
Introgressive Hybridization and a Natural Barrier: Molecular Analyses Lay Out a Conservation Blueprint for the Critically Endangered Vietnamese Pond Turtle (Mauremys annamensis)

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
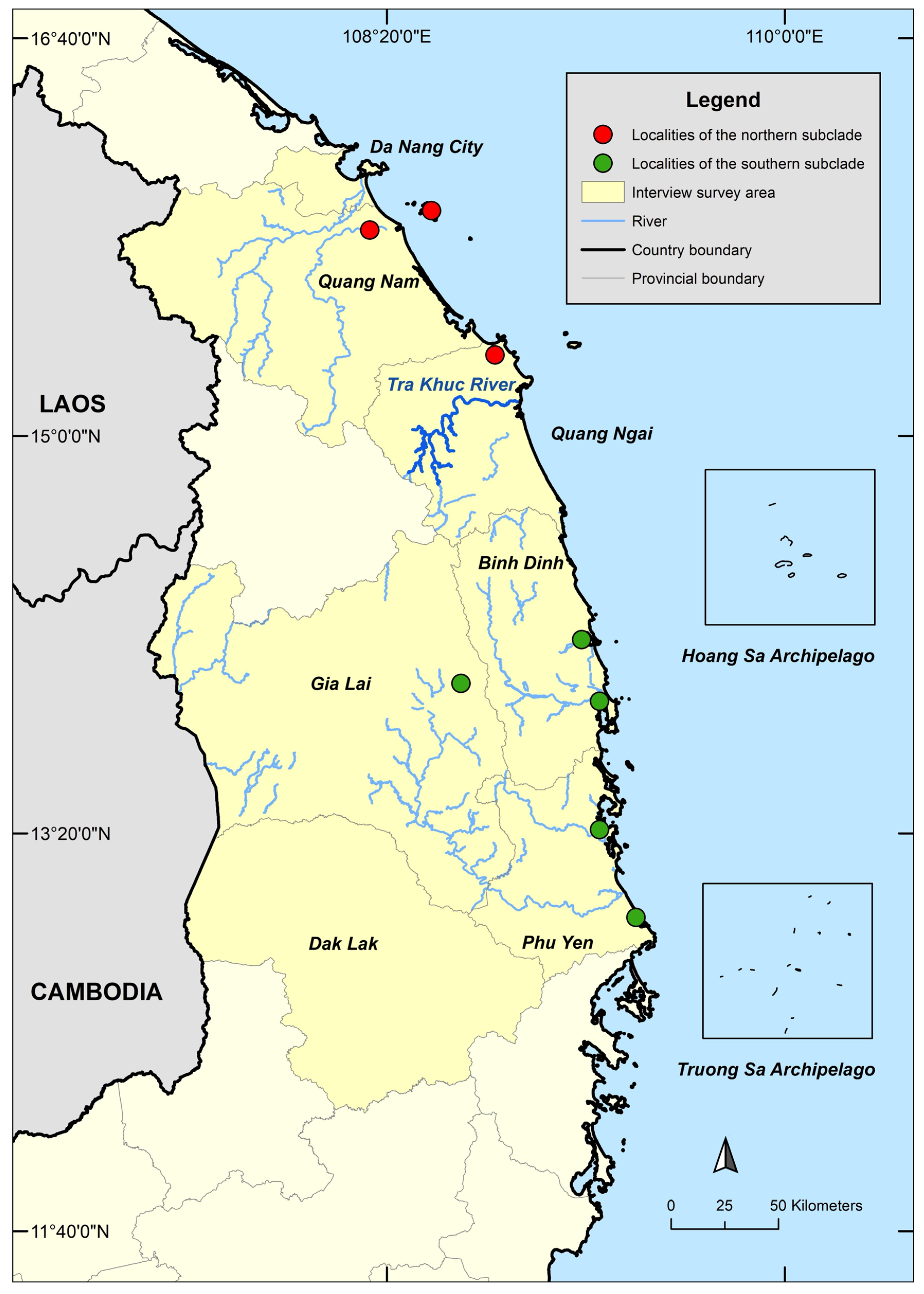
2.1. Interviews
2.2. Taxonomic Sampling
2.3. Molecular Data
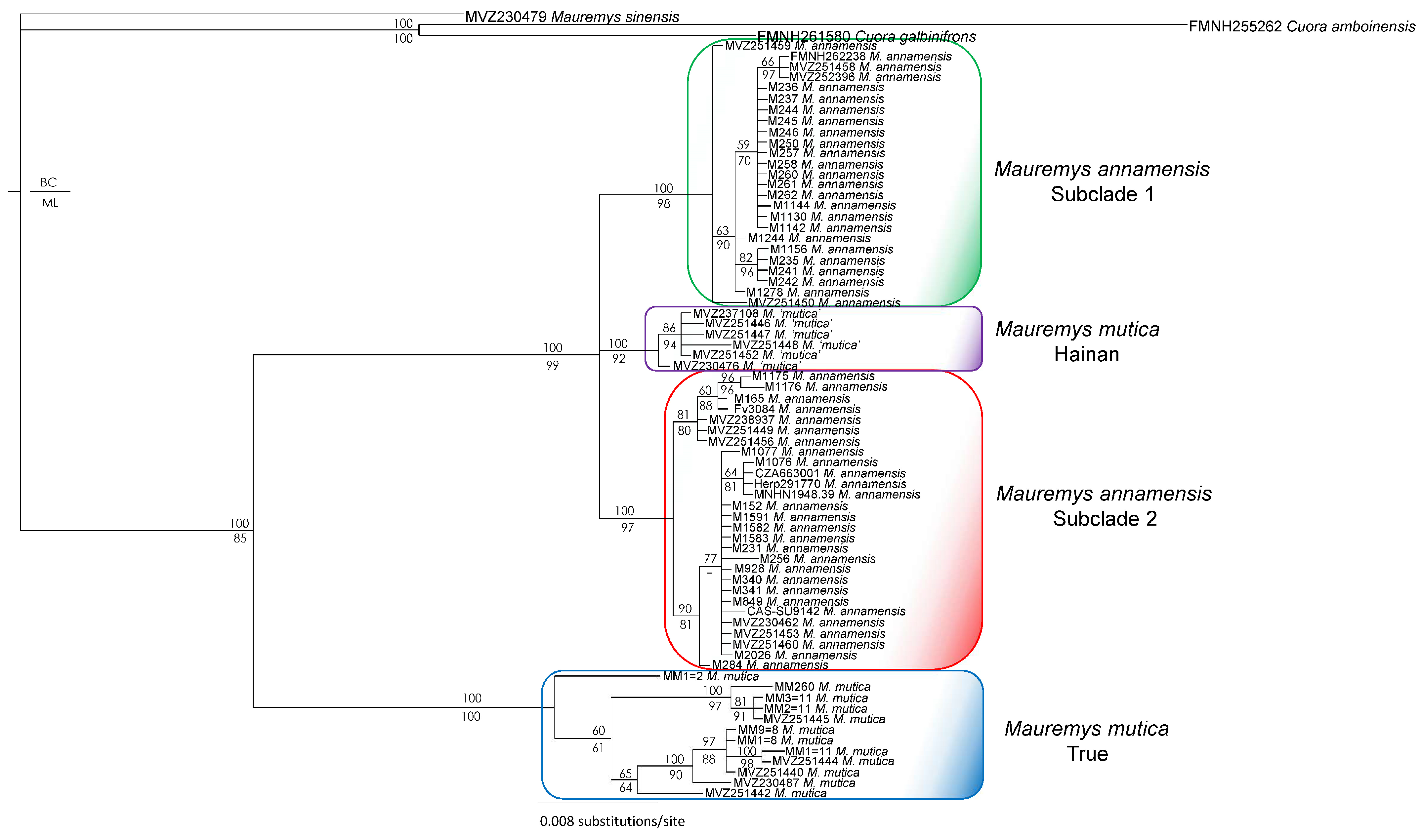
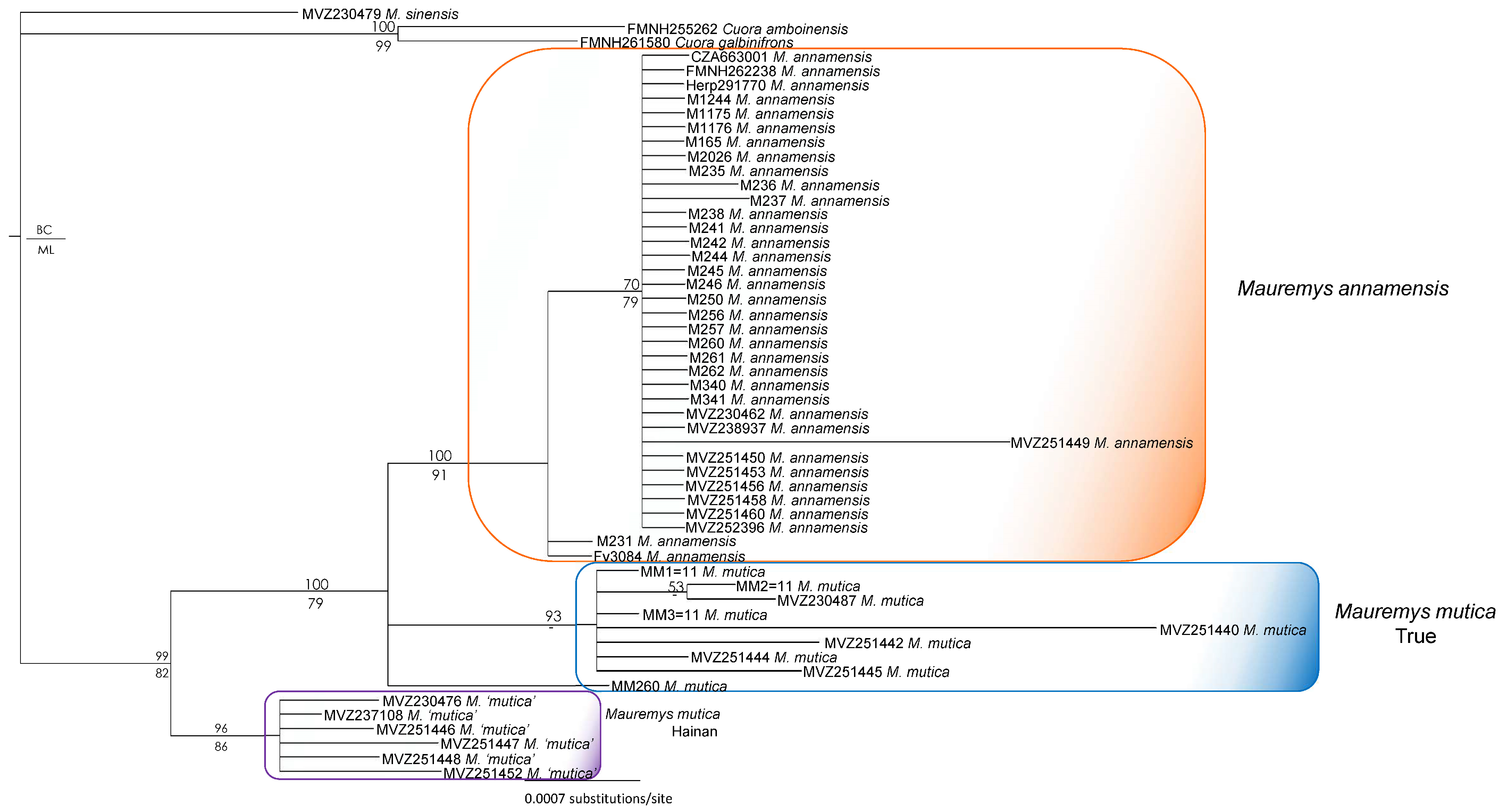
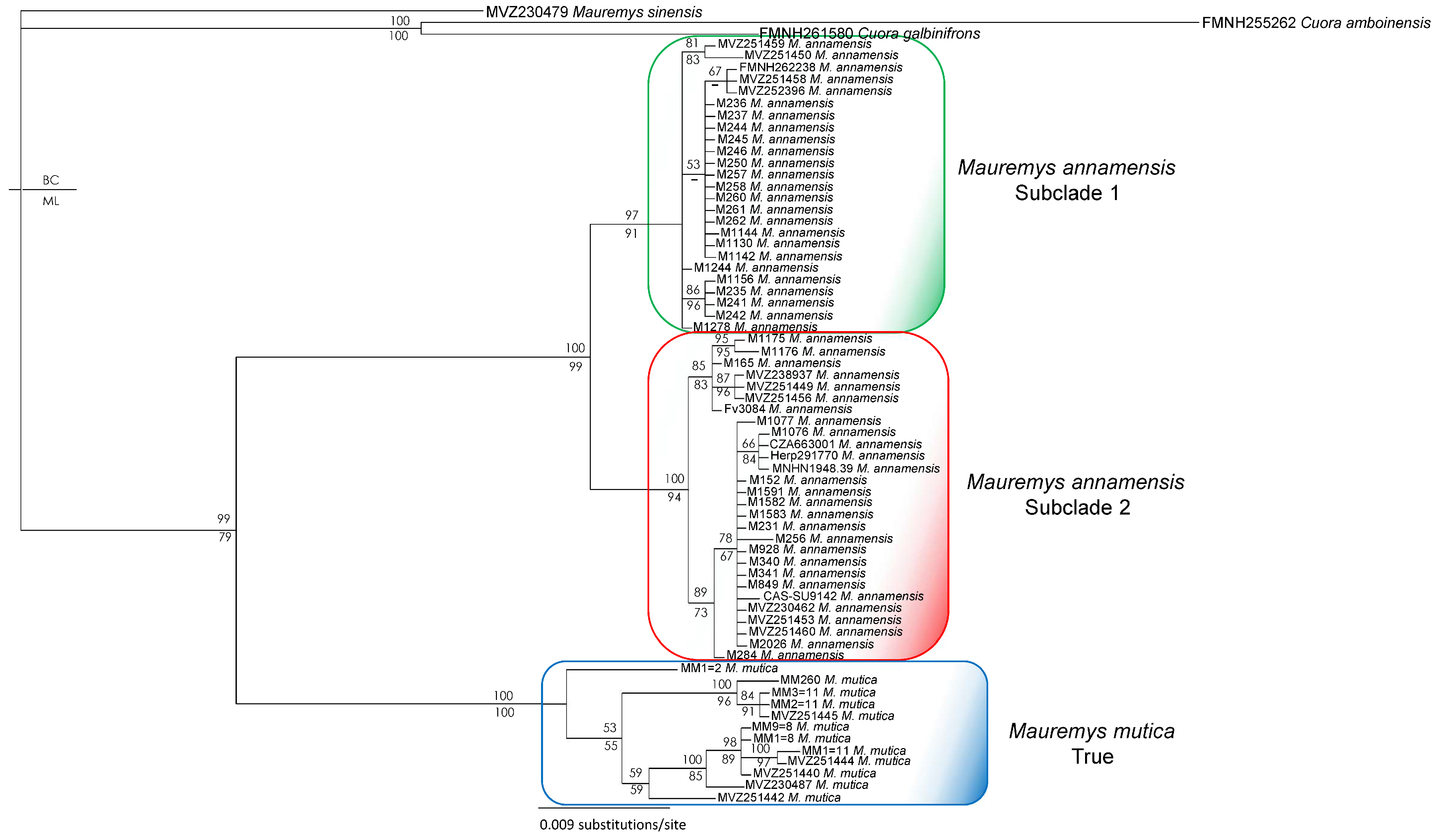
2.4. Phylogenetic and Network Analyses
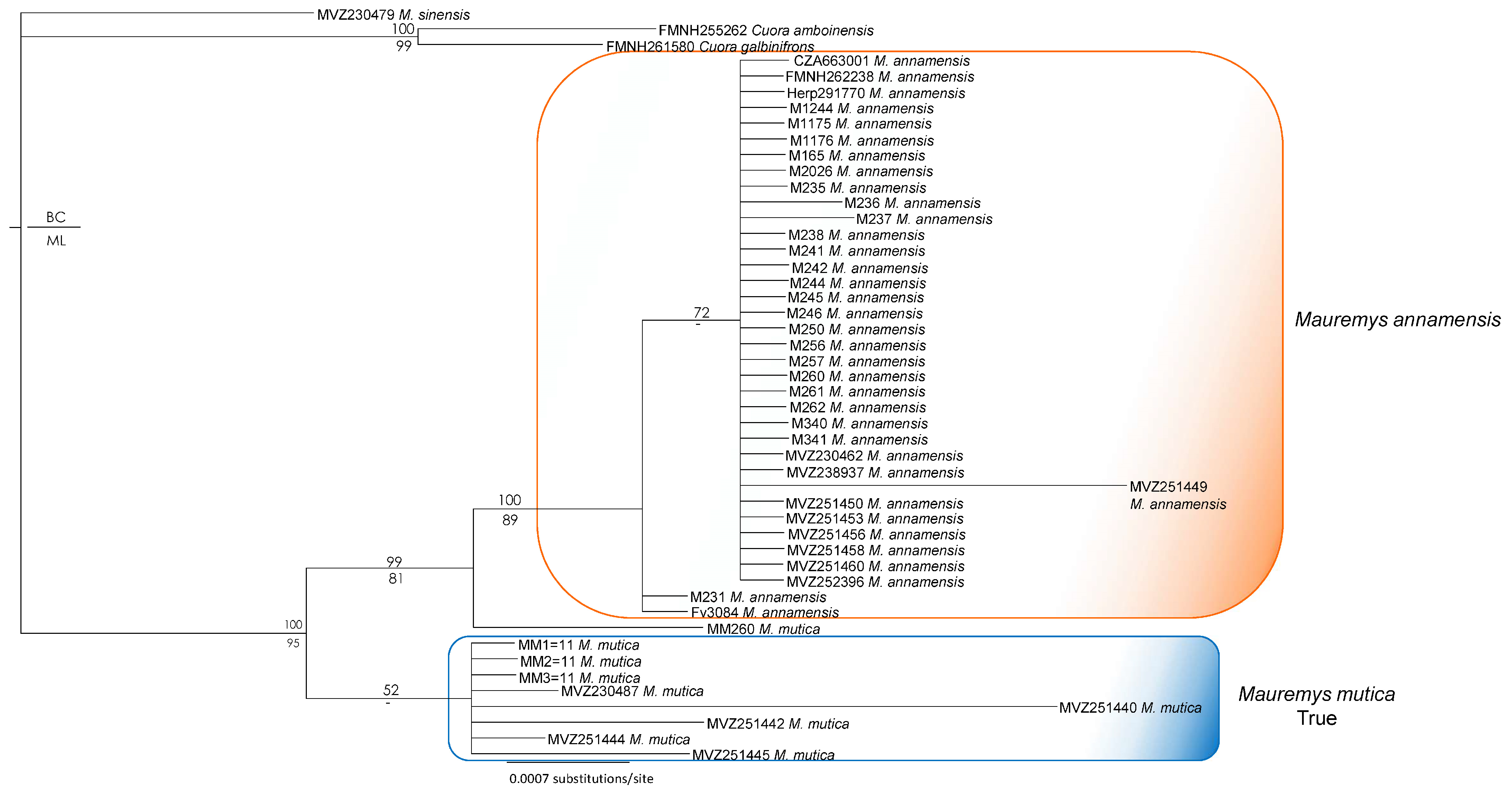
3. Results
4. Discussion
5. Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bourret, R. Les tortues de l’Indochine; Institut Océanographique Indochine: Nha Trang, Vietnam, 1941; pp. 1–235. [Google Scholar]
