
Response of Bird Communities to Human-Dominated Habitats in Southern Anhui Mountainous Area, China

Abstract
1. Introduction
2. Materials and Methods
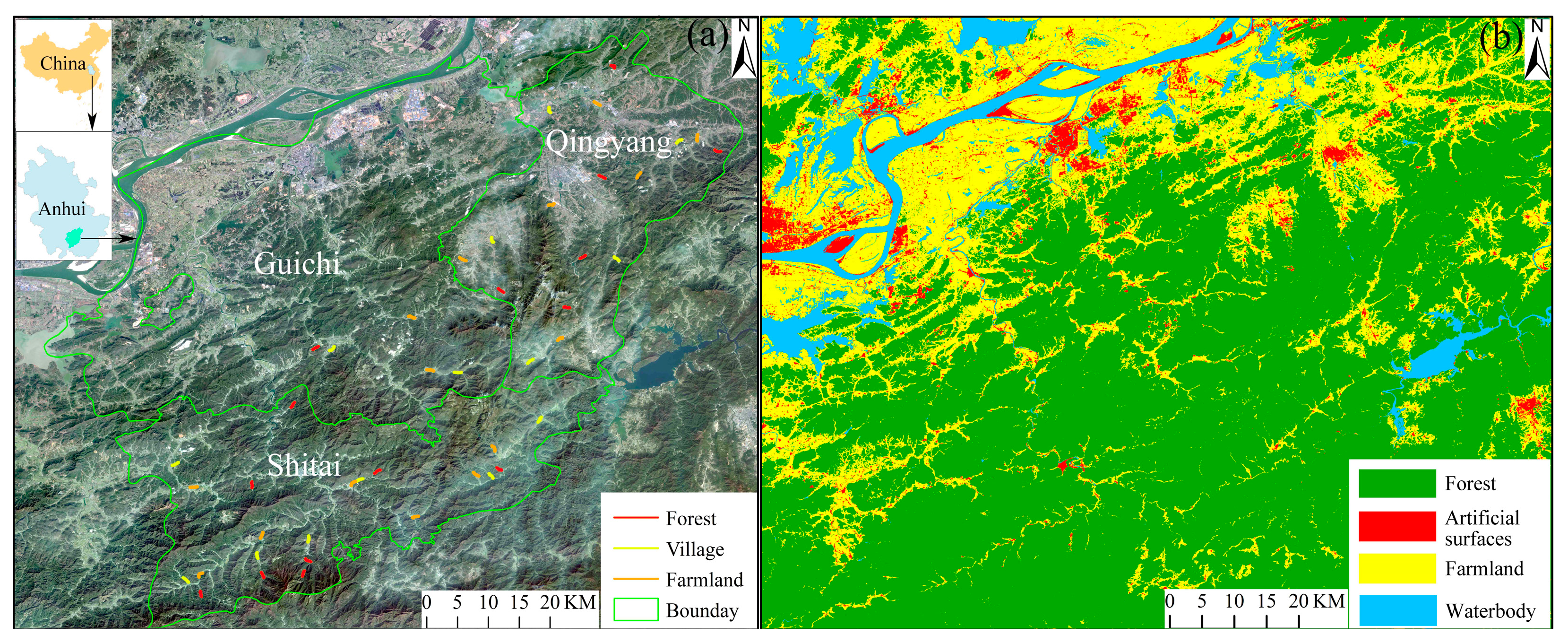
2.1. Study Area
2.2. Bird Surveys
2.3. Function Traits and Phylogenies Data
2.4. Diversity Metrics
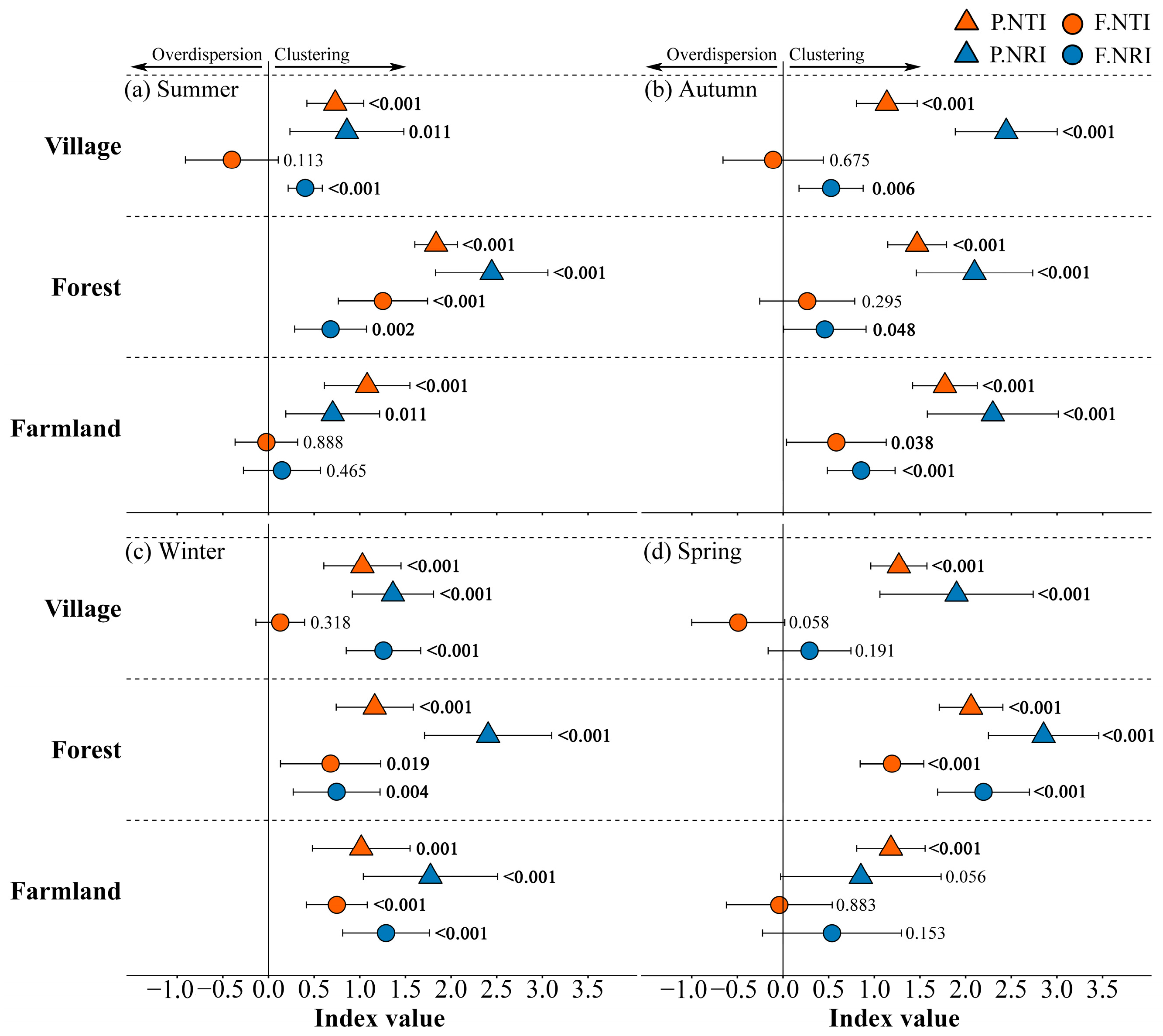
2.5. Assembly Process and Null Models
2.6. Data Analyses
3. Results
3.1. Birds Communities
3.2. Phylogenetic Signal of Bird Functional Traits
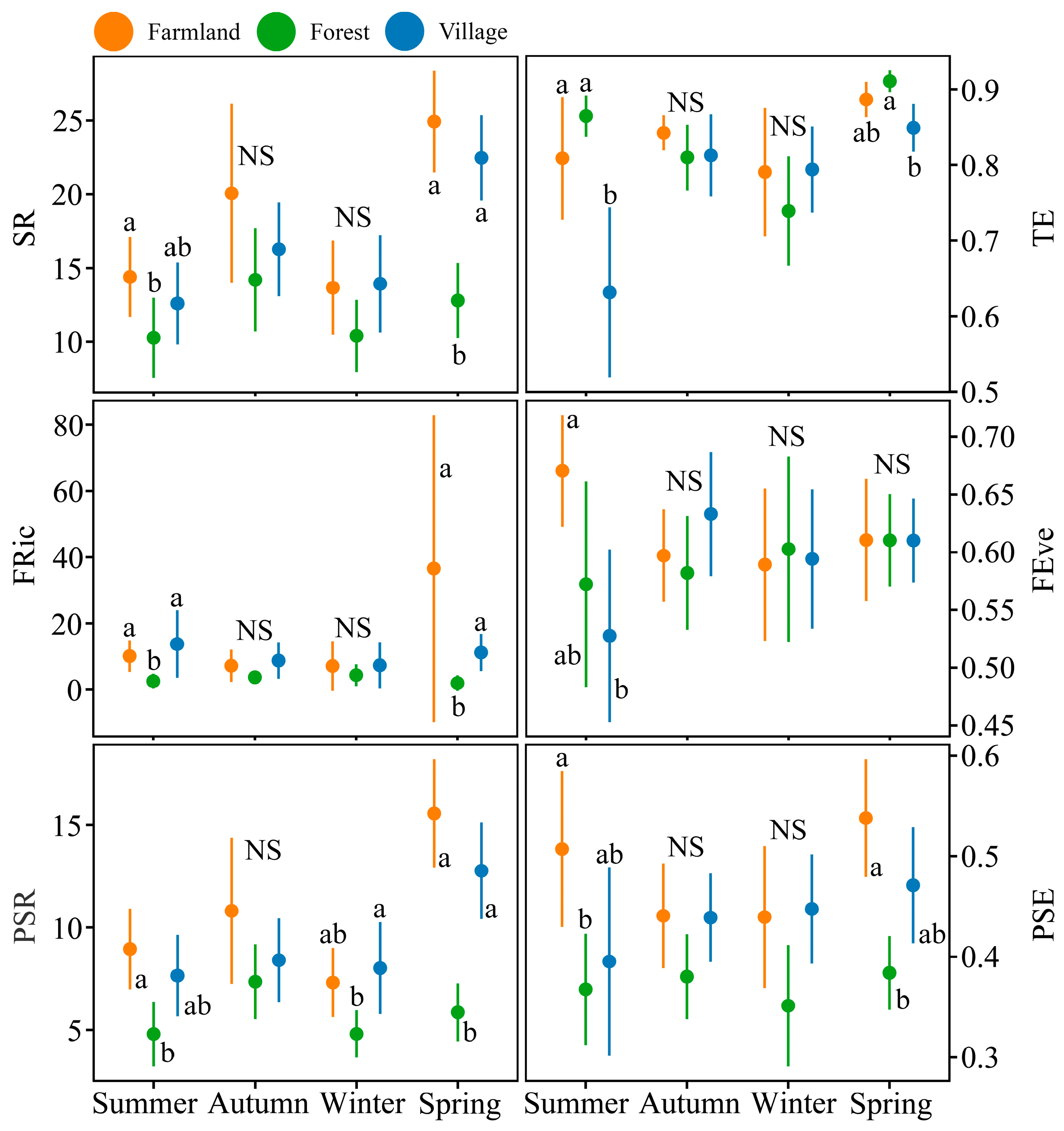
3.3. Multidimensional Diversity Analysis of Bird Communities
3.4. Standardized Functional and Phylogenetic Diversity
3.5. Effects of Land Use Type and Season on Bird Body Mass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yang, C.; Li, Q.; Wang, X.; Cui, A.; Chen, J.; Liu, H.; Ma, W.; Dong, X.; Shi, T.; Meng, F. Human expansion-induced biodiversity crisis over Asia from 2000 to 2020. Research 2023, 6, 0226. [Google Scholar] [PubMed]
- Crist, E.; Mora, C.; Engelman, R. The interaction of human population, food production, and biodiversity protection. Science 2017, 356, 260–264. [Google Scholar] [PubMed]
- Simkin, R.D.; Seto, K.C.; McDonald, R.I.; Jetz, W. Biodiversity impacts and conservation implications of urban land expansion projected to 2050. Proc. Natl. Acad. Sci. USA 2022, 119, e2117297119. [Google Scholar] [PubMed]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 2016, 7, 12558. [Google Scholar]
- Mayor, S.; Cahill, J., Jr.; He, F.; Sólymos, P.; Boutin, S. Regional boreal biodiversity peaks at intermediate human disturbance. Nat. Commun. 2012, 3, 1142. [Google Scholar]
