
Correlation Between Individual Body Condition and Seasonal Activity in Buresch’s Crested Newt, Triturus ivanbureschi


Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Population Statistics
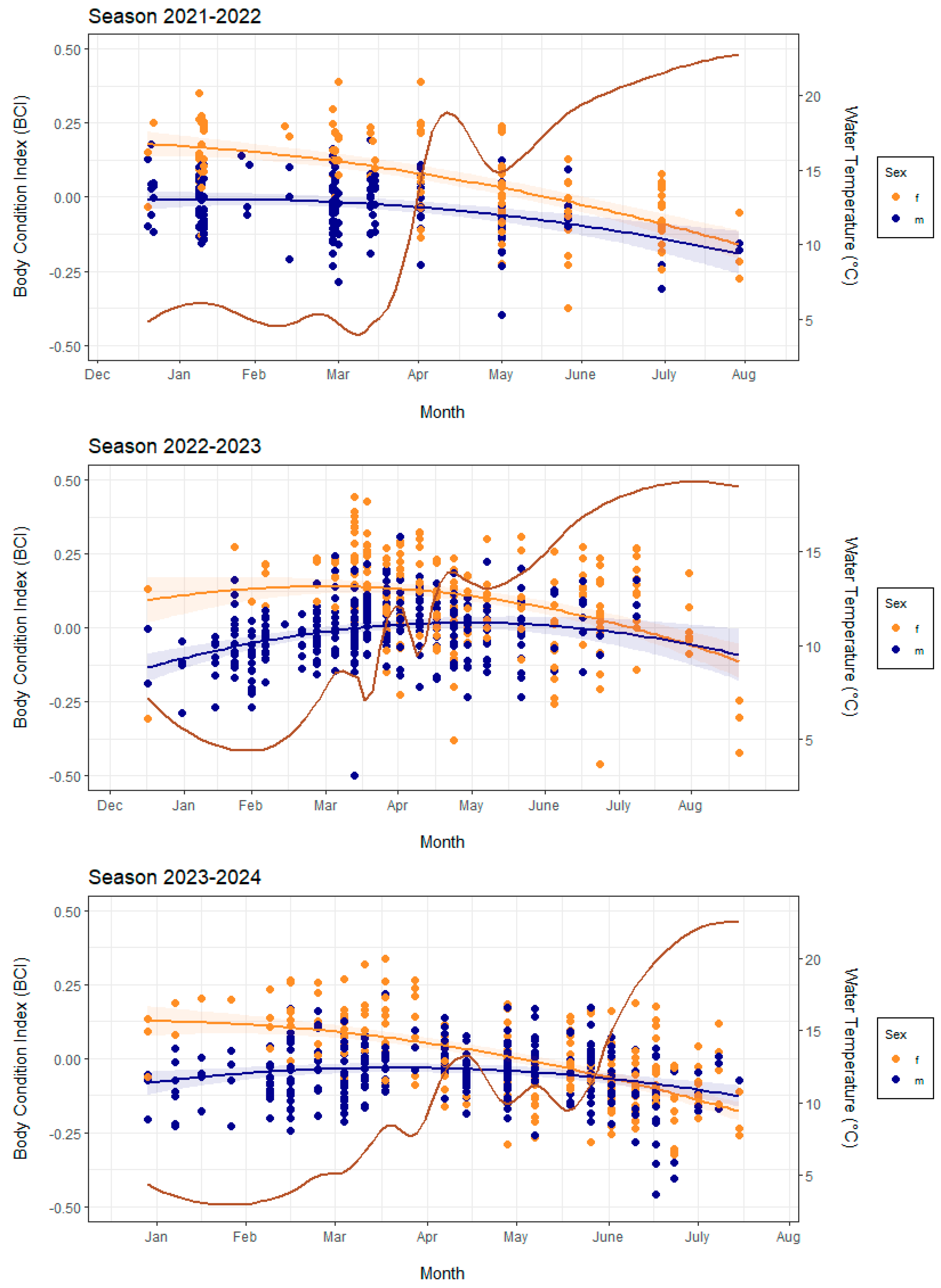
3.2. BCI Change