- Iverson, J.B. A Revised Checklist with Distribution Maps of the Turtles of the World; Privately Printed: Richmond, IN, USA, 1992; pp. 1–359. [Google Scholar]
- Le, M.; Hoang, T.; Le, D. Trade data and some comments on the distribution of Mauremys annamensis (Siebenrock, 1903). Asiat. Herpetol. Res. 2004, 10, 110–113. [Google Scholar]
- Dawson, J.E.; Hatcher, R.E.; Calloway-Burke, G.E. Mauremys annamensis: Locality and longevity. Herpetol. Rev. 2013, 44, 91–93. [Google Scholar]
- McCormack, T.E.M.; Dawson, J.E.; Hendrie, D.B.; Ewert, M.A.; Iverson, J.B.; Hatcher, R.E.; Goode, J.M. Mauremys annamensis (Siebenrock 1903)—Vietnamese Pond Turtle, Annam Pond Turtle, Rùa Trung Bộ. In Conservation Biology of Freshwater Turtles and Tortoises; Rhodin, A.G.J., Pritchard, P.C.H., van Dijk, P.P., Saumure, R.A., Buhlmann, K.A., Iverson, J.B., Mittermeier, R.A., Eds.; A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle Specialist Group; Chelonian Research Monographs; IUCN Species Survival Commission: Gland, Switzerland, 2014; Volume 57, pp. 1–14. [Google Scholar] [CrossRef]
- McCormack, T.; Hendrie, D.; Nguyen, X.T. Rediscovery after 67 years of the critically endangered Vietnamese Pond Turtle, Mauremys annamensis, in central Vietnam. In Program and Abstracts of the Fifth Annual Symposium on the Conservation and Biology of Tortoises and Freshwater Turtles; Schaffer, C., Ed.; Turtle Survival Alliance: Atlanta, GA, USA, 2007. [Google Scholar]
- Parham, J.F.; Stuart, B.L.; Orlov, N.L. Geographic distribution: Mauremys annamensis. Herpetol. Rev. 2006, 37, 239. [Google Scholar]
- CRES (Centre for Natural Resources and Environmental Studies). CEPF Final Project Completion Report; CRES: Hanoi, Vietnam, 2016; pp. 1–8. [Google Scholar]
- Stanford, C.B.; Rhodin, A.G.J.; van Dijk, P.P.; Horne, B.D.; Blanck, T.; Goode, E.V.; Hudson, R.; Mittermeier, R.A.; Currylow, A.; Eisemberg, C.; et al. Turtles in Trouble: The World’s 25+ Most Endangered Tortoises and Freshwater Turtles—2018; Hemlock Printers: Ojai, CA, USA, 2018. [Google Scholar]
- McCormack, T.; van Dijk, P.P.; Roberton, S.; Dawson, J.E. Mauremys annamensis (Amended Version of 2020 Assessment). In The IUCN Red List of Threatened Species. 2020. e.T12876A182354172. Available online: https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T12876A182354172.en (accessed on 30 March 2025).
