
In Situ Investigation of Ecological and Molecular Stress Mechanisms Triggered by Marine Heat Waves in Adriatic Populations of the Mediterranean Mussel

 ,
,  ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
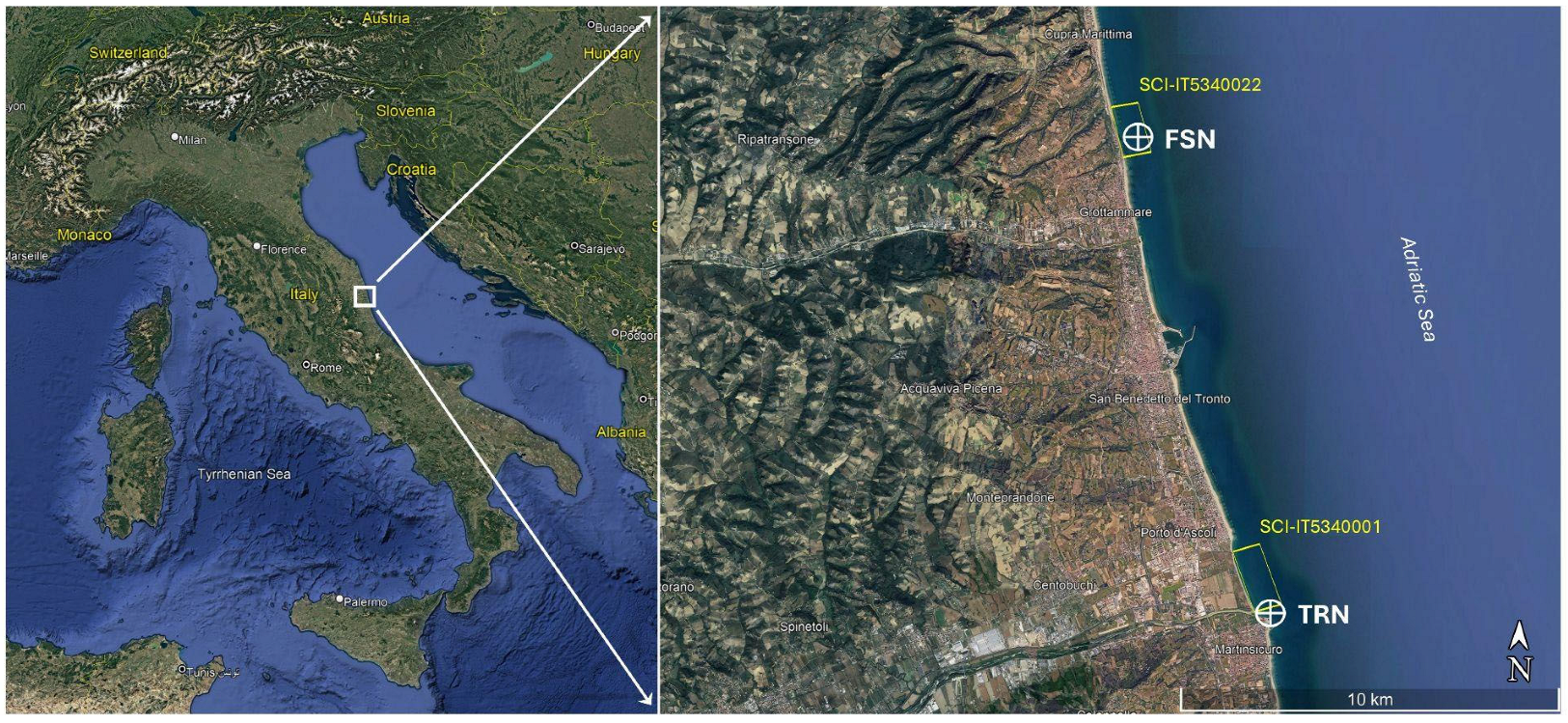
2.1. Study Area
2.2. Environmental Data and Analyses
2.3. Mussel Bed Coverage
2.4. Mussel Collection and Processing
2.5. Molecular Analyses
3. Results
3.1. Climatic Analysis
3.2. Mussel Coverage and Growing
3.3. Transcription Modulation over the Months
4. Discussion
4.1. Ecological Observation at the Population Level
4.2. Modulation of Molecular Biomarkers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, L.; von Schuckmann, K.; Abraham, J.P.; Trenberth, K.E.; Mann, M.E.; Zanna, L.; England, M.H.; Zika, J.D.; Fasullo, J.T.; Yu, Y.; et al. Past and future ocean warming. Nat. Rev. Earth Environ. 2022, 3, 776–794. [Google Scholar] [CrossRef]
- Hobday, A.J.; Oliver, E.C.; Gupta, A.S.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Holbrook, N.J.; Moore, P.J.; Thomsen, M.S.; Wernberg, T.; et al. Categorizing and naming marine heatwaves. Oceanography 2018, 31, 162–173. [Google Scholar] [CrossRef]
- Hobday, A.J.; Alexander, L.V.; Perkins, S.E.; Smale, D.A.; Straub, S.C.; Oliver, E.C.J.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Feng, M.; et al. A hierarchical approach to defining marine heatwaves. Prog. Oceanogr. 2016, 141, 227–238. [Google Scholar] [CrossRef]
- Malhi, Y.; Franklin, J.; Seddon, N.; Solan, M.; Turner, M.G.; Field, C.B.; Knowlton, N. Climate change and ecosystems: Threats, opportunities and solutions. Philos. Trans. R. Soc. A 2020, 375, 20190104. [Google Scholar] [CrossRef]
- Smale, D.A.; Wernberg, T.; Oliver, E.C.J.; Thomsen, M.; Harvey, B.P.; Straub, S.C.; Burrows, M.T.; Alexander, L.V.; Benthuysen, J.A.; Donat, M.G.; et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Change 2019, 9, 306–312. [Google Scholar] [CrossRef]
- Beever, E.A.; O’Leary, J.; Mengelt, C.; West, J.M.; Julius, S.; Green, N.; Managness, D.; Petes, L.E.; Stein, B.A.; Nicotra, A.B.; et al. Improving conservation outcomes with a new paradigm for understanding species’ fundamental and realized adaptive capacity. Conserv. Lett. 2016, 9, 131–137. [Google Scholar] [CrossRef]
- Greenhough, H.; Vignier, J.; Smith, K.F.; Brown, C.M.; Kenny, N.J.; Rolton, A. Multi-stressor dynamics: Effects of marine heatwave stress and harmful algal blooms on juvenile mussel (Perna canaliculus) survival and physiology. Sci. Total Environ. 2025, 964, 178590. [Google Scholar] [CrossRef]