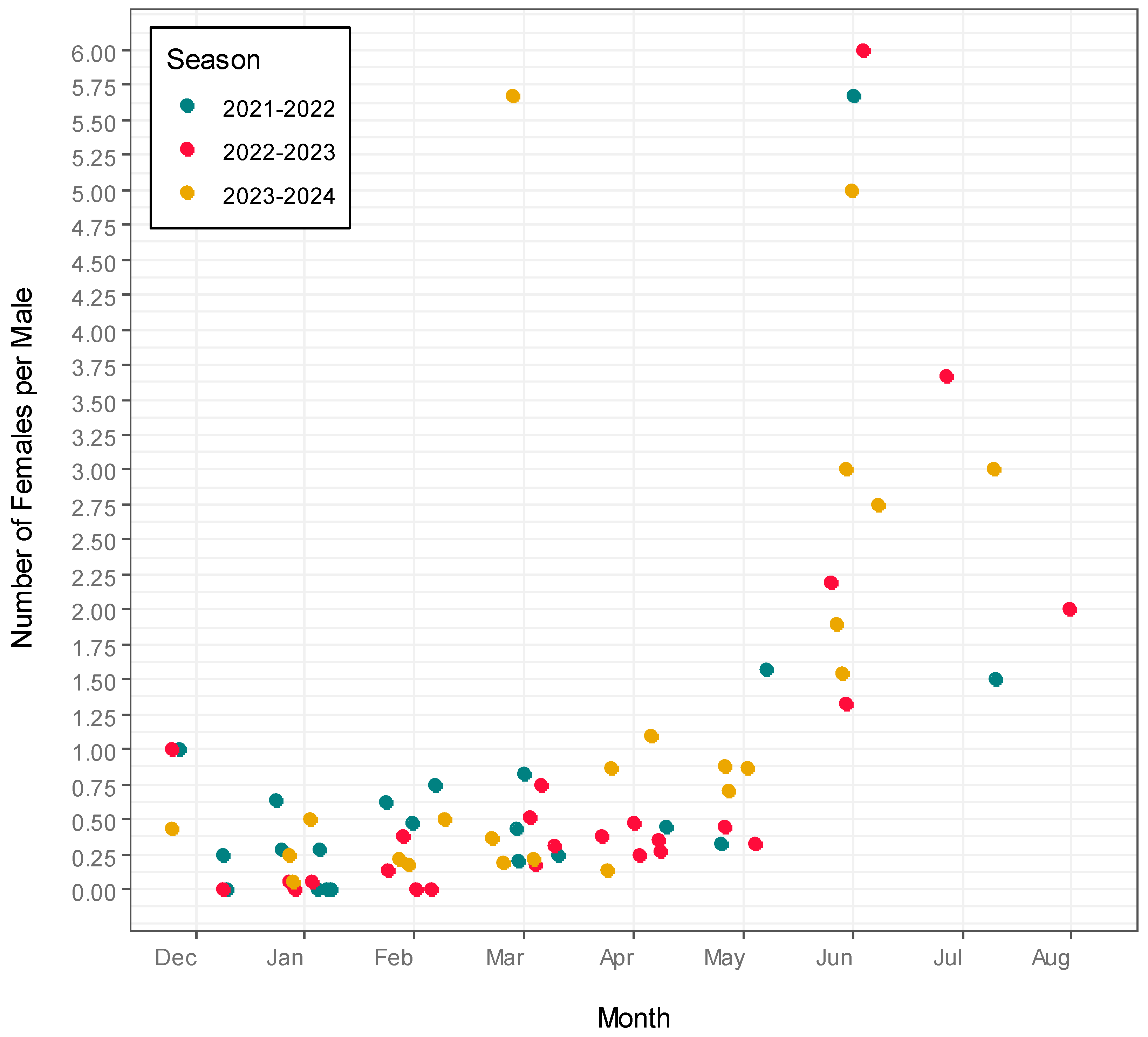
3.3. Population Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- The IUCN Red List of Threatened Species. Version 2024-3. Available online: https://www.iucnredlist.org (accessed on 21 February 2025).
