
Characterization of the Unarmored Dinoflagellate Karlodinium decipiens (Dinophyceae) from Jiaozhou Bay, China


 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Culture Establishment
2.2. Light Microscopy (LM)
2.3. Scanning Electron Microscopy (SEM)
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Phylogenetic Analyses
2.6. Pigment Analyses
2.7. Bioassays for Possible Effects on Aquatic Organisms
3. Results
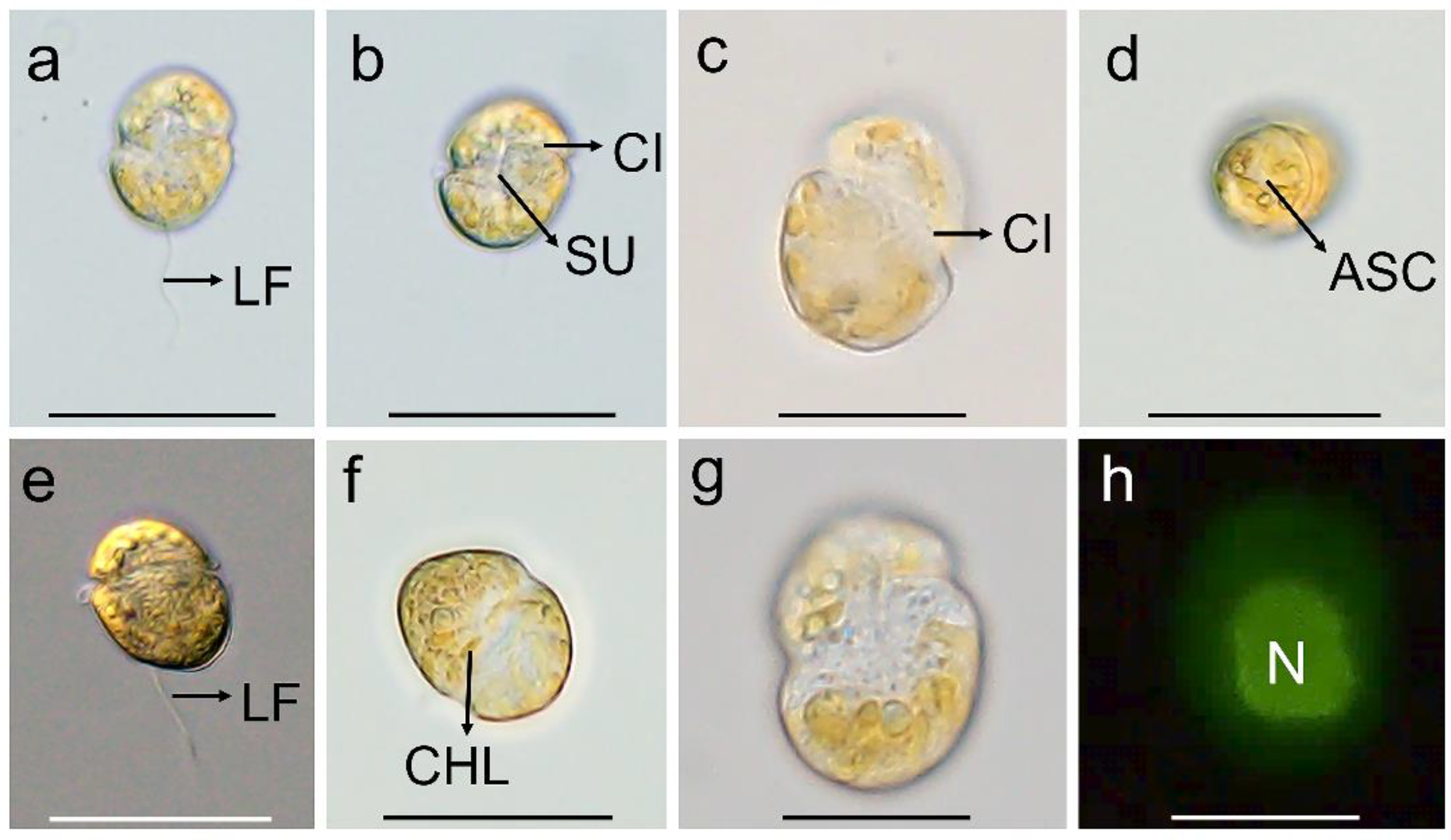
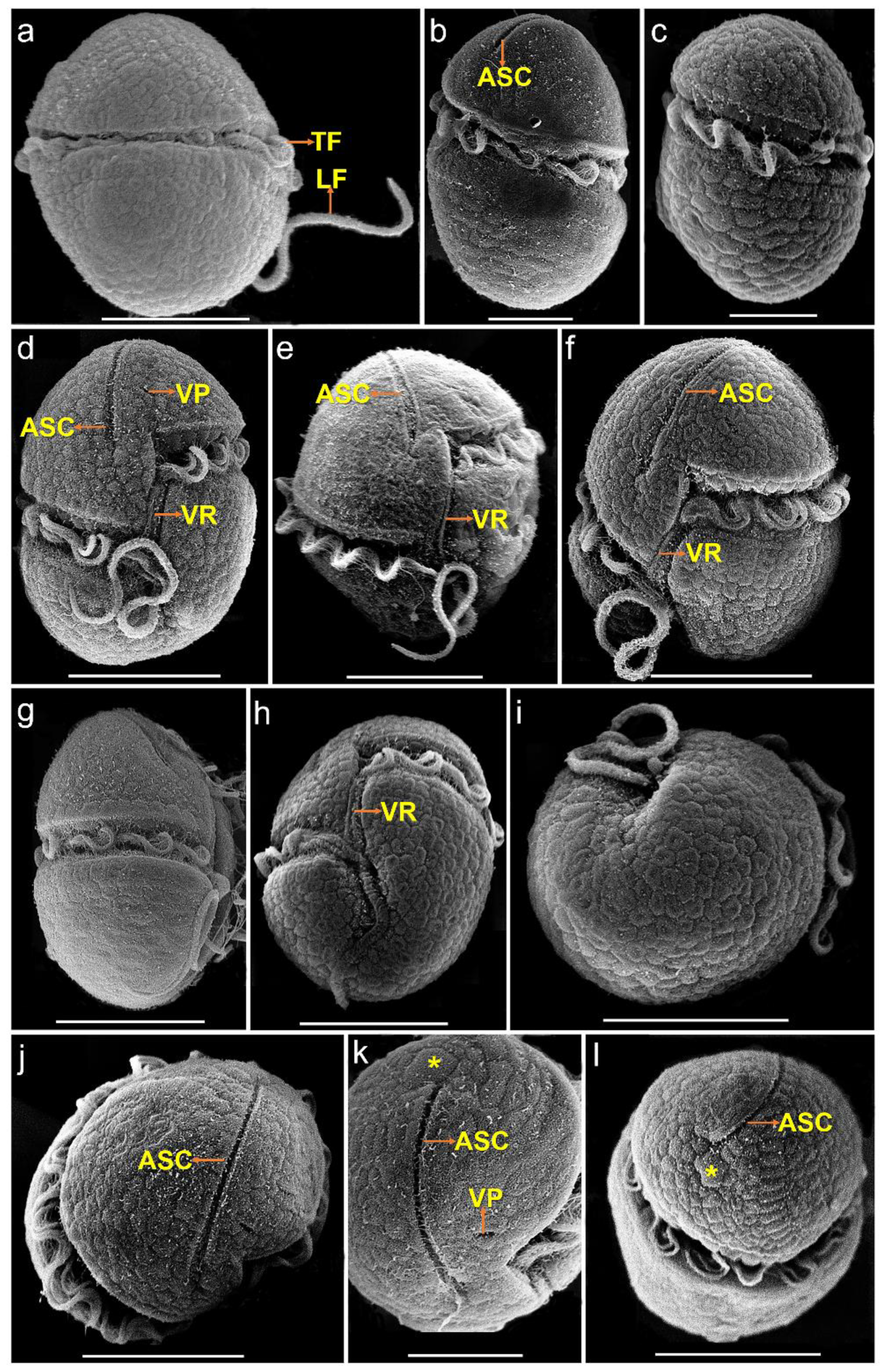
3.1. Morphology
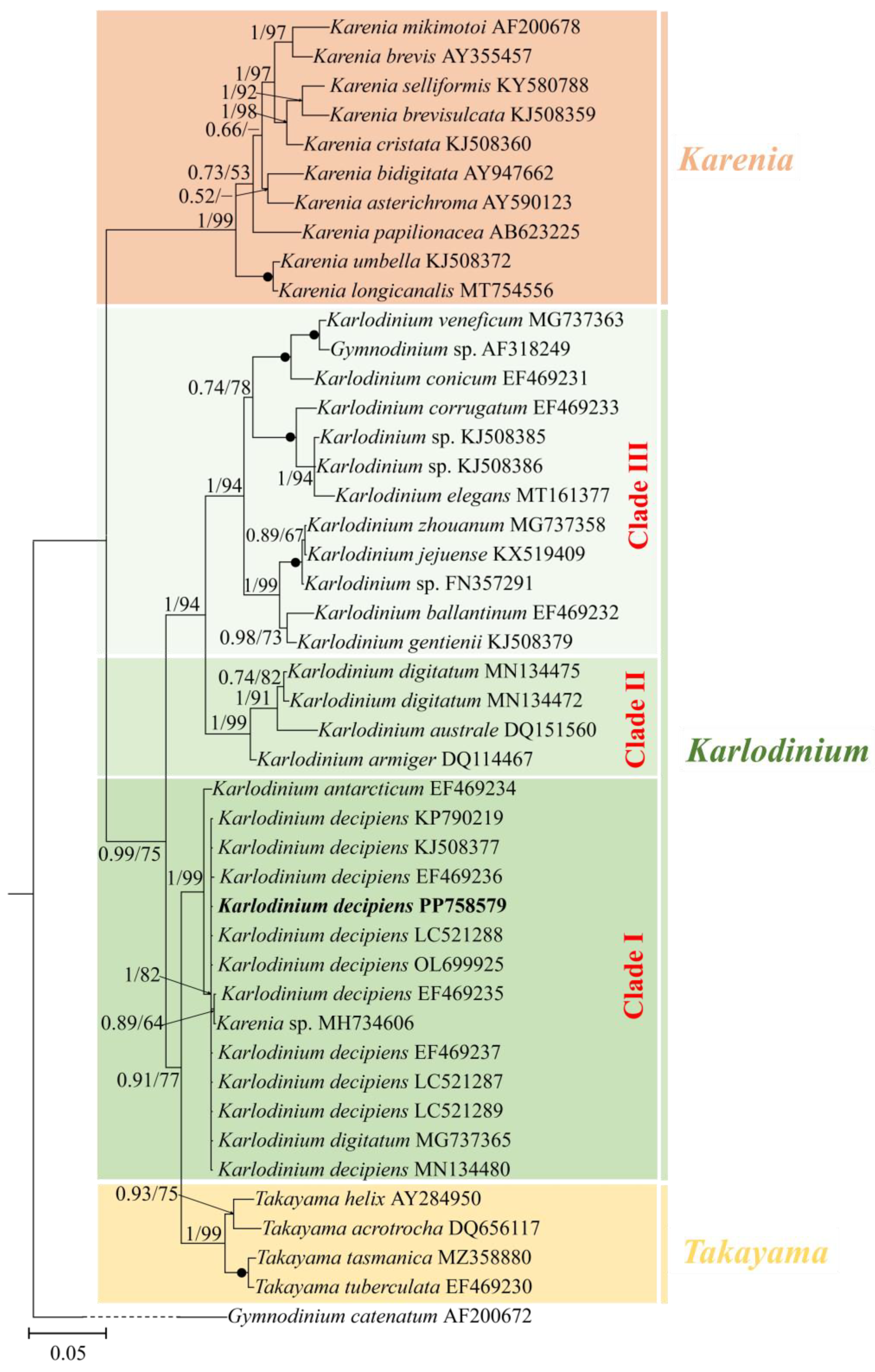
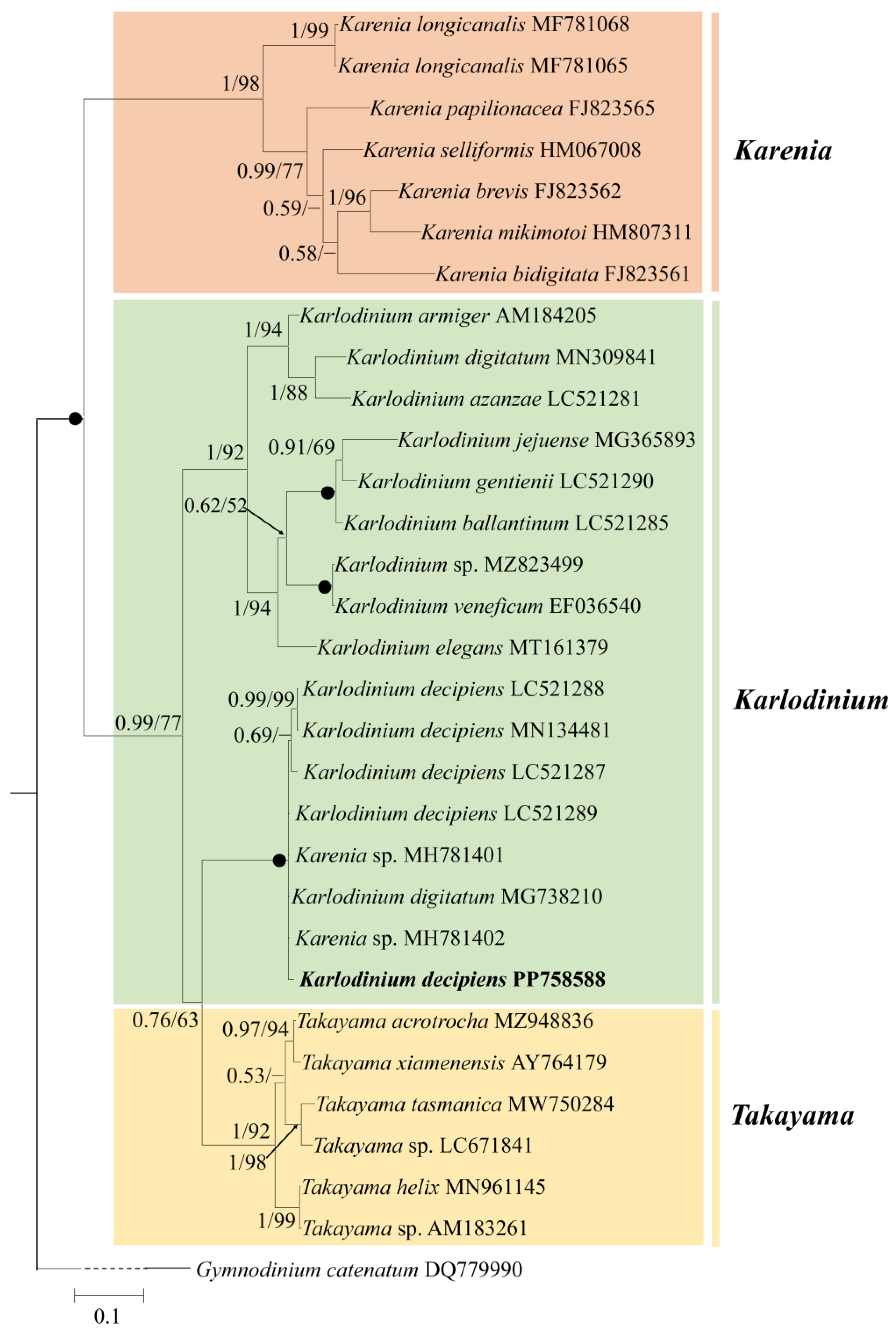
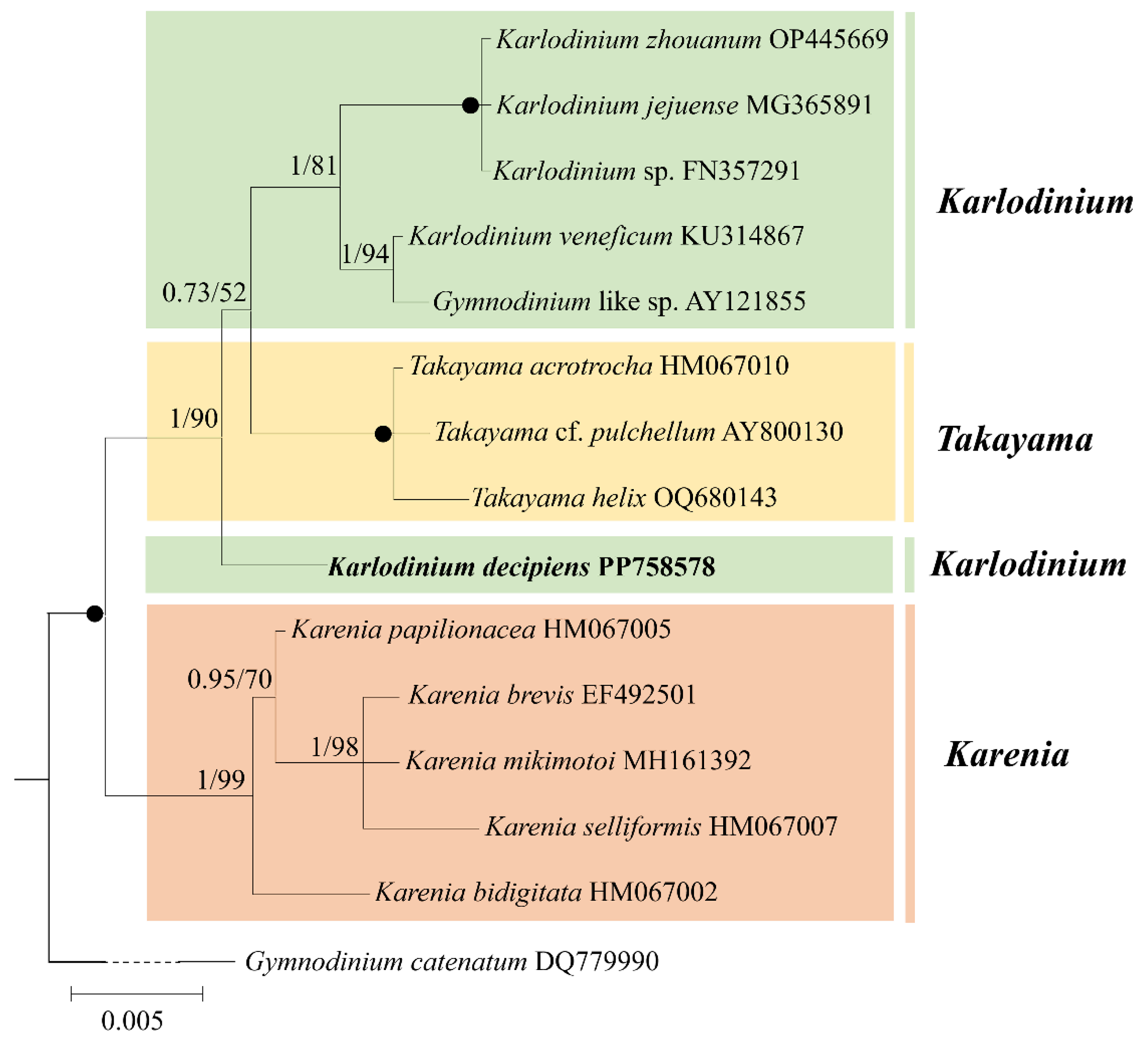
3.2. Phylogeny
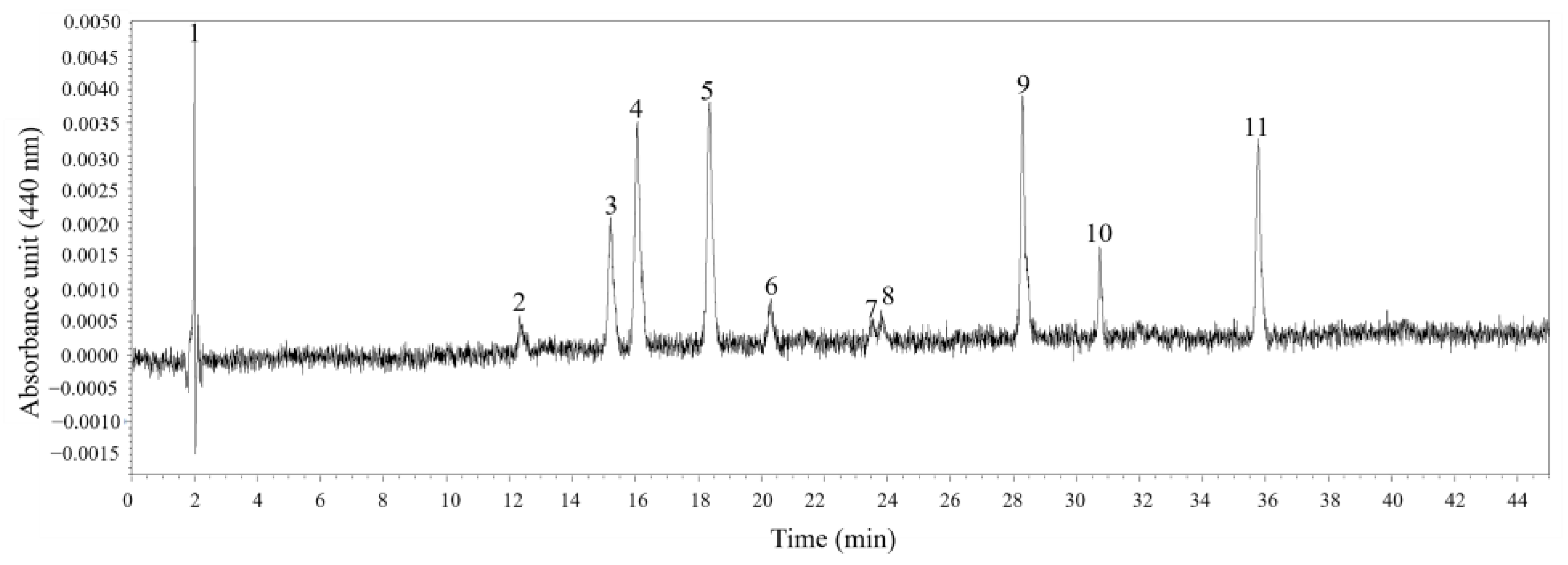
3.3. Pigment
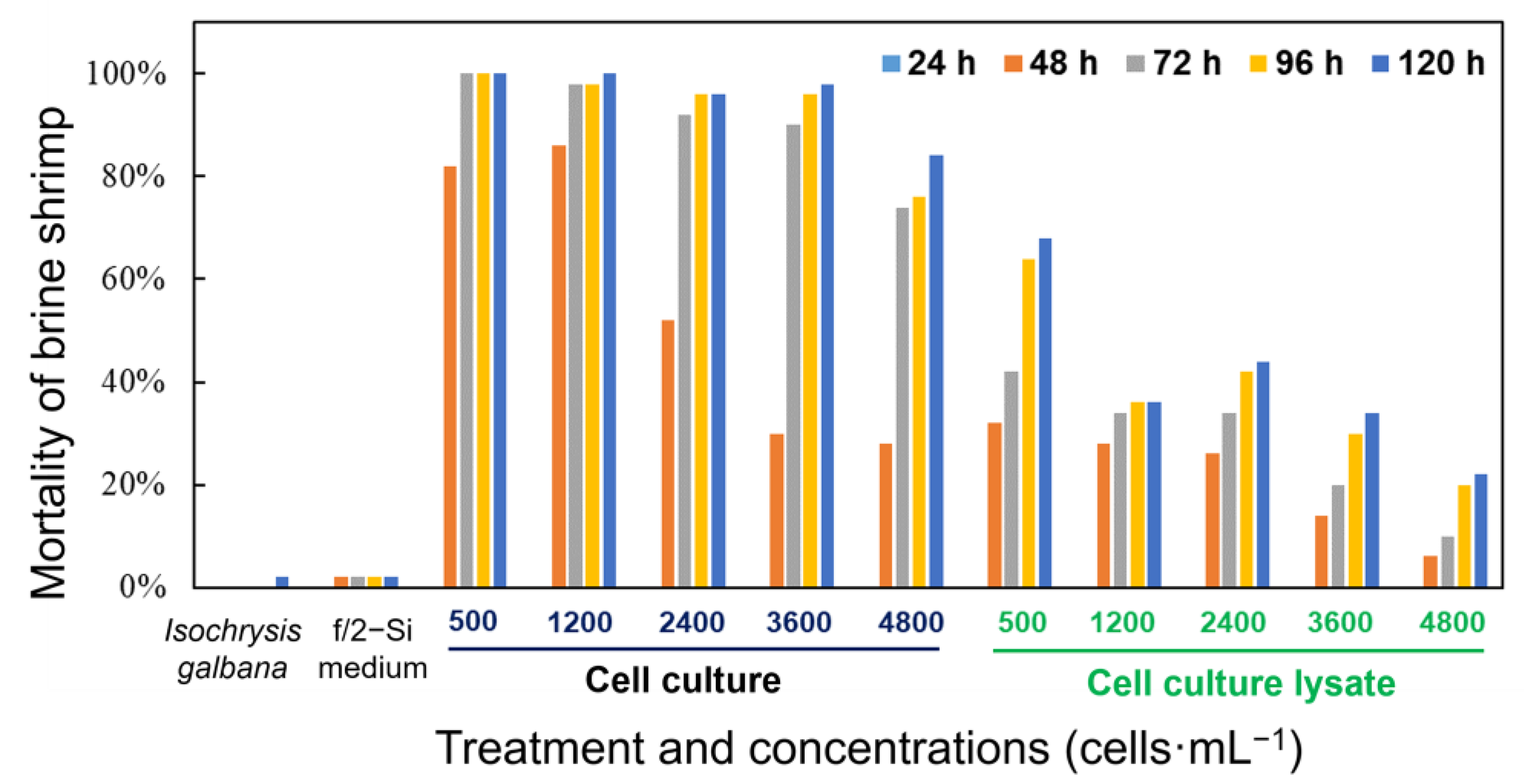
3.4. Toxicity of Karlodinium decipiens to Brine Shrimp
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez, F. Diversity and Classification of Dinoflagellates. In Dinoflagellates: Classification, Evolution, Physiology, and Ecological Significance; Rao, D.V.S., Ed.; Nova Science Publisher: New York, NY, USA, 2020; p. 38. [Google Scholar]