- Connell, J.H. Diversity in tropical rain forests and coral reefs: High diversity of trees and corals is maintained only in a nonequilibrium state. Science 1978, 199, 1302–1310. [Google Scholar]
- Wilkinson, D.M. The disturbing history of intermediate disturbance. Oikos 1999, 84, 145–147. [Google Scholar]
- Cadotte, M.W. Competition–colonization trade-offs and disturbance effects at multiple scales. Ecology 2007, 88, 823–829. [Google Scholar]
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar]
- Huston, M.A. Disturbance, productivity, and species diversity: Empiricism vs. logic in ecological theory. Ecology 2014, 95, 2382–2396. [Google Scholar]
- Zhao, Y.; Dunn, R.R.; Zhou, H.; Si, X.; Ding, P. Island area, not isolation, drives taxonomic, phylogenetic and functional diversity of ants on land-bridge islands. J. Biogeogr. 2020, 47, 1627–1637. [Google Scholar]
- Wang, X.; Zhu, G.; Ma, H.; Wu, Y.; Zhang, W.; Zhang, Y.; Li, C.; de Boer, W.F. Bird communities’ responses to human-modified landscapes in the southern Anhui Mountainous Area. Avian Res. 2022, 13, 100006. [Google Scholar]
- Li, C.; Zhang, Y.; Zha, D.; Yang, S.; Huang, Z.Y.X.; de Boer, W.F. Assembly processes of waterbird communities across subsidence wetlands in China: A functional and phylogenetic approach. Divers. Distrib. 2019, 25, 1118–1129. [Google Scholar]
- MacArthur, R.; Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar]
- Tobias, J.A.; Ottenburghs, J.; Pigot, A.L. Avian diversity: Speciation, macroevolution, and ecological function. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 533–560. [Google Scholar]