- Üveges, B.; Mahr, K.; Szederkényi, M.; Bókony, V.; Hoi, H.; Hettyey, A. Experimental evidence for beneficial effects of projected climate change on hibernating amphibians. Sci. Rep. 2016, 6, 26754. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Thuiller, W.; Pearson, R.G. Climate warming and the decline of amphibians and reptiles in Europe. J. Biogeogr. 2006, 33, 1712–1728. [Google Scholar] [CrossRef]
- Benard, M.F. Warmer winters reduce frog fecundity and shift breeding phenology, which consequently alters larval development and metamorphic timing. Glob. Change Biol. 2015, 21, 1058–1065. [Google Scholar] [CrossRef]
- Hayes, J.P.; Shonkwiler, J.S. Morphometric indicators of body condition: Worthwhile or wishful thinking? In Body Composition Analysis of Animals: A Handbook of Non-Destructive Methods; Speakman, J.R., Ed.; Cambridge University Press: Cambridge, UK, 2001; pp. 8–38. [Google Scholar]
- Stevenson, R.D.; Woods, W.A. Condition indices for conservation: New uses for evolving tools. Integr. Comp. Biol. 2006, 46, 1169–1190. [Google Scholar] [CrossRef]
- McCaffrey, K.R.; Balaguera-Reina, S.A.; Falk, B.G.; Gati, E.V.; Cole, J.M.; Mazzotti, F.J. How to estimate body condition in large lizards? Argentine black and white tegu (Salvator merianae, Dumeril and Bibron, 1839) as a case study. PLoS ONE 2023, 18, e0282093. [Google Scholar] [CrossRef] [PubMed]
- Jakob, E.M.; Marshall, S.D.; Uetz, G.W. Estimating fitness: A comparison of body condition indices. Oikos 1996, 77, 61–67. [Google Scholar] [CrossRef]
- Green, A.J. Mass/length residuals: Measures of body condition or generators of spurious results? Ecology 2001, 82, 1473–1483. [Google Scholar] [CrossRef]
- Labocha, M.K.; Schutz, H.; Hayes, J.P. Which body condition index is best? Oikos 2014, 123, 111–119. [Google Scholar] [CrossRef]
- Bucciarelli, G.M.; Clark, M.A.; Delaney, K.S.; Riley, S.P.D.; Shaffer, H.B.; Fisher, R.N.; Honeycutt, R.L.; Kats, L.B. Amphibian responses in the aftermath of extreme climate events. Sci. Rep. 2020, 10, 3409. [Google Scholar] [CrossRef]
- McCaffery, R.M.; Maxell, B.A. Decreased winter severity increases viability of a montane frog population. Proc. Natl. Acad. Sci. USA 2010, 107, 8644–8649. [Google Scholar] [CrossRef]
- Caruso, N.M.; Sears, M.W.; Adams, D.C.; Lips, K.R. Widespread rapid reductions in body size of adult salamanders in response to climate change. Glob. Change Biol. 2014, 20, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Lukanov, S.; Lazarkevich, I.; Dimitrova, B. Persistent winter activity in Triturus ivanbureschi Arntzen & Wielstra, 2013 (Amphibia: Caudata). Acta Zool. Bulg. 2022, 74, 281–285. [Google Scholar]
- Wielstra, B.; Litvinchuk, S.N.; Naumov, B.; Tzankov, N.; Arntzen, J.W. A revised taxonomy of crested newts in the Triturus karelinii group (Amphibia: Caudata: Salamandridae), with the description of a new species. Zootaxa 2013, 3682, 441–453. [Google Scholar] [CrossRef]
- Stojanov, A.; Tzankov, N.; Naumov, B. Die Amphibien und Reptilien Bulgariens; Chimaira: Frankfurt am Main, Germany, 2011; p. 582. [Google Scholar]
- Lovich, J.E.; Gibbons, J.W. A review of techniques for quantifying sexual size dimorphism. Growth Dev. Aging 1992, 56, 269–281. [Google Scholar]
- Crall, J.P.; Stewart, C.V.; Berger-Wolf, T.Y.; Rubenstein, D.I.; Sundaresan, S.R. HotSpotter—Patterned species instance recognition. In Proceedings of the 2013 IEEE Workshop on Applications of Computer Vision (WACV), Clearwater Beach, FL, USA, 15–17 January 2013; pp. 230–237. [Google Scholar] [CrossRef]
- Naumov, B.; Lukanov, S. Notes on age-related changes in body size and color pattern in captive Triturus dobrogicus (Kiritzescu, 1903). Herpetozoa 2013, 30, 159–168. [Google Scholar]
- Lukanov, S. Inter-pond migration during the aquatic phase by male Triturus ivanbureschi. Russ. J. Herpetol. 2022, 29, 373–376. [Google Scholar] [CrossRef]
- Parris, K.; McCarthy, M. Identifying effects of toe clipping on anuran return rates: The importance of statistical power. Amphib-Reptilia 2001, 22, 275–289. [Google Scholar]
- Naumov, B.; Popgeorgiev, G.; Kornilev, Y.; Plachiyski, G.; Stojanov, A.; Tzankov, N. Distribution and ecology of the Alpine newt Ichthyosaura alpestris (Laurenti, 1768) (Amphibia: Salamandridae) in Bulgaria. Acta Zool. Bulg. 2020, 72, 83–102. [Google Scholar]
- Schwarz, C.J.; Arnason, A.N. A general methodology for the analysis of open-model capture recapture experiments. Biometrics 1996, 52, 860–873. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 10 October 2024).