- Tran, K.; Ho, C.T.; Nguyen, S.V.; Pham, T. [Reptiles and amphibians]. Part 1. Animals. In Vietnam Red Data Book; Dang, N.T., Tran, K., Dang, H.H., Nguyen, C., Nguyen, T.N., Nguyen, Y.N., Dang, D.T., Eds.; Science and Technics Publishing House: Hanoi, Vietnam, 2007; pp. 219–276. (In Vietnamese) [Google Scholar]
- Raffel, M.; Meier, E. Best Year Ever at the International Centre for the Conservation of Turtles (IZS) at Muenster Zoo; Turtle Survival—A Publication of the Turtle Survival Alliance: Forth Worth, TX, USA, 2013. [Google Scholar]
- Shaffer, C. Vietnamese Pond Turtle program. Turt. Tortoise Newsl. 2006, 9, 13–14. [Google Scholar]
- Somerová, B.; Rehák, I.; Velenský, P.; Palupčíková, K.; Protiva, T.; Frynta, D. Haplotype variation in founders of the Mauremys annamensis population kept in European zoos. Acta Herpetol. 2015, 10, 7–15. [Google Scholar] [CrossRef]
- Parham, J.F.; Simison, W.B.; Kozak, K.H.; Feldman, C.R.; Shi, H. New Chinese turtles: Endangered or invalid? A reassessment of two species using mitochondrial DNA, allozyme electrophoresis and known-locality specimens. Anim. Conserv. 2001, 4, 357–367. [Google Scholar] [CrossRef]
- Shi, H.; Parham, J.F. Preliminary observations of a large turtle farm in Hainan Province, People’s Republic of China. Turt. Tortoise Newsl. 2001, 3, 4–6. [Google Scholar]
- Zhou, T.; Blanck, T.; McCord, W.P.; Li, P.P. Tracking Cuora mccordi Ernst, 1988: The first records of its natural habitat; a re-description; with data on captive populations and its vulnerability. Hamadryad 2008, 32, 57–69. [Google Scholar]
- Fong, J.J.; Chen, T.H. DNA evidence for the hybridization of wild turtles in Taiwan: Possible genetic pollution from trade animals. Conserv. Genet. 2010, 11, 2061–2066. [Google Scholar] [CrossRef]
- Feldman, C.R.; Parham, J.F. Molecular systematics of Old World Stripe-Necked Turtle (Testudines: Mauremys). Asiat. Herpetol. Res. 2004, 10, 28–37. [Google Scholar]
- Fong, J.J.; Parham, J.F.; Shi, H.; Stuart, B.L.; Carter, R.L. A genetic survey of heavily exploited, endangered turtles: Caveats on the conservation value of trade animals. Anim. Conserv. 2007, 10, 452–460. [Google Scholar] [CrossRef]
- Fong, J.J.; Stuart, B.L.; McCormack, T.E.M.; Parham, J.F. First genetic data of the critically endangered Vietnamese Pond Turtle (Mauremys annamensis) from known-locality specimens. Curr. Herpetol. 2019, 38, 140–152. [Google Scholar] [CrossRef]
- Zhao, J.; Li, W.; Wen, P.; Zhang, D.; Zhu, X. Genetic diversity and relationship of Mauremys mutica and M. annamensis assessed by DNA barcoding sequences. Mitochondrial DNA Part A 2015, 27, 3507–3510. [Google Scholar] [CrossRef]
- Le, M.D.; McCormack, T.E.M.; Hoang, H.V.; Duong, H.T.; Nguyen, T.Q.; Ziegler, T.; Nguyen, H.D.; Ngo, H.T. Threats from wildlife trade: The importance of genetic data in safeguarding the endangered Four-eyed Turtle (Sacalia quadriocellata). Nat. Conserv. 2020, 41, 91–111. [Google Scholar] [CrossRef]
- Creswell, J.W.; Poth, C.N. Qualitative Inquiry Research Design: Choosing Among Five Approaches; Sage Publications: Thousand Oaks, CA, USA, 2017. [Google Scholar]
- Spinks, P.Q.; Shaffer, H.B.; Iverson, J.B.; McCord, W.P. Phylogenetic hypotheses for the turtle family Geoemydidae. Mol. Phylogenetics Evol. 2004, 32, 164–182. [Google Scholar] [CrossRef]
- Engstrom, T.N.; Shaffer, H.B.; McCord, W.P. Phylogenetic diversity of endangered and critically endangered southeast Asian softshell turtles (Trionychidae: Chitra). Biol. Conserv. 2002, 104, 173–179. [Google Scholar] [CrossRef]
- Arévalo, E.; Davis, S.K.; Sites, J.W. Mitochondrial DNA sequence divergence and phylogenetic relationships among eight chromosome races of the Sceloporus grammicus complex (Phrynosomatidae) in central Mexico. Syst. Biol. 1994, 43, 387–418. [Google Scholar] [CrossRef]
- Fujita, M.K.; Engstrom, T.N.; Starkey, D.E.; Shaffer, H.B. Turtle phylogeny: Insights from a novel nuclear intron. Mol. Phylogenetics Evol. 2004, 31, 1031–1040. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A. MrBayes 3.2: Efficient Bayesian phylogenetic. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Bui, M.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Huson, H.D.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Nylander, J.A.A.; Ronquist, F.; Huelsenback, J.P.; Nieves-Aldrey, J.L. Bayesian phylogenetic analysis of combined data. Syst. Biol. 2004, 53, 47–67. [Google Scholar] [CrossRef]