- Taylor, F.J.R.; Hoppenrath, M.; Saldarriaga, J.F. Dinoflagellate diversity and distribution. Biodivers. Conserv. 2008, 17, 407–418. [Google Scholar] [CrossRef]
- Morse, D.; Tse, S.P.K.; Lo, S.C.L. Exploring dinoflagellate biology with high-throughput proteomics. Harmful Algae 2018, 75, 16–26. [Google Scholar] [CrossRef]
- Taylor, F.J.R. Dinoflagellate Morphology. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell Scientific Publications: Oxford, UK, 1987; pp. 24–91. [Google Scholar]
- Fensome, R.A.; Taylor, F.J.R.; Norris, G.; Sargeant, W.A.S.; Wharton, D.I.; Williams, G.L. A Classification of Living and Fossil Dinoflagellates; Sheridan Press: Hanover, PA, USA, 1993; Volume 7, pp. 1–351. [Google Scholar]
- Kofoid, C.A.; Swezy, O. The Free-Living Unarmored Dinoflagellata; University of California Press: Berkeley, CA, USA, 1921; Volume 5, p. 559. [Google Scholar]
- Takayama, H. Apical grooves of unarmored dinoflagellates. Bull. Plankton Soc. Jpn. 1985, 32, 129–140. [Google Scholar]
- Daugbjerg, N.; Hansen, G.; Larsen, J.; Moestrup, Ø. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–317. [Google Scholar] [CrossRef]