- Gregory, R.D.; Noble, D.; Field, R.; Marchant, J.; Raven, M.; Gibbons, D. Using birds as indicators of biodiversity. Ornis Hung. 2003, 12, 11–24. [Google Scholar]
- Şekercioğlu, Ç.H.; Daily, G.C.; Ehrlich, P.R. Ecosystem consequences of bird declines. Proc. Natl. Acad. Sci. USA 2004, 101, 18042–18047. [Google Scholar]
- García, D.; Miñarro, M.; Martínez-Sastre, R. Birds as suppliers of pest control in cider apple orchards: Avian biodiversity drivers and insectivory effect. Agric. Ecosyst. Environ. 2018, 254, 233–243. [Google Scholar]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, X.; Song, W.; Li, Q.; Onditi, K.; Khanal, L.; Jiang, X. Small mammal species richness and turnover along elevational gradient in Yulong Mountain, Yunnan, Southwest China. Ecol. Evol. 2020, 10, 2545–2558. [Google Scholar] [CrossRef] [PubMed]
- Matthews, T.J.; Triantis, K.A.; Wayman, J.P.; Martin, T.E.; Hume, J.P.; Cardoso, P.; Faurby, S.; Mendenhall, C.D.; Dufour, P.; Rigal, F. The global loss of avian functional and phylogenetic diversity from anthropogenic extinctions. Science 2024, 386, 55–60. [Google Scholar] [CrossRef]
- Penjor, U.; Jamtsho, R.; Sherub, S. Anthropogenic land-use change shapes bird diversity along the eastern Himalayan altitudinal gradient. J. Appl. Ecol. 2022, 59, 847–859. [Google Scholar] [CrossRef]
- Lin, S.; Qiao, X.; Geng, Y.; Fan, C.; Zhang, C.; Zhao, X.; von Gadow, K. Environmental filtering drives biodiversity–spatial stability relationships in a large temperate forest region. Funct. Ecol. 2023, 37, 1688–1702. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H.; Xu, Y.; Wang, S. Composition of ‘fast–slow’ traits drives avian community stability over North America. Funct. Ecol. 2021, 35, 2831–2840. [Google Scholar] [CrossRef]
