a false first impression because the preservation of the lamellae is relatively poor. Haemal spine lamellae and the caudal complex are absent due to preservation gaps.Unpaired fins: Both dorsal and anal fins display two to three pterygiophores per interneural or interhaemal space. Their ceratotrichia are comparatively short (shorter than the body’s maximum depth, distinguishing it from †Palaeorhynchus). The dorsal fin retains 77 rays, though the original number was higher due to missing posterior and degraded anterior sections. The anal fin preserves 39 rays, with seven positioned anterior to the first haemal spine; the complete count remains undetermined.Pectoral fin and girdle: The shoulder girdle is ventrally exposed and distorted, with partial pectoral fins retaining nine lepidotrichia.Pelvic fin elements are absent in the specimen.Remarks: This specimen is catalogued in the Paleontological Collection of the Natural Sciences Museum of Piatra-Neamț as †Palaeorhynchus longirostris Agassiz, 1843 [33]. The collection records include no descriptions or additional information besides collecting data. While preservation quality limits detailed morphological analysis, the specimen’s identification as †Homorhynchus colei remains strongly supported. Its meristic values align with the documented parameters of the species, and its geological setting and geographic origin are consistent with this taxonomic assignment.

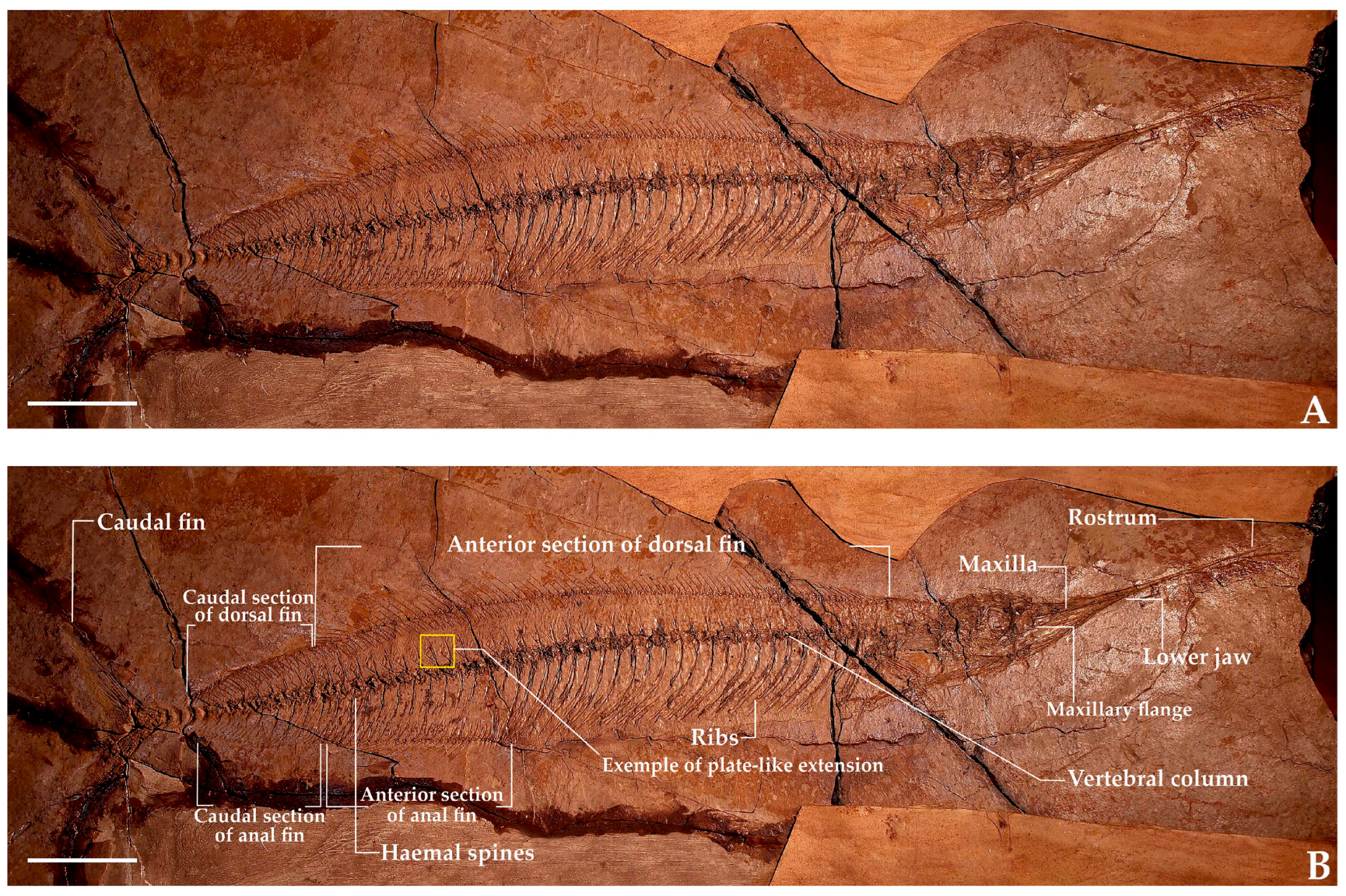
- Name in collection: †Palaeorhynchus longirostris Agassiz, 1843 [33]Number: MSNPN-PC 197 (Figure 4).Location in collection: Public exhibition.General overview: Complete fish on slab.Size: Length from distal tip of rostrum to last preserved vertebra—525 mm.General body features: The specimen exhibits a slightly fusiform, slender, and elongated body, likely covered by large scales.Head: The head (including branchiostegal rays) is shallow, approximating 64% of the maximum body depth. The skull roof is thick and ornamented with lines (shallow grooves). The rostrum is elongated, measuring 67% of total head length. The premaxillary flange is prominent, partially obscuring the dentary, while the lower jaw is short, extending only to the midpoint of the rostrum.Vertebral column: The vertebrae (total count: 27 + 28) are hourglass-shaped, with pronounced constrictions and stout neural and haemal spines. Precaudal vertebrae lack parapophyses, with ribs articulating directly; these are robust, caudally bent, and thick. Neural and haemal spines exhibit leaf-shaped caudal lamellae, though they are poorly preserved, creating a false impression of forked spines. The caudal complex is incompletely preserved, obscuring certain elements: there are possibly one to two epurals and ambiguity regarding the autonomy of the fifth hypural. The posterior hypural plate is fragmented.Unpaired fins: The dorsal fin contains 107 preserved rays (incomplete), with two to three pterygiophores per interneural space. The ceratotrichia are relatively short (shorter than the maximum body depth, contrasting with †Palaeorhynchus). The anal fin has twenty-nine rays