- Băncilă, R.I.; Hartel, T.; Plăiaşu, R.; Cogălniceanu, D.; Smets, J. Comparing three body condition indices in amphibians: A case study of yellow-bellied toad Bombina variegata. Amphib-Reptilia 2010, 31, 558–562. [Google Scholar]
- Jarvis, L.E. Terrestrial ecology of juvenile great crested newts (Triturus cristatus) in a woodland area. Herpetol. J. 2016, 26, 287–296. [Google Scholar]
- Lukanov, S.; Tzankov, N. Life history, age and normal development of the Balkan-Anatolian crested newt (Triturus ivanbureschi Arntzen and Wielstra, 2013) from Sofia district. North-West. J. Zool. 2016, 12, 22–32. [Google Scholar]
- Jarvis, L.E. Factors affecting body condition in a great crested newt Triturus cristatus population. Herpetol. Bull. 2015, 134, 1–5. [Google Scholar]
- Berzin, D.L.; Burakova, A.V. Diet features of the Crested newt Triturus cristatus (Laurenti, 1768) at the eastern border of its range. Russ. J. Ecol. 2022, 53, 221–227. [Google Scholar] [CrossRef]
- Naumov, B.; Vacheva, E.; Lukanov, S. Diet and feeding habits of Triturus cristatus (Laurenti, 1768) (Amphibia: Salamandridae) in its southernmost locality. North-West. J. Zool. 2024, 20, 116–125. [Google Scholar]
- Hedlund, L. Factors affecting differential mating success in male crested newts, Triturus cristatus. J. Zool. 1990, 220, 33–40. [Google Scholar] [CrossRef]
- Green, A.J. Large male crests, an honest indicator of condition, are preferred by female smooth, newts, Triturus vulgaris (Salamandridae) at the spermatophore transfer stage. Anim. Behav. 1991, 41, 367–369. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Smithson, A.; Oldham, R.S. Marking and tissue sampling effects on body condition and survival in the newt Triturus cristatus. J. Herpetol. 1999, 33, 567. [Google Scholar] [CrossRef]
- Halliday, T.; Tejedo, M. Intrasexual selection and alternative mating behaviour. In Amphibian Biology; Heatwole, H., Sullivan, B.K., Eds.; Surrey Beatty & Sons Pty Ltd.: Baulkham Hills, Australia, 1995; Volume 2, pp. 419–468. [Google Scholar]
- Brzeziński, M.; Eliava, G.; Żmihorski, M. Road mortality of pond-breeding amphibians during spring migrations in the Mazurian Lakeland, NE Poland. Eur. J. Wildl. Res. 2012, 58, 685–693. [Google Scholar] [CrossRef]
- Griffiths, R.A.; Sewell, D.; McCrea, R.S. Dynamics of a declining amphibian metapopulation: Survival, dispersal and the impact of climate. Biol. Conserv. 2010, 143, 485–491. [Google Scholar] [CrossRef]
- Jørgensen, C.B. External and internal control of patterns of feeding, growth and gonadal function in a temperate zone anuran, the toad Bufo bufo. J. Zool. 1986, 210, 211–241. [Google Scholar] [CrossRef]
- Lukanov, S.; Doncheva, T.; Kostova, N.; Naumov, B. Effects of selected environmental parameters on the activity and body condition of the Buresch’s crested newt (Triturus ivanbureschi) with notes on skin secretions. North-West. J. Zool. 2021, 17, 34–38. [Google Scholar]