- de Salas, M.F.; Bolch, C.J.S.; Botes, L.; Nash, G.; Wright, S.W.; Hallegraeff, G.M. Takayama gen. nov. (Gymnodiniales, Dinophyceae), A new genus of unarmored dinoflagellates with sigmoid apical grooves, including the description of two new species. J. Phycol. 2003, 39, 1233–1246. [Google Scholar] [CrossRef]
- Moestrup, Ø.; Hakanen, P.; Hansen, G.; Daugbjerg, N.; Ellegaard, M. On Levanderina fissa gen. & comb. nov. (Dinophyceae) (syn. Gymnodinium fissum, Gyrodinium instriatum, Gyr. uncatenum), a dinoflagellate with a very unusual sulcus. Phycologia 2014, 53, 265–292. [Google Scholar] [CrossRef]
- Hoppenrath, M. Dinoflagellate taxonomy—A review and proposal of a revised classification. Mar. Biodivers. 2017, 47, 381–403. [Google Scholar] [CrossRef]
- Gao, M.; Hu, Z.; Luo, Z.; Deng, Y.; Shang, L.; Sun, Y.; Tang, Y. Morphological and molecular characterization of the unarmored dinoflagellate Gymnodinium trapeziforme (Dinophyceae) from Jiaozhou Bay, China. Diversity 2023, 15, 1186. [Google Scholar] [CrossRef]