preserved, with four rays anterior to the first haemal spine; the total count is indeterminate due to fragmentation. The caudal fin is forked, near-semilunate (this shape is more pronounced in smaller specimens), hypurostegic, with an estimated 30 rays.Pectoral fin and girdle: The pectoral fin, with 13 preserved lepidotrichia, is likely incomplete.The pelvic fin comprises two short lepidotrichia.Remarks: The specimen was initially described as †Palaeorhynchus longirostris by [23], with the following meristic counts: vertebrae: 60–61 (34 + 27); dorsal fin: 94/94; anal fin: 40; pectoral fin: 14; pelvic fin: seven; caudal fin: 15/16. Our meristic counts again differ from Ciobanu’s, probably because of the difficulty of investigating poorly preserved specimens, which we overcame by examining the specimens together. Subsequently, the specimen was reidentified as †Homorhynchus colei by Bannikov in [32], though without explicit morphological justification. Our analysis confirms this taxonomic assignment. At the same time, we note a minor discrepancy in vertebral count from the diagnosis in [32] (our 27 + 28 vs. 26 + 28–29 in [32]). All other diagnostic characters, including the identical geological provenance, support classification as †H. colei.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Vertebral Count | Key Morphological Features | Geographic Range | Age Range | References |
|---|---|---|---|---|---|
| †H. colei | 26–28 + 28–29 | None specifically, as other species are not well known. †H. colei is identified by its geographical and geological range. | Switzerland, Russia, Romania | Early Oligocene | [15,32] |
| †H. bruxelliensis | >21 caudal | Twenty caudal-fin rays. The species is poorly known. No other features are clearly described. | Belgium | Middle Eocene (early Lutetian) | [17] |
| †H. deshayesi | Not specified | Not specified. | France | Middle Eocene | [33] |
- †Palaeorhynchus de Blainville, 1818 [16]Type species: †Palaeorhynchum glarisianus de Blainville, 1818, p. 314, [16], by monotypy.Composition: We have recognised seven species, namely †Palaeorhynchus zorzini Fierstine, Bannikov, Monsch, 2008 [38] (Eocene of Italy); †Palaeorhynchus senectus Danil’chenko, 1962 [39] (Middle Eocene of Georgia); †Palaeorhynchus parini Bannikov, 1992 [40] (Middle Eocene of North Caucasus, Russia); †Palaeorhynchus zitteli (Kramberger, 1879) [41] (Oligocene of Poland, Romania, Russia); †Palaeorhynchus glarisianus de Blainville, 1818 [16] (Oligocene of France, Switzerland, Romania, possibly Germany, see [42]); †Palaeorhynchus humorensis Brustur and Grigorescu, 1973 [24] (Oligocene of Romania); †Palaeorhynchus altivelis Arambourg, 1967 [43] (Oligocene of Iran). Fierstine [1] mentioned nine to eleven species but did not mention them, so we do not know which taxonomy he applied; he also mentioned occurrences of this genus in the Early Oligocene of the former and possibly the Early Miocene of Germany and Switzerland.Diagnosis from [32]: The upper and lower jaws are straight, elongated, and equal in length. The vertebrae number 50–60. Dorsal fin elements roughly match the number of vertebrae.