- Lattos, A.; Papadopoulos, D.K.; Feidantsis, K.; Giantsis, I.A.; Georgoulis, I.; Karagiannis, D.; Michaelidis, B. Antioxidant Defense of Mytilus galloprovincialis Mussels Induced by Marine Heatwaves in Correlation with Marteilia Pathogen Presence. Fishes 2023, 8, 408. [Google Scholar] [CrossRef]
- Joyce, P.W.; Tong, C.B.; Yip, Y.L.; Falkenberg, L.J. Marine heatwaves as drivers of biological and ecological change: Implications of current research patterns and future opportunities. Mar. Biol. 2024, 171, 20. [Google Scholar] [CrossRef]
- Bracchetti, L.; Capriotti, M.; Fazzini, M.; Cocci, P.; Palermo, F.A. Mass Mortality Event of Mediterranean Mussels (Mytilus galloprovincialis) in the Middle Adriatic: Potential Implications of the Climate Crisis for Marine Ecosystems. Diversity 2024, 16, 130. [Google Scholar] [CrossRef]
- Galil, B.S.; Mienis, H.K.; Mendelson, M.; Gayer, K.; Goren, M. Here today, gone tomorrow-the Levantine population of the Brown mussel Perna perna obliterated by unprecedented heatwave. Aquat. Invasions 2022, 17, 174–185. [Google Scholar] [CrossRef]
- Garrabou, J.; Coma, R.; Bensossan, N.; Bally, M.; Chevaldonné, P.; Cigliano, M.; Diaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass mortality in Northwestern Mediterranean rocky benthic communities: Effects of the 2003 heat wave. Glob. Change Biol. 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Bonacci, O.; Bonacci, D.; Patekar, M.; Pola, M. Increasing Trends in Air and Sea Surface Temperature in the Central Adriatic Sea (Croatia). J. Mar. Sci. Eng. 2021, 9, 358. [Google Scholar] [CrossRef]
- Lattos, A.; Papadopoulos, D.K.; Feidantsis, K.; Karagiannis, D.; Giantsis, I.A.; Michaelidis, B. Are marine heatwaves responsible for mortalities of farmed Mytilus galloprovincialis? A pathophysiological analysis of Marteilia infected mussels from Thermaikos Gulf, Greece. Animals 2022, 12, 2805. [Google Scholar] [CrossRef] [PubMed]
- Mandić, M.; Nikolić, S.; Kokić, I.; Jokanović, S. Mass mortality of farmed mussels—A phenomenon without explanation? Studia Marina 2024, 37, 5–21. [Google Scholar]
- Marini, M.; Grilli, F. The Role of Nitrogen and Phosphorus in Eutrophication of the Northern Adriatic Sea: History and Future Scenarios. Appl. Sci. 2023, 13, 9267. [Google Scholar] [CrossRef]
- Cozzi, S.; Giani, M. River water and nutrient discharges in the Northern Adriatic Sea: Current importance and long term changes. Cont. Shelf Res. 2011, 31, 1881–1893. [Google Scholar] [CrossRef]