- Wang, H.; Hu, J.; Mei, H. Present situation and promotion strategy of agricultural mechanization in hilly areas of Anhui Province. Mod. Agric. Sci. Technol. (In Chinese). 2021, 11, 159–162. (In Chinese) [Google Scholar]
- Zhang, L.; Wang, Z.; Du, P. Construction of Mountain Ecological Security Pattern Coupled with Service and Safety Evaluation: A Case Study of the Mountainous Area of Southern Anhui Province. J. China West Norm. Univ. 2025. Available online: https://link.cnki.net/urlid/51.1699.N.20250313.1702.002 (accessed on 23 March 2025). (In Chinese).
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Sutherland, W.J.; Newton, I.; Green, R. Bird Ecology and Conservation: A Handbook of Techniques; OUP Oxford: Oxford, UK, 2004. [Google Scholar]
- MacKinnon, J. Guide to the Birds of China; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- Ding, Z.; Feeley, K.J.; Wang, Y.; Pakeman, R.J.; Ding, P. Patterns of bird functional diversity on land-bridge island fragments. J. Anim. Ecol. 2013, 82, 781–790. [Google Scholar] [CrossRef]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar]
- Wang, Y.; Song, Y.; Zhong, Y.; Chen, C.; Zhao, Y.; Zeng, D.; Wu, Y.; Ding, P. A Dataset on the Life-history and Ecological Traits of Chinese Birds. Biodivers. Sci. 2021, 29, 1149–1153. [Google Scholar] [CrossRef]
- Wilman, H.; Belmaker, J.; Simpson, J.; de la Rosa, C.; Rivadeneira, M.M.; Jetz, W. EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar] [CrossRef]