- †Palaeorhynchus humorensis Brustur and Grigorescu, 1973 [24]Name in collection: †Palaeorhynchus humorensis Brustur and Grigorescu, 1973 [24].Number: CLPUB–LPB/II640.Location in the collection: The Geology and Geophysics Faculty Hall, University of Bucharest.Diagnosis after [24]: Elongated, fusiform body, with a maximum height of 8.2% of the body length excluding the caudal fin. The head length is 28% of the vertebral column length. The rostrum, formed by two elongated maxillaries, is 58% of the skull length. The large orbit, with nearly equal horizontal and vertical diameters, is 22.6% of the postrostral part length. There are 61 vertebrae, 21 of which are precaudal. The dorsal fin begins anteriorly, above the middle of the orbit, extending to the middle of the caudal peduncle. The anal fin is double; the first anal fin is more developed, with its anterior rays inserting opposite the 13th precaudal vertebra. The second anal fin is located behind the first, at a distance corresponding to the length of seven vertebrae. The body is covered with small cycloid scales.Remarks: For a detailed analysis and description of the species, see [24]. The most notable distinction in †P. humorensis is the presence of a double anal fin, a feature undocumented in other †Palaeorhynchus species. This could represent either a specialised adaptation (e.g., for improved manoeuvrability) or a unique taxonomic characteristic. However, the apparent “double” fin might also result from misinterpretation or a fossil preservation artefact, as no other members of the †Palaeorhynchidae family exhibit this trait. Because it is not yet clear if the double fins are a true feature and because we have only one specimen, we cannot draw any conclusions on the function of this feature.Additionally, †P. humorensis possesses 61 vertebrae, exceeding the typical range of 45–60 observed in other Palaeorhynchus species (see Table 2). This suggests either a more elongated body or a higher degree of segmentation. Detailed morphometric data for †P. humorensis (e.g., body height at 8.2%, rostrum length at 58% of total length) lack direct equivalents in other species descriptions, complicating comparative analysis. These proportions indicate a highly streamlined body shape, possibly reflecting distinct ecological or locomotory adaptations.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MSNPN-PC | Palaeontological Collection of the Natural Sciences Museum Piatra-Neamţ; |
| CLPUB–LPB | Palaeontological Collection of the Geology and Geophysics Faculty Hall, University of Bucharest. |
References
- Fierstine, H.L. Fossil History of Billfishes (Xiphioidei). Bull. Mar. Sci. 2006, 79, 433–453. [Google Scholar]
- Monsch, K.A. Revision of the Scombroid Fishes from the Cenozoic of England. Trans. R. Soc. Edinb. Earth Sci. 2005, 95, 445–489. [Google Scholar] [CrossRef]
- Monsch, K.A.; Fierstine, H.L.; Weems, R.E. Taxonomic Revision and Stratigraphic Provenance of ‘Histiophorus rotundus’ Woodward 1901 (Teleostei, Perciformes). J. Vertebr. Paleontol. 2005, 25, 274–279. [Google Scholar] [CrossRef]
- Fierstine, H.L. A New †Aglyptorhynchus (Perciformes: Scombroidei: †?Blochiidae) from the Late Oligocene of Oregon. J. Vertebr. Paleontol. 2001, 21, 24–33. [Google Scholar] [CrossRef]