- Hu, Z.; Deng, Y.; Li, Y.; Tang, Y.Z. The morphological and phylogenetic characterization for the dinoflagellate Margalefidinium fulvescens (=Cochlodinium fulvescens) isolated from the Jiaozhou Bay, China. Acta Oceanol. Sin. 2018, 37, 11–17. [Google Scholar] [CrossRef]
- Hu, Z.; Deng, Y.; Luo, Z.; Shang, L.; Kong, F.; Gu, H.; Zhao, Z.; Tang, Y.Z. Characterization of the unarmored dinoflagellate Pseliodinium pirum (Ceratoperidiniaceae) from Jiaozhou Bay, China. Phycol. Res. 2020, 68, 3–13. [Google Scholar] [CrossRef]
- Hu, Z.; Li, Z.; Deng, Y.; Iwataki, M.; Luo, Z.; Wang, J.; Sun, Y.; Zhao, Z.; Gu, H.; Shin, H.H.; et al. Morphology, ultrastructure, and molecular phylogeny of the unarmoured dinoflagellate Kirithra sigma sp. nov. (Ceratoperidiniaceae, Dinophyceae). Phycologia 2020, 59, 385–396. [Google Scholar] [CrossRef]
- Hu, Z.; Xu, N.; Gu, H.; Chai, Z.; Takahashi, K.; Li, Z.; Deng, Y.; Iwataki, M.; Matsuoka, K.; Tang, Y.Z. Morpho-molecular description of a new HAB species, Pseudocochlodinium profundisulcus gen. et sp. nov., and its LSU rRNA gene based genetic diversity and geographical distribution. Harmful Algae 2021, 108, 102098. [Google Scholar] [CrossRef] [PubMed]