- Juza, M.; Fernández-Mora, À.; Tintoré, J. Sub-Regional Marine Heat Waves in the Mediterranean Sea from Observations: Long-Term Surface Changes, Sub-Surface and Coastal Responses. Front. Mar. Sci. 2022, 9, 785771. [Google Scholar] [CrossRef]
- Harley, C.D. Tidal dynamics, topographic orientation, and temperature-mediated mass mortalities on rocky shores. Mar. Ecol. Prog. Ser. 2008, 371, 37–46. [Google Scholar] [CrossRef]
- Anestis, A.; Lazou, A.; Portner, H.O.; Michaelidis, B. Behavioral, metabolic, and molecular stress responses of marine bivalve Mytilus galloprovincialis during long-term acclimation at increasing ambient temperature. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R911–R921. [Google Scholar] [CrossRef]
- Hofmann, G.E.; Somero, G.N. Interspecific variation in thermal denaturation of proteins in the congeneric mussels Mytilus trossulus and M. galloprovincialis: Evidence from the heat-shock response and protein ubiquitination. Mar. Biol. 1996, 126, 65–75. [Google Scholar] [CrossRef]
- Fly, E.K.; Hilbish, T.J.; Wethey, D.S.; Rognstad, R.L. Physiology and Biogeography: The Response of European Mussels (Mytilus spp.) to Climate Change. Am. Malacol. Bull. 2015, 33, 136–149. [Google Scholar] [CrossRef]
- Grimmelpont, M.; Payton, L.; Lefrançois, C.; Tran, D. Molecular and behavioural responses of the mussel Mytilus edulis exposed to a marine heatwave. Mar. Environ. Res. 2024, 196, 106418. [Google Scholar] [CrossRef]
- De Marco, A.; Baldassarro, V.A.; Calzà, L.; Giardino, L.; Dondi, F.; Ferrari, M.G.; Bignami, G.; Parma, L.; Bonaldo, A. Prolonged heat waves reduce the condition index and alter the molecular parameters in the pacific oyster Crassostrea gigas. Fish Shellfish Immunol. 2023, 133, 108518. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Z.; Zhang, Y.; Liang, J.; He, G.; Liu, X.; Zhao, L. Transcriptome analysis reveals acclimation responses of pearl oysters to marine heatwaves. Sci. Total Environ. 2022, 810, 151189. [Google Scholar] [CrossRef]
- Fabbri, E.; Valbonesi, P.; Franzellitti, S. HSPexpression in bivalves. Invertebr. Surviv. J. 2008, 5, 135–161. [Google Scholar]
- WMO (World Meteorological Organization). Guide to Meteorological Instruments and Methods of Observation. 2008. Available online: https://www.weather.gov/media/epz/mesonet/CWOP-WMO8.pdf (accessed on 12 February 2025).
- SNPA. Il Clima in Italia Nel 2023, Report Ambientali SNPA, n. 42/2024. Available online: https://www.puntosicuro.it/_resources/files/Rapporto-SNPA-clima-2023.pdf (accessed on 12 February 2025).