- Ockleford, C.; Adriaanse, P.; Berny, P.; Brock, T.; Duquesne, S.; Grilli, S.; Hernandez-Jerez, A.F.; Bennekou, S.H.; Klein, M.; Kuhl, T.; et al. Scientific Opinion on the state of the science on pesticide risk assessment for amphibians and reptiles. EFSA J. 2018, 16, e05125. [Google Scholar] [CrossRef]
- Hailey, A.; Willemsen, R.E. Population density and adult sex ratio of the tortoise Testudo hermanni in Greece: Evidence for intrinsic population regulation. J. Zool. 2000, 251, 325–338. [Google Scholar] [CrossRef]
- Pickett, E.J.; Stockwell, M.P.; Pollard, C.J.; Garnham, J.I.; Clulow, J.; Mahony, M.J. Estimates of sex ratio require the incorporation of unequal catchability between sexes. Wildl. Res. 2012, 39, 350–354. [Google Scholar] [CrossRef]
- Berven, K.A. Factors affecting population fluctuations in larval and adult stages of the wood frog (Rana sylvatica). Ecology 1990, 71, 1599–1608. [Google Scholar] [CrossRef]
- Anholt, B.R.; Hotz, H.; Guex, G.D.; Semlitsch, R.D. Overwinter survival of Rana lessonae and its hemiclonal associate Rana esculenta. Ecology 2003, 84, 391–397. [Google Scholar] [CrossRef]
- Amrhein, V.; Scaar, B.; Baumann, M.; Minéry, N.; Binnert, J.P.; Korner-Nievergelt, F. Estimating adult sex ratios from bird mist netting data. Methods Ecol. Evol. 2012, 3, 713–720. [Google Scholar] [CrossRef]
- McKnight, D.T.; Ligon, D.B. Correcting for unequal catchability in sex ratio and population size estimates. PLoS ONE 2017, 12, e0184101. [Google Scholar] [CrossRef]
- Denoël, M.; Dalleur, S.; Langrand, E.; Besnard, A.; Cayuela, H. Dispersal and alternative breeding site fidelity strategies in an amphibian. Ecography 2018, 41, 1543–1555. [Google Scholar] [CrossRef]
- Cayuela, H.; Schmidt, B.R.; Weinbach, A.; Besnard, A.; Joly, P. Multiple density-dependent processes shape the dynamics of a spatially structured amphibian population. J. Anim. Ecol. 2019, 88, 164–177. [Google Scholar] [CrossRef]
- Arntzen, J. Seasonal variation in sex ratio and asynchronous presence at ponds of male and female Triturus newts. J. Herpetol. 2002, 36, 30–35. [Google Scholar] [CrossRef]
- Lunghi, E.; Bruni, G. Long-term reliability of Visual Implant Elastomers in the Italian cave salamander (Hydromantes italicus). Salamandra 2018, 54, 283–286. [Google Scholar]
- Kafash, A.; Ashrafi, S.; Ohler, A.; Yousefi, M.; Malakoutikhah, S.; Koehler, G.; Schmidt, B.R. Climate change produces winners and losers: Differential responses of amphibians in mountain forests of the Near East. Glob. Ecol. Conserv. 2018, 16, e00471. [Google Scholar] [CrossRef]
- Moldowan, P.D.; Tattersall, G.J.; Rollinson, N. Climate-associated decline of body condition in a fossorial salamander. Glob. Change Biol. 2022, 28, 1725–1739. [Google Scholar] [CrossRef]
- Lannoo, M.; Stiles, R. Effects of short-term climate variation on a long-lived frog. Copeia 2017, 105, 726–733. [Google Scholar] [CrossRef]