- Boutrup, P.V.; Moestrup, Ø.; Tillmann, U.; Daugbjerg, N. Ultrastructure and phylogeny of Kirithra asteri gen. et sp. nov. (Ceratoperidiniaceae, Dinophyceae)—A free-living, thin-walled marine photosynthetic dinoflagellate from Argentina. Protist 2017, 168, 586–611. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F. A checklist and classification of living dinoflagellates (Dinoflagellata, Alveolata). CICIMAR Ocean. 2012, 27, 65–140. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? A reprise. Four kingdoms, 14 phyla, 63 classes and still growing. J. Phycol. 2024, 60, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, Y.; Chai, Z.; Hu, Z.; Tang, Y.Z. A combined approach detected novel species diversity and distribution of dinoflagellate cysts in the Yellow Sea, China. Mar. Pollut. Bull. 2023, 187, 114567. [Google Scholar] [CrossRef] [PubMed]
- Djurhuus, A.; Closek, C.J.; Kelly, R.P.; Pitz, K.J.; Michisaki, R.P.; Starks, H.A.; Walz, K.R.; Andruszkiewicz, E.A.; Olesin, E.; Hubbard, K.; et al. Environmental DNA reveals seasonal shifts and potential interactions in a marine community. Nat. Commun. 2020, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.F.; Horak, A.; Andrew, R.L.; Coffroth, M.-A.; Andersen, R.A.; Küpper, F.C.; Jameson, I.; Hoppenrath, M.; Véron, B.; Kasai, F.; et al. Environmental barcoding reveals massive dinoflagellate diversity in marine environments. PLoS ONE 2010, 5, e13991. [Google Scholar] [CrossRef]
- Sournia, A. Quatre nouveaux dinoflagellés du plancton marin. Phycologia 1972, 11, 71–74. [Google Scholar] [CrossRef]
- Takahashi, K.; Benico, G.; Lum, W.M.; Iwataki, M. Gertia stigmatica gen. et sp. nov. (Kareniaceae, Dinophyceae), a new marine unarmored dinoflagellate possessing the peridinin-type chloroplast with an eyespot. Protist 2019, 170, 125680. [Google Scholar] [CrossRef]
- Ok, J.H.; Jeong, H.J.; Lee, S.Y.; Park, S.A.; Noh, J.H. Shimiella gen. nov. and Shimiella gracilenta sp. nov. (Dinophyceae, Kareniaceae), a kleptoplastidic dinoflagellate from Korean waters and its survival under starvation. J. Phycol. 2021, 57, 70–91. [Google Scholar] [CrossRef]