- Botta-Dukát, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Swenson, N.G. Functional and Phylogenetic Ecology in R; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Rubolini, D.; Liker, A.; Garamszegi, L.Z.; Møller, A.P.; Saino, N. Using the BirdTree. org website to obtain robust phylogenies for avian comparative studies: A primer. Curr. Zool. 2015, 61, 959–965. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLOS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Mammola, S.; Carmona, C.P.; Guillerme, T.; Cardoso, P. Concepts and applications in functional diversity. Funct. Ecol. 2021, 35, 1869–1885. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Helmus, M.R.; Bland, T.J.; Williams, C.K.; Ives, A.R. Phylogenetic Measures of Biodiversity. Am. Nat. 2007, 169, E68–E83. [Google Scholar] [CrossRef]
- Ali, A.; Yan, E.-R. Consequences of phylogenetic conservativeness and functional trait similarity on aboveground biomass vary across subtropical forest strata. For. Ecol. Manag. 2018, 429, 28–35. [Google Scholar] [CrossRef]
- Webb, C.O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.P.; Bravo, G.A.; Brumfield, R.T.; Tello, J.G.; Cadena, C.D. A phylogenetic approach to disentangling the role of competition and habitat filtering in community assembly of Neotropical forest birds. J. Anim. Ecol. 2010, 79, 1181–1192. [Google Scholar] [PubMed]
- Laliberté, E.; Legendre, P.; Shipley, B. Package ‘FD’. Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. 2014. Available online: https://www.scirp.org/reference/ReferencesPapers?ReferenceID=1820451 (accessed on 2 March 2025).