- Brandley, M.C.; Schmitz, A.; Reeder, T.W. Partitioned Bayesian analyses, partition choice, and the phylogenetic relationships of scincid lizards. Syst. Biol. 2005, 54, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (* and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.; Moulton, V. Neighbor-Net: An Agglomerative Method for the Construction of Planar Phylogenetic Networks. Mol. Biol. Evol. 2004, 21, 255–265. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Nguyen, L.T.; Tran, T.-D.T.; Phan, S.C.; Ziegler, T.; Le, M.; McCormack, T.E.M. First offshore island record of Mauremys annamensis (Siebenrock, 1903) (Testudines, Geoemydidae) in Vietnam. Check List 2025, 21, 1–5. [Google Scholar]
- Boettger, O. Materialien zur herpetologi-schen fauna von China III. I. liste der von der insel Hainan bekannten kriechtiere. In Bericht der Senckenbergischen Naturforschenden Gesell-Schaft in Frankfurt am Main; Die Gesellschaft: Frankfurt, Germany, 1894; Volume 25, pp. 129–136. [Google Scholar]
- Mallet, J. Hybrid speciation. Nature 2007, 446, 279–283. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Schumer, M.; Kruesi, K.; Walter, R.; Andolfatto, P.; Rosenthal, G.G. Phylogenomics reveals extensive reticulate evolution in Xiphophorus fishes. Evolution 2013, 67, 2166–2179. [Google Scholar] [CrossRef] [PubMed]
- Spinks, P.Q.; Shaffer, H.B. Conflicting mitochondrial and nuclear phylogenies for the widely disjunct Emys (Testudines: Emydidae) species complex, and what they tell us about biogeography and hybridization. Syst. Biol. 2009, 61, 1–20. [Google Scholar] [CrossRef]
- Thomson, R.C.; Spinks, P.Q.; Shaffer, H.B. Molecular phylogeny and divergence of the map turtles (Emydidae: Graptemys). Mol. Phylogenetics Evol. 2018, 121, 61–70. [Google Scholar] [CrossRef]
- Kehlmaier, C.; Zhang, X.; Georges, A.; Campbell, P.D.; Thomson, S.; Fritz, U. Mitogenomics of historical type specimens of Australasian turtles: Clarification of taxonomic confusion and old mitochondrial introgression. Sci. Rep. 2019, 9, 5841. [Google Scholar] [CrossRef]
- Herbert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Protiva, T. Genetic and shell shape variability in Mauremys annamensis. Turt. Surviv. Mag. 2015, 15, 56. [Google Scholar]
- Zwartepoorte, H.; Becker, H.; Meier, E.; Raffel, M.; McCormack, T. Captive breeding critical to saving the highly endangered Vietnamese Pond Turtle (Mauremys annamensis). Turt. Surviv. Mag. 2015, 15, 54–55. [Google Scholar]
- Ziegler, T.; Nguyen, T.Q.; Le, M.D.; Dieckmann, R.; Haase, C.; Haizak, C.; Heckel, J.-O.; Lefaux, B.; Mager, C.; Michel, V.; et al. The ‘VIETNAMAZING’ EAZA conservation campaign 2024–2025. WAZA News 2024, 1, 42–47. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Names | GenBank No. (ND4) | GenBank No. (R35) | Voucher Number/ Lab Number | Origin | Subclade Assignment |
|---|---|---|---|---|---|
| C. amboinensis | AY364609 | DQ386654 | FMNH255262 | Laos: Champasak Province | - |
| C. galbinifrons | AY364611 | DQ386660 | FMNH261580 | Pet trade | - |
| M. sinensis | AY337345 | DQ386678 | MVZ230479 | China: Hainan Province | - |
| M. mutica | EF034089 | EF587932 | MVZ251440 | China: Turtle farm; Hainan Province | M. mutica true |
| M. mutica | EF034091 | EF587916 | MVZ251442 | China: Turtle farm; Hainan Province | M. mutica true |
| M. mutica | EF034093 | EF587931 | MVZ251444 | China: Turtle farm; Hainan Province | M. mutica true |
| M. mutica | EF034094 | EF587927 | MVZ251445 | China: Turtle farm; Hainan Province | M. mutica true |
| M. mutica | GQ259450 | - | MM1 = 2 | Taiwan: Changhua Country | M. mutica true |
| M. mutica | GQ259455 | - | MM9 = 8 | Taiwan: Keelung City | M. mutica true |
| M. mutica | GQ259456 | - | MM1 = 8 | Taiwan: Hsinchu Country | M. mutica true |
| M. mutica | GQ259457 | GQ259474 | MM1 = 11 | Taiwan: Yilan Country | M. mutica true |
| M. mutica | GQ259452 | GQ259471 | MM260 | Taiwan: Hualien Country | M. mutica true |
| M. mutica | GQ259453 | GQ259472 | MM3 = 11 | Taiwan: Yilan Country | M. mutica true |
| M. mutica | GQ259454 | GQ259473 | MM2 = 11 | Taiwan: Taoyuan City | M. mutica true |