- Fierstine, H.L. A New Aglyptorhynchus (Perciformes: Scombroidei) from the Lincoln Creek Formation (Late Oligocene, Washington, U.S.A.). J. Vertebr. Paleontol. 2005, 25, 288–299. [Google Scholar] [CrossRef]
- Gosline, W.A. The Suborders of Perciform Fishes. Proc. U. S. Natl. Mus. 1968, 124, 1–78. [Google Scholar] [CrossRef]
- 5: Billfishes of the World: An Annotated and Illustrated Catalogue of Marlins, Sailfishes, Spearfishes and Swordfishes Known to Date. In FAO Species Catalogue (FAO Fisheries Synopsis); Nakamura, I., Ed.; FAO: Rome, Italy, 1985; ISBN 978-92-5-102232-0. [Google Scholar]
- Schultz, O. Der Erstnachweis Eines Schwertfischverwandten Aus Dem Südostspanischen Pliozän: Makaira sp. (Istiophoridae, Xiphioidei, Osteichthyes). Doc. Naturae 1996, 1, 13–23. [Google Scholar]
- Collette, B.; McDowell, J.; Graves, J. Phylogeny of recent billfishes (Xiphioidei). Bull. Mar. Sci. 2006, 79, 455–468. [Google Scholar]
- Near, T.J.; Eytan, R.I.; Dornburg, A.; Kuhn, K.L.; Moore, J.A.; Davis, M.P.; Wainwright, P.C.; Friedman, M.; Smith, W.L. Resolution of Ray-Finned Fish Phylogeny and Timing of Diversification. Proc. Natl. Acad. Sci. USA 2012, 109, 13698–13703. [Google Scholar] [CrossRef]
- Betancur-R, R.; Broughton, R.E.; Wiley, E.O.; Carpenter, K.; López, J.A.; Li, C.; Holcroft, N.I.; Arcila, D.; Sanciangco, M.; Cureton Ii, J.C.; et al. The Tree of Life and a New Classification of Bony Fishes. PLoS Curr. 2013, 5, 26288. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 1st ed.; Wiley: Hoboken, NJ, USA, 2016; ISBN 978-1-118-34233-6. [Google Scholar]
- Wu, B.; Feng, C.; Zhu, C.; Xu, W.; Yuan, Y.; Hu, M.; Yuan, K.; Li, Y.; Ren, Y.; Zhou, Y.; et al. The Genomes of Two Billfishes Provide Insights into the Evolution of Endothermy in Teleosts. Mol. Biol. Evol. 2021, 38, 2413–2427. [Google Scholar] [CrossRef]
- De Gracia, C.; Villalobos-Segura, E.; Ballen, G.A.; Carnevale, G.; Kriwet, J. Phylogenetic Patterns in Fossil and Living Billfishes (Istiophoriformes, Istiophoridae): Evidence from the Central Mediterranean. Pap. Palaeontol. 2024, 10, e1559. [Google Scholar] [CrossRef]
- Danil’cenko, P.G. Bony Fishes of the Maikop Deposits of the Caucasus; Originally in Russian, Translated by A. Mercado, Israel Program for Scientific Translations, Jerusalem, 1967; Academy of Sciences of the U.S.S.R.; Transactions of the Paleontological Institute: Moscow, Russia, 1960; Volume 78. [Google Scholar]
- Blainville, H.M. Poissons Fossiles. In Noveau Dictionnaire d’Histoire Naturelle, Appliqué Aux Arts, à l’Agriculture, à l’Economie Rurale et Domestique, à La Médicine; Chez Déterville: Paris, France, 1818; Volume 27. [Google Scholar]
- Van Beneden, P.J. Notice Sur Un Nouveau Poisson du Terrain Bruxellien. Bull. Acad. R. Belg. 1873, 35, 255–259. [Google Scholar]
- Casier, E. Faune Ichthyologique Du London Clay; British Museum (National History): London, UK, 1966. [Google Scholar]
- Gottfried, M.D.; Fordyce, R.E.; Rust, S. A New Billfish (Perciformes, Xiphioidei) from the Late Oligocene of New Zealand. J. Vertebr. Paleontol. 2012, 32, 27–34. [Google Scholar] [CrossRef]
- Cosmovici, L.C. Les Couches à Poissons Des Monts Petricica et Cozla, District de Neamtz, Ville de Peatra. Bul. Soc. Medici Nat. Iași 1887, I, 96–105. [Google Scholar]
- Simionescu, I.T. Asupra Câtorva Pesci Fosili Din Terțiarul Românesc. Academia Română Publicaţiile Fondului Adamachi 1904, XII, 205–222. [Google Scholar]