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Team, R.C. R Version 4.3.3; R Core Team: Vienna, Austria, 2024. [Google Scholar]
- Dong, K. β-diversity Patterns of Bird Communities in Natural Protected Areas in Anhui by Separating the Turnover and Nestedness Components. Pak. J. Zool. 2023. [Google Scholar]
- Martínez-Abraín, A.; Jiménez, J. Anthropogenic areas as incidental substitutes for original habitat. Conserv. Biol. 2016, 30, 593–598. [Google Scholar]
- Salaverri, L.; Guitián, J.; Munilla, I.; Sobral, M. Bird richness decreases with the abandonment of agriculture in a rural region of SW Europe. Reg. Environ. Chang. 2019, 19, 245–250. [Google Scholar] [CrossRef]
- Padilla, B.J.; Sutherland, C. Drivers of avian diversity and abundance across gradients of human influence. Landsc. Ecol. 2022, 37, 969–981. [Google Scholar]
- Sullivan, M.J.; Davies, R.G.; Mossman, H.L.; Franco, A.M. An anthropogenic habitat facilitates the establishment of non-native birds by providing underexploited resources. PLoS ONE 2015, 10, e0135833. [Google Scholar]
- Sol, D.; Griffin, A.S.; Bartomeus, I.; Boyce, H. Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS ONE 2011, 6, e19535. [Google Scholar] [CrossRef]
- Alroy, J. Effects of habitat disturbance on tropical forest biodiversity. Proc. Natl. Acad. Sci. USA 2017, 114, 6056–6061. [Google Scholar]
- Pérez, G.; Mills, S.C.; Socolar, J.B.; Martínez-Revelo, D.E.; Haugaasen, T.; Gilroy, J.J.; Edwards, D.P. Avian phylogenetic and functional diversity are better conserved by land-sparing than land-sharing farming in lowland tropical forests. J. Appl. Ecol. 2024, 61, 2497–2509. [Google Scholar] [CrossRef]
- Htay, T.; Røskaft, E.; Ringsby, T.H.; Ranke, P.S. Spatio-temporal variation in avian taxonomic, functional, and phylogenetic diversity and its relevance for conservation in a wetland ecosystem in Myanmar. Biodivers. Conserv. 2023, 32, 2841–2867. [Google Scholar] [CrossRef]
- Liang, C.; Yang, G.; Wang, N.; Feng, G.; Yang, F.; Svenning, J.-C.; Yang, J. Taxonomic, phylogenetic and functional homogenization of bird communities due to land use change. Biol. Conserv. 2019, 236, 37–43. [Google Scholar] [CrossRef]
- Somveille, M.; Manica, A.; Butchart, S.H.M.; Rodrigues, A.S.L. Mapping Global Diversity Patterns for Migratory Birds. PLoS ONE 2013, 8, e70907. [Google Scholar]
- Xie, S.; Ouyang, Z.; Gong, C.; Meng, N.; Lu, F. Seasonal fluctuations of urban birds and their responses to immigration: An example from Macau, China. Urban For. Urban Green. 2021, 59, 126936. [Google Scholar]
- Sreekar, R.; Si, X.; Sam, K.; Liu, J.; Dayananda, S.; Goodale, U.; Kotagama, S.; Goodale, E. Land use and elevation interact to shape bird functional and phylogenetic diversity and structure: Implications for designing optimal agriculture landscapes. J. Appl. Ecol. 2021, 58, 1738–1748. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Ackerly, D.D. Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest. Ecol. Monogr. 2010, 80, 401–422. [Google Scholar]