| M. mutica | AF348278 | DQ386666 | MVZ230487 | China: Zhoushan Island, Zhejiang Province | M. mutica true |
| M. ‘mutica’ | EF034095 | EF587930 | MVZ251446 | China: Turtle farm; Hainan Province | M. mutica Hainan |
| M. ‘mutica’ | EF034096 | EF587929 | MVZ251447 | China: Turtle farm; Hainan Province | M. mutica Hainan |
| M. ‘mutica’ | EF034097 | EF587925 | MVZ251448 | China: Turtle farm; Hainan Province | M. mutica Hainan |
| M. ‘mutica’ | EF034101 | EF587917 | MVZ251452 | China: Qiongzhong Country, Hainan Province | M. mutica Hainan |
| M.‘mutica’ | EF034114 | EF587915 | MVZ237108 | China: Qiongzhong Country, Hainan Province | M. mutica Hainan |
| M.耙mutica’ | AF348282 | DQ386664 | MVZ230476 | China: Turtle farm; Hainan Province | M. mutica Hainan |
| M. annamensis | AY337338 | EF587933 | MVZ238937 | China: Turtle farm; Hainan Province | M. annamensis subclade 2 |
| M. annamensis | EF034098 | EF587934 | MVZ251449 | China: Turtle farm; Hainan Province | M. annamensis subclade 2 |
| M. annamensis | EF034099 | EF587919 | MVZ251450 | China: Turtle farm; Hainan Province | M. annamensis subclade 1 |
| M. annamensis | EF034102 | EF587920 | MVZ251453 | Vietnam Trade: Ninh Binh Province | M. annamensis subclade 2 |
| M. annamensis | EF034105 | EF587922 | MVZ251456 | Vietnam Trade: Ninh Binh Province | M. annamensis subclade 2 |
| M. annamensis | EF034107 | EF587924 | MVZ251458 | Vietnam Trade: Thanh Hoa Province | M. annamensis subclade 1 |
| M. annamensis | EF034109 | EF587926 | MVZ251460 | Vietnam Trade: Binh Dinh Province | M. annamensis subclade 2 |
| M. annamensis | EF034108 | - | MVZ251459 | Vietnam Trade: Binh Dinh Province | M. annamensis subclade 1 |
| M. annamensis | EF034113 | DQ386655 | MVZ230462 | China Market: Hainan Province | M. annamensis subclade 2 |
| M. annamensis | EF587914 | EF587921 | MVZ252396 | Vietnam Trade: Thanh Hoa Province | M. annamensis subclade 1 |
| M. annamensis | EF034112 | DQ386656 | FMNH262238 | Pet trade | M. annamensis subclade 1 |
| M. annamensis | MK726299 | MK726303 | CZA663001 | Vietnam: Da Nang Municipality | M. annamensis subclade 2 |
| M. annamensis | MK726300 | MK726304 | Herp291770 | Vietnam: Quang Nam Province, Dien Ban District | M. annamensis subclade 2 |
| M. annamensis | MK726301 | - | MNHN 1948.39 | Vietnam: Quang Nam Province, Fai Fo (Hoi An) | M. annamensis subclade 2 |
| M. annamensis | MK726302 | - | CAS-SU9142 | Vietnam: Quang Nam Province, Fai Fo (Hoi An) | M. annamensis subclade 2 |
| M. annamensis | PV388604 | PV388581 | M1244 | Vietnam: Gia Lai Province | M. annamensis subclade 1 |
| M. annamensis | PV388605 | - | M1278 | Vietnam: Gia Lai Province | M. annamensis subclade 1 |
| M. annamensis | PV388606 | - | M284 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388607 | - | M849 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388608 | PV388582 | M1175 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388609 | PV388583 | M1176 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388610 | - | M1077 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388611 | - | M1076 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388612 | - | M1156 | Vietnam: Binh Dinh Province | M. annamensis subclade 1 |
| M. annamensis | PV388613 | - | M1591 | Vietnam: Quang Ngai Province | M. annamensis subclade 2 |
| M. annamensis | PV388614 | PV388584 | M2026 | Vietnam: Quang Ngai Province | M. annamensis subclade 2 |
| M. annamensis | PV388615 | - | M1582 | Vietnam: Quang Ngai Province | M. annamensis subclade 2 |
| M. annamensis | PV388616 | - | M1583 | Vietnam: Quang Ngai Province | M. annamensis subclade 2 |
| M. annamensis | PV388617 | - | M928 | Vietnam: Quang Ngai Province | M. annamensis subclade 2 |
| M. annamensis | PV388618 | - | M1144 | Vietnam: Phu Yen Province | M. annamensis subclade 1 |
| M. annamensis | PV388619 | - | M1130 | Vietnam: Phu Yen Province | M. annamensis subclade 1 |
| M. annamensis | PV388620 | - | M1142 | Vietnam: Phu Yen Province | M. annamensis subclade 1 |
| M. annamensis | PV388621 | PV388585 | Fv3084 | Vietnam: Quang Nam Province | M. annamensis subclade 2 |
| M. annamensis | PV388622 | - | M152 | Vietnam: TCC | M. annamensis subclade 2 |