- Bergholtz, T.; Daugbjerg, N.; Moestrup, Ø.; Fernández-Tejedor, M. On the identity of Karlodinium veneficum and description of Karlodinium armiger sp. nov. (Dinophyceae), based on light and electron microscopy, nuclear-encoded LSU rDNA, and pigment composition. J. Phycol. 2006, 42, 170–193. [Google Scholar] [CrossRef]
- Cen, J.; Lu, S.; Moestrup, Ø.; Jiang, T.; Ho, K.C.; Li, S.; Li, M.; Huan, Q.; Wang, J. Five Karenia species along the Chinese coast: With the description of a new species, Karenia hui sp. nov. (Kareniaceae, Dinophyta). Harmful Algae 2024, 137, 102645. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: https://www.algaebase.org/browse/taxonomy/#91756 (accessed on 21 May 2024).
- Yang, H.; Hu, Z.; Tang, Y.Z. Plasticity and multiplicity of trophic modes in the dinoflagellate Karlodinium and their pertinence to population maintenance and bloom dynamics. J. Mar. Sci. Eng. 2021, 9, 51. [Google Scholar] [CrossRef]
- Place, A.R.; Bowers, H.A.; Bachvaroff, T.R.; Adolf, J.E.; Deeds, J.R.; Sheng, J. Karlodinium veneficum—The little dinoflagellate with a big bite. Harmful Algae 2012, 14, 179–195. [Google Scholar] [CrossRef]