- Cayuela, H.; Arsovski, D.; Bonnaire, E.; Duguet, R.; Joly, P.; Besnard, A. The impact of severe drought on survival, fecundity, and population persistence in an endangered amphibian. Ecosphere 2016, 7, e01246. [Google Scholar] [CrossRef]
- Dodd, C.K. Cost of living in an unpredictable environment: The ecology of Striped Newts Notophthalmus perstriatus during a prolonged drought. Copeia 1993, 1993, 605–614. [Google Scholar] [CrossRef]
- Fahrbach, M.; Gerlach, U. The Genus Triturus: History, Biology, Systematics, Captive Breeding; Chimaira Verlag: Frankfurt, Germany, 2018; 550p. [Google Scholar]
- Bickford, D.; Howard, S.D.; Ng, D.J.J.; Sheridan, J.A. Impacts of climate change on the amphibians and reptiles of Southeast Asia. Biodivers. Conser. 2010, 19, 1043–1062. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2021–2022 | 2022–2023 | 2023–2024 | ||||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |
| TC | 270 | 138 | 482 | 232 | 394 | 249 |
| R | 143 | 50 | 245 | 99 | 242 | 92 |
| NR | 127 | 88 | 237 | 133 | 152 | 157 |
| RR | 52.96% | 36.23% | 50.83% | 42.67% | 61.42% | 36.95% |
| SVL | 7.6 ± 0.6 (5.8–8.7) | 7.8 ± 0.5 (6.8–9.0) | 7.5 ± 0.4 (6.4–8.8) | 7.8 ± 0.5 (5.7–9.2) | 7.6 ± 0.4 (6.7–9.0) | 8.0 ± 0.6 (6.0–10.0) |
| Mass | 12.2 ± 2.5 (6.4–19.2) | 14.2 ± 2.9 (8.1–22.9) | 12.3 ± 2.2 (7.9–18.8) | 15.0 ± 3.1 (6.7–24.8) | 12.2 ± 2.1 (7.3–19.0) | 14.2 ± 2.6 (8.4–22.4) |
| SDI | −0.03 | −0.04 | −0.05 | |||
| N | 254 ± 15.4 (226–286) | 195 ± 21.9 (156–243) | 376 ± 17.5 (343–412) | 259 ± 22.8 (218–308) | 353 ± 33.9 (292–426) | 233 ± 13.6 (207–261) |
| p | 0.092 ± 0.007 (0.080–0.112) | 0.061 ± 0.009 (0.053–0.080) | 0.311 ± 0.017 (0.272–0.350) | 0.111 ± 0.013 (0.082–0.131) | 0.200 ± 0.021 (0.171–0.242) | 0.101 ± 0.011 (0.081–0.132) |
| b | 0.040 ± 0.003 (0.031–0.053) | 0.051 ± 0.003 (0.412–0.062) | 0.040 ± 0.001 (0.039–0.042) | 0.040 ± 0.001 (0.039–0.041) | 0.043 ± 0.001 (0.041–0.046) | 0.038 ± 0.002 (0.033–0.043) |
| Phi | 0.921 ± 0.014 (0.881–0.942) | 0.901 ± 0.010 (0.860–0.931) | 0.941 ± 0.015 (0.891–0.953) | 0.831 ± 0.012 (0.801–0.850) | 0.910 ± 0.021 (0.861–0.941) | 0.911 ± 0.012 (0.881–0.931) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukanov, S.; Atanasova, I. Correlation Between Individual Body Condition and Seasonal Activity in Buresch’s Crested Newt, Triturus ivanbureschi. Diversity 2025, 17, 350. https://doi.org/10.3390/d17050350
Lukanov S, Atanasova I. Correlation Between Individual Body Condition and Seasonal Activity in Buresch’s Crested Newt, Triturus ivanbureschi. Diversity. 2025; 17(5):350. https://doi.org/10.3390/d17050350
Chicago/Turabian StyleLukanov, Simeon, and Irena Atanasova. 2025. "Correlation Between Individual Body Condition and Seasonal Activity in Buresch’s Crested Newt, Triturus ivanbureschi" Diversity 17, no. 5: 350. https://doi.org/10.3390/d17050350
APA StyleLukanov, S., & Atanasova, I. (2025). Correlation Between Individual Body Condition and Seasonal Activity in Buresch’s Crested Newt, Triturus ivanbureschi. Diversity, 17(5), 350. https://doi.org/10.3390/d17050350