| M. annamensis | PV388623 | PV388586 | M165 | Vietnam: TCC | M. annamensis subclade 2 |
| M. annamensis | PV388624 | PV388587 | M231 | Vietnam: TCC | M. annamensis subclade 2 |
| M. annamensis | PV388625 | PV388588 | M235 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388626 | PV388589 | M236 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388627 | PV388590 | M237 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388628 | PV388591 | M241 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388629 | PV388592 | M242 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388630 | PV388593 | M244 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388631 | PV388594 | M245 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388632 | PV388595 | M246 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388633 | PV388596 | M250 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388634 | PV388597 | M256 | Vietnam: TCC | M. annamensis subclade 2 |
| M. annamensis | PV388635 | PV388598 | M257 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388636 | - | M258 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388637 | PV388599 | M260 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388638 | PV388600 | M261 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388639 | PV388601 | M262 | Vietnam: TCC | M. annamensis subclade 1 |
| M. annamensis | PV388640 | PV388602 | M340 | Vietnam: TCC | M. annamensis subclade 2 |
| M. annamensis | PV388641 | PV388603 | M341 | Vietnam: TCC | M. annamensis subclade 2 |
| Primer | Sequence | Reference |
|---|---|---|
| ND672 (f) | 5′-TGACTACCAAAAGCTCATGTAGAAGC—3′ | [26] |
| Hist (r) | 5′-CCTATTTTTAGAGCCACAGTCTAATG—3′ | [27] |
| R35Ex1 (f) | 5′-ACGATTCTCGCTGATTCTTGC—3′ | [28] |
| R35Ex2 (r) | 5′-GCAGAAAACTGAATGTCTCAAAGG—3′ | [28] |
| Data Analysis | Models Selected | Burn-In Value |
|---|---|---|
| CONCATENATED MATRIX | ||
| Combined analysis | ||
| With Hainan’s M. mutica | HKY + I + G | 39,000 |
| Without Hainan’s M. mutica | TPM3UF + I + G | 40,000 |
| Partitioned Bayesian analysis | ||
| With Hainan’s M. mutica | ||
| ND4 1st position | TIM + I | 40,000 |
| ND4 2nd position | TRN | |
| ND4 3rd position | K81UF + G | |
| R35 | HKY + I + G | |
| Without Hainan’s M. mutica | ||
| ND4 1st position | GTR + I | 44,000 |
| ND4 2nd position | HKY + I | |
| ND4 3rd position | HKY + I | |
| R35 | HKY + I + G | |
| SEPARATE GENE MATRIX | ||
| ND4 with Hainan’s M. mutica | TIM1 + G | 38,000 |
| ND4 without Hainan’s M. mutica | TIM3 + I | 41,000 |
| R35 with Hainan’s M. mutica | HKY + G | 39,000 |
| R35 without Hainan’s M. mutica | TRN + G | 42,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngo, H.T.; McCormack, T.E.M.; Hoang, H.V.; Nguyen, L.T.; Nguyen, T.T.; Nguyen, V.T.H.; Do, H.T.; Trinh, H.L.T.; Ziegler, T.; Blair, M.E.; et al. Introgressive Hybridization and a Natural Barrier: Molecular Analyses Lay Out a Conservation Blueprint for the Critically Endangered Vietnamese Pond Turtle (Mauremys annamensis). Diversity 2025, 17, 260. https://doi.org/10.3390/d17040260
Ngo HT, McCormack TEM, Hoang HV, Nguyen LT, Nguyen TT, Nguyen VTH, Do HT, Trinh HLT, Ziegler T, Blair ME, et al. Introgressive Hybridization and a Natural Barrier: Molecular Analyses Lay Out a Conservation Blueprint for the Critically Endangered Vietnamese Pond Turtle (Mauremys annamensis). Diversity. 2025; 17(4):260. https://doi.org/10.3390/d17040260
Chicago/Turabian StyleNgo, Hanh T., Timothy E. M. McCormack, Ha V. Hoang, Luan T. Nguyen, Thuy T. Nguyen, Van T. H. Nguyen, Hao T. Do, Ha L. T. Trinh, Thomas Ziegler, Mary E. Blair, and et al. 2025. "Introgressive Hybridization and a Natural Barrier: Molecular Analyses Lay Out a Conservation Blueprint for the Critically Endangered Vietnamese Pond Turtle (Mauremys annamensis)" Diversity 17, no. 4: 260. https://doi.org/10.3390/d17040260
APA StyleNgo, H. T., McCormack, T. E. M., Hoang, H. V., Nguyen, L. T., Nguyen, T. T., Nguyen, V. T. H., Do, H. T., Trinh, H. L. T., Ziegler, T., Blair, M. E., & Le, M. D. (2025). Introgressive Hybridization and a Natural Barrier: Molecular Analyses Lay Out a Conservation Blueprint for the Critically Endangered Vietnamese Pond Turtle (Mauremys annamensis). Diversity, 17(4), 260. https://doi.org/10.3390/d17040260