- Zhao, Y.; Mendenhall, C.D.; Matthews, T.J.; Wang, D.; Li, W.; Liu, X.; Tang, S.; Han, P.; Wei, G.; Kang, Y. Land-use change interacts with island biogeography to alter bird community assembly. Proc. R. Soc. B 2024, 291, 20232245. [Google Scholar]
- Tilman, D. Niche tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 10854–10861. [Google Scholar]
- Diamond, J.M. The island dilemma: Lessons of modern biogeographic studies for the design of natural reserves. Biol. Conserv. 1975, 7, 129–146. [Google Scholar] [CrossRef]
- Si, X.; Cadotte, M.W.; Zeng, D.; Baselga, A.; Zhao, Y.; Li, J.; Wu, Y.; Wang, S.; Ding, P. Functional and phylogenetic structure of island bird communities. J. Anim. Ecol. 2017, 86, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Cadotte, M.W.; Carboni, M.; Si, X.; Tatsumi, S. Do traits and phylogeny support congruent community diversity patterns and assembly inferences? J. Ecol. 2019, 107, 2065–2077. [Google Scholar] [CrossRef]
- Che, X.; Chen, D.; Zhang, M.; Quan, Q.; Møller, A.P.; Zou, F. Seasonal dynamics of waterbird assembly mechanisms revealed by patterns in phylogenetic and functional diversity in a subtropical wetland. Biotropica 2019, 51, 421–431. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Traits | Trait Types | Value Type | K | p |
|---|---|---|---|---|
| Body mass | Body mass in grams | Continuous | 0.469 | <0.001 |
| Diet | Invertebrates | Binary | 0.363 | 0.007 |
| Vertebrates | Binary | 0.041 | 0.547 | |
| Scavenger | Binary | 0.259 | 0.042 | |
| Nectar and fruit | Binary | 0.285 | <0.001 | |
| Seed | Binary | 0.272 | <0.001 | |
| Plant material | Binary | 0.189 | 0.128 | |
| Foraging stratum | Water | Binary | 2.405 | <0.001 |
| Ground | Binary | 0.243 | 0.003 | |
| Understory | Binary | 0.169 | 0.578 | |
| Mid to high | Binary | 0.236 | 0.279 | |
| Canopy | Binary | 0.461 | 0.071 | |
| Aerial | Binary | 0.257 | 0.041 |
| Land Use Types | Abundance | Species Richness | Unique Species | Threated Species |
|---|---|---|---|---|
| Villages | 6468 | 111 | 5 (8) | 14 (94) |
| Farmlands | 5544 | 147 | 30 (161) | 24 (196) |
| Forests | 3703 | 104 | 11 (30) | 16 (297) |
| All habitats | 15,715 | 169 | / | 31 (587) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, S.; He, Y.; Wang, X.; Wang, G.; Zhou, Y.; Li, C. Response of Bird Communities to Human-Dominated Habitats in Southern Anhui Mountainous Area, China. Diversity 2025, 17, 261. https://doi.org/10.3390/d17040261
Dai S, He Y, Wang X, Wang G, Zhou Y, Li C. Response of Bird Communities to Human-Dominated Habitats in Southern Anhui Mountainous Area, China. Diversity. 2025; 17(4):261. https://doi.org/10.3390/d17040261
Chicago/Turabian StyleDai, Shuheng, Yurong He, Xue Wang, Guangyao Wang, Yixue Zhou, and Chunlin Li. 2025. "Response of Bird Communities to Human-Dominated Habitats in Southern Anhui Mountainous Area, China" Diversity 17, no. 4: 261. https://doi.org/10.3390/d17040261
APA StyleDai, S., He, Y., Wang, X., Wang, G., Zhou, Y., & Li, C. (2025). Response of Bird Communities to Human-Dominated Habitats in Southern Anhui Mountainous Area, China. Diversity, 17(4), 261. https://doi.org/10.3390/d17040261