- Pastor, F.; Valiente, J.A.; Khodayar, S. A Warming Mediterranean: 38 Years of Increasing Sea Surface Temperature. Remote Sens. 2020, 12, 2687. [Google Scholar] [CrossRef]
- Qualità delle Acque della Costa. Available online: https://www.uniurb.it/ricerca/organizzazione-della-ricerca/strutture-della-ricerca/qualita-delle-acque-della-costa (accessed on 6 December 2024).
- Murray, S.N.; Ambrose, R.F.; Dethier, M.N. Methods for Performing Monitoring, Impact, and Ecological Studies on Rocky Shores; MMS U.S. Department of the Interior Minerals Management Service: Pacific OCS Region, USA, 2002; pp. 116–147.
- Parsell, D.A.; Lindquist, S. The function of heat-shock proteins in stress tolerance: Degradation and reactivation of damaged proteins. Annu. Rev. Genet. 1993, 27, 437–497. [Google Scholar] [CrossRef]
- Fourie, A.M.; Hupp, T.R.; Lane, D.P.; Sang, B.C.; Barbosa, M.S.; Sambrook, J.F.; Gething, M.J.H. HSP70 binding sites in the tumor suppressor protein p53. J. Biol. Chem. 1997, 272, 19471–19479. [Google Scholar] [CrossRef]
- Earnshaw, W.C.; Martins, L.M.; Kaufmann, S.H. Mammalian caspases: Structure, activation, substrates, and functions during apoptosis. Annu. Rev. Biochem. 1999, 68, 383–424. [Google Scholar] [CrossRef]
- Meng, X.; Li, F.; Wang, X.; Liu, J.; Ji, C.; Wu, H. Toxicological effects of graphene on mussel Mytilus galloprovincialis hemocytes after individual and combined exposure with triphenyl phosphate. Mar. Pollut. Bull. 2020, 151, 110838. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Borghi, C.; Fabbri, R.; Ciacci, C.; Lorusso, L.C.; Gallo, G.; Vergani, L. Effects of 17β-estradiol on mussel digestive gland. Gen. Comp. Endocrinol. 2007, 153, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Banni, M.; Sforzini, S.; Franzellitti, S.; Oliveri, C.; Viarengo, A.; Fabbri, E. Molecular and cellular effects induced in Mytilus galloprovincialis treated with oxytetracycline at different temperatures. PLoS ONE 2015, 10, e0128468. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ji, C.; Wei, L.; Zhao, J.; Lu, H. Proteomic and metabolomic responses in hepatopancreas of Mytilus galloprovincialis challenged by Micrococcus luteus and Vibrio anguillarum. J. Proteom. 2013, 94, 54–67. [Google Scholar] [CrossRef]
- Laurent, L.; Buoncristiani, J.-F.; Pohl, B.; Zekollari, H.; Farinotti, D.; Huss, M.; Mugnier, J.-L.; Pergaud, J. The impact of climate change and glacier mass loss on the hydrology in the Mont-Blanc massif. Sci. Rep. 2020, 10, 10420. [Google Scholar] [CrossRef]
- Fazzini, M.; Baione, L.; Raspanti, A.; Capizzi, P.; Casagli, N. Climate crisis and influence on snowfall in the Italian physical territory in the last thirty years—CLINO 1991–2020. In Proceedings of the 6th World Landslide Forum. Landslide Science for Sustainable Development, Florence, Italy, 14–17 November 2023; ISBN 9791221048063. [Google Scholar]
- Rosa, M.; Capriotti, M.; Austin, K.; Shumway, S.E.; Ward, J.E. Effect of seasonal changes in temperature on capture efficiency in the blue mussel, Mytilus edulis, fed seston and microplastics. Invertebr. Biol. 2024, 143, e12446. [Google Scholar] [CrossRef]
- Bracchetti, L.; Capriotti, M. Le formazioni a Reef della Costa Picena. Studi Costieri. I Litorali Marchig. E Il Nuovo Piano di Gestione Integr. Delle Zone Costiere 2021, 29, 83. [Google Scholar]
- Masanja, F.; Xu, Y.; He, G.; Liang, F.; Liu, X.; Yang, K.; Zhao, L. Exploring HSP90 as a biomarker for marine heatwaves in pinctada maxima. Front. Mar. Sci. 2022, 9, 913920. [Google Scholar] [CrossRef]