- Paucă, M. Die Fossile Fauna und Flora Aus Dem Oligozan von Suslănești-Muscel in Rumänien. Eine Systematische Und Paläobiologische Studie. Anuarul l’Institut Géologique Roumanie București 1934, 16, 672–768. [Google Scholar]
- Ciobanu, M. Fauna Fosilă din Oligocenul de la Piatra Neamț; Editura Academiei Republicii Socialiste România: București, Romania, 1977. [Google Scholar]
- Brustur, T.; Grigorescu, D. Une Nouvelle Espèce du Genre Palaeorhynchus: Palaeorhynchus Humorensis dans les Dépots Oligocènes de la Zone de Gura Humorului. Rev. Roum. Géol. 1973, 17, 99–114. [Google Scholar]
- Săndulescu, M. Geotectonica României; Editura Tehnică: București, Romania, 1984. [Google Scholar]
- Mutihac, V.; Ionesi, L. Geologia României; Editura Tehnică: București, Romania, 1974. [Google Scholar]
- Bădescu, D. Evoluția Tectono-Stratigrafică a Carpaților Orientali în Decursul Mezozoicului și Neozoicului; Editura Economică: București, Romania, 2005. [Google Scholar]
- Grasu, C.; Catană, C.; Grinea, D. Flișul Carpatic. Petrografie și Considerații Economice; Editura Tehnică: București, Romania, 1988. [Google Scholar]
- Grasu, C.; Miclăuș, C.; Florea, F.; Șaramet, M. Geologia și Valorificarea Economică a Rocilor Bituminoase din România; Editura Universității Alexandru Ioan Cuza: Iași, Romania, 2007. [Google Scholar]
- Rosen, D.E.; Patterson, C. The structure and relationships of the paracanthopterygian fishes. Bull. Am. Mus. Nat. Hist 1969, 141, 357–474. [Google Scholar]
- Günther, A. An Introduction to the Study of Fishes; R&R Clark: Edinburgh, Scotland, 1880. [Google Scholar]
- Monsch, K.A.; Bannikov, A.F. New Taxonomic Synopses and Revision of the Scombroid Fishes (Scombroidei, Perciformes), Including Billfishes, from the Cenozoic of Territories of the Former USSR. Earth Environ. Sci. Trans. R. Soc. Edinb. 2011, 102, 253–300. [Google Scholar] [CrossRef]
- Agassiz, L. Recherches sur les Poissons Fossiles; Imprimerie de Petitpierre: Neuchâtel, Switzerland, 1833; Volume 5. [Google Scholar]
- Monsch, K.A. The Phylogeny of the Scombroid Fishes. Ph.D. Thesis, University of Bristol, Bristol, UK, 2000. [Google Scholar]
- Wettstein, A. Über Die Fischfauna des Tertiären Glanerschiefers. Abh. Senckenb. Paläont. Ges. 1886, 8, 1–103. [Google Scholar]
- Bannikov, A.F. Iskopayemye Pozvonochnye Rossii i Sopredelnykh Stran. Iskopayemye Kolyuchepyorye Ryby (Teleostei, Acanthopterygii); GEOS: Moscow, Russia, 2010. [Google Scholar]
- Woodward, A.S. Catalogue of the Fossil Fishes in the British Museum (Natural History); British Museum (Natural History): London, UK, 1901. [Google Scholar]
- Fierstine, H.L.; Bannikov, A.F.; Monsch, K.A. A New Species of the Palaeorhynchid Fish Genus Palaeorhynchus from the Late Early Eocene of Bolca in Northern Italy (Xiphioidei, Palaeorhynchidae). Misc. Paleontol. N.9 Studi Ric. Giacimenti Terziari Bolca 2008, 12, 7–28. [Google Scholar]
- Danil’cenko, P.G. Ryby Dabakhanskoi Svity Gruzii. Paleontol. Zhurnal 1962, 1, 111–126. [Google Scholar]
- Bannikov, A.F. Novyj Vid Roda Palaeorhynchus (Perciformes, Palaeorhynchidae) Iz Verkhnego Eotsena Severnogo Kavkaza. Vopr. Ikhtiologii 1992, 32, 27–32. [Google Scholar]
- Kramberger, D. Beiträge zur Kenntnis der fossilen Fische der Karpathen. Palaeontographica 1879, 26, 51–68. [Google Scholar]