- Carella, F.; Aceto, S.; Mangoni, O.; Mollica, M.P.; Cavaliere, G.; Trinchese, G.; Aniello, F.; De Vico, G. Assessment of the health status of mussels Mytilus galloprovincialis along the Campania coastal areas: A multidisciplinary approach. Front. Physiol. 2018, 9, 683. [Google Scholar] [CrossRef]
- Lakin, N.D.; Jackson, S.P. Regulation of p53 in response to DNA damage. Oncogene 1999, 18, 7644–7655. [Google Scholar] [CrossRef]
- Cocci, P.; Capriotti, M.; Mosconi, G.; Palermo, F.A. Transcriptional variations in biomarkers of Mytilus galloprovincialis sampled from Central Adriatic coastal waters (Marche region, Italy). Biomarkers 2017, 22, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xu, L.; Xiong, X.; Liu, X. Expansion and diversity of caspases in Mytilus coruscus contribute to larval metamorphosis and environmental adaptation. BMC Genom. 2024, 25, 314. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xu, L.; Wang, Y.; Lin, S.; Li, H.; Qi, P.; Buttino, I.; Wang, W.; Guo, B. Insights into the Response in Digestive Gland of Mytilus coruscus under Heat Stress Using TMT-Based Proteomics. Animals 2023, 13, 2248. [Google Scholar] [CrossRef] [PubMed]
- Peruzza, L.; Tucci, C.F.; Frizzo, R.; Riello, T.; Quagliariello, A.; Martino, M.E.; Manuzzi, A.; Rovere, G.D.; Bonsembiante, F.; Gelain, M.E.; et al. Impaired reproduction, energy reserves and dysbiosis: The overlooked consequences of heatwaves in a bivalve mollusc. Mar. Pollut. Bull. 2023, 193, 115192. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence Forward | Primer Sequence Reverse | Reference |
|---|---|---|---|
| Caspase 3 | GATCTTGGAAGTGGTGTAGAACG | CACTGCTAGGAAATCTGCTTCAT | [35] |
| p53 | CCAACTTGCTAAATTTGTTGAAGA | TTGGTCCTCCTACACATGAC | [36] |
| HSP70 | CCGTCCACACCACCCACC | GTGAGGTTAGCTGACAATGGTGG | [37] |
| HSP90 | AAGCTGATCTGGTCAATAACCTGG | AACCTACACCAAACTGTCCAATCA | [37] |
| 18S rRNA | AGAAACGGCTACCACATCC | TGCCCTCCAATAGATCCTC | [38] |
| Site | February | March | April | June | July | August | September |
|---|---|---|---|---|---|---|---|
| TRN | 90 | 90 | 95 | 95 | 75 | 20 | 0 |
| FSN | 50 | 95 | 95 | 85 | 70 | 5 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capriotti, M.; Bracchetti, L.; Cocci, P.; Mazzocchi, V.; Fazzini, M.; Corvaro, F.; Palermo, F.A. In Situ Investigation of Ecological and Molecular Stress Mechanisms Triggered by Marine Heat Waves in Adriatic Populations of the Mediterranean Mussel. Diversity 2025, 17, 385. https://doi.org/10.3390/d17060385
Capriotti M, Bracchetti L, Cocci P, Mazzocchi V, Fazzini M, Corvaro F, Palermo FA. In Situ Investigation of Ecological and Molecular Stress Mechanisms Triggered by Marine Heat Waves in Adriatic Populations of the Mediterranean Mussel. Diversity. 2025; 17(6):385. https://doi.org/10.3390/d17060385
Chicago/Turabian StyleCapriotti, Martina, Luca Bracchetti, Paolo Cocci, Valentina Mazzocchi, Massimiliano Fazzini, Francesco Corvaro, and Francesco Alessandro Palermo. 2025. "In Situ Investigation of Ecological and Molecular Stress Mechanisms Triggered by Marine Heat Waves in Adriatic Populations of the Mediterranean Mussel" Diversity 17, no. 6: 385. https://doi.org/10.3390/d17060385
APA StyleCapriotti, M., Bracchetti, L., Cocci, P., Mazzocchi, V., Fazzini, M., Corvaro, F., & Palermo, F. A. (2025). In Situ Investigation of Ecological and Molecular Stress Mechanisms Triggered by Marine Heat Waves in Adriatic Populations of the Mediterranean Mussel. Diversity, 17(6), 385. https://doi.org/10.3390/d17060385