- Monsch, K.A.; Micklich, N. Catalogue of the Scombroid Fishes from the Grube Unterfeld (»Frauenweiler«) Fossil Site at Rauenberg (Baden-Württemberg, S. Germany). Kaupia. Darmstädter Beiträge Naturkunde 2018, 22, 7–154. [Google Scholar]
- Arambourg, C. Résultats Scientifiques de la Mission C. arambourg en Syrie et en Iran: (1938–1939). 2. Les Poissons Oligocènes de l’Iran; Muséum National d’Histoire Naturelle: Paris, France, 1967; Volume 8 of Notes et Mèmoires Moyen-Orient. [Google Scholar]
- Pharisat, A. La Paleoichthyofaune du Rupelien Marin de Froidefontaine (Territoire de Belfort). Taxonomie et Populations, Genese Du Gisement Implications Paleobiographiques. Ann. Sci. l’Université Fr. Comté. Géologie 1991, 4, 13–97. [Google Scholar]
- Carnevale, G.; Bannikov, A.F.; Marrama, G.; Tyler, J.C.; Zorzin, R. The Pesciara-Monte Postale Fossil-Lagerstatte: 2. Fishes and Other Vertebrates. Bolca Foss. Lagerstätten Window Eocene World 2014, 4, 37–64. [Google Scholar]
- Papazzoni, C.A.; Carnevale, G.; Fornaciari, E.; Giusberti, L.; Trevisani, E. The Pesciara-Monte Postale Fossil-Lagerstätte: 1. Biostratigraphy, Sedimentology and Depositional Model. Bolca Foss. Lagerstätten Window Eocene World 2014, 4, 29–36. [Google Scholar]

| Species Name | Vertebral Count | Key Morphological Features | Geographic Range | Age Range | References |
|---|---|---|---|---|---|
| †P. altivelis | 53–54, of which 32–33 caudal | Anal fin originating under the penultimate precaudal vertebra | Iran | Oligocene | [43] |
| †P. humorensis | 61, of which 40 caudal | Double anal fin (1st originating under 13th vertebra) | Romania (Gura Humorului) | Oligocene | [24] |
| †P. glarisianus | 50–60, of which 35–36 caudal | Anal fin originating under the 18th precaudal vertebra | Switzerland, France | Early Oligocene | [44] |
| †P. zitteli | 58–61 (?62) vertebrae, of which 33–35 caudal | Anal fin originating under 5th–7th before last precaudal vertebra | Poland, Russia, Romania? | Early Oligocene | [32] |
| †P. parini | 58–60 vertebrae, of which 35–37 caudal | Anal fin originating under third or fourth before last precaudal vertebra | Northern Caucasus (Russia) | Middle Eocene | [32] |
| †P. senectus | 50–60? | Elongated body, long rostrum, continuous dorsal fin | Georgia | Middle Eocene | [32] |
| †P. zorzini | 50, of which 29 caudal | Fewer vertebrae (most primitive), long rostrum, continuous dorsal fin | Northern Italy (Bolca) | Late Early Eocene (Ypresian, ~50.5 Ma) | [38] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grădianu, I.; Monsch, K.A.; Baciu, D.S. Systematic Revision of the Oligocene Billfishes (Istiophoriformes: Palaeorhynchidae) from Romania. Diversity 2025, 17, 393. https://doi.org/10.3390/d17060393
Grădianu I, Monsch KA, Baciu DS. Systematic Revision of the Oligocene Billfishes (Istiophoriformes: Palaeorhynchidae) from Romania. Diversity. 2025; 17(6):393. https://doi.org/10.3390/d17060393
Chicago/Turabian StyleGrădianu, Ionuț, Kenneth A. Monsch, and Dorin Sorin Baciu. 2025. "Systematic Revision of the Oligocene Billfishes (Istiophoriformes: Palaeorhynchidae) from Romania" Diversity 17, no. 6: 393. https://doi.org/10.3390/d17060393
APA StyleGrădianu, I., Monsch, K. A., & Baciu, D. S. (2025). Systematic Revision of the Oligocene Billfishes (Istiophoriformes: Palaeorhynchidae) from Romania. Diversity, 17(6), 393. https://doi.org/10.3390/d17060393