- Lim, H.C.; Leaw, C.P.; Tan, T.H.; Kon, N.F.; Yek, L.H.; Hii, K.S.; Teng, S.T.; Razali, R.M.; Usup, G.; Iwataki, M.; et al. A bloom of Karlodinium australe (Gymnodiniales, Dinophyceae) associated with mass mortality of cage-cultured fishes in West Johor Strait, Malaysia. Harmful Algae 2014, 40, 51–62. [Google Scholar] [CrossRef]
- Song, X.; Hu, Z.; Shang, L.; Leaw, C.P.; Lim, P.T.; Tang, Y.Z. Contact micropredation may play a more important role than exotoxicity does in the lethal effects of Karlodinium australe blooms: Evidence from laboratory bioassays. Harmful Algae 2020, 99, 101926. [Google Scholar] [CrossRef]
- Yang, H.; Hu, Z.; Shang, L.; Deng, Y.; Tang, Y.Z. A strain of the toxic dinoflagellate Karlodinium veneficum isolated from the East China Sea is an omnivorous phagotroph. Harmful Algae 2020, 93, 101775. [Google Scholar] [CrossRef]
- Cen, J.; Wang, J.; Huang, L.; Ding, G.; Qi, Y.; Cao, R.; Cui, L.; Lü, S. Who is the “murderer” of the bloom in coastal waters of Fujian, China, in 2019? J. Oceanol. Limnol. 2020, 38, 722–732. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, L.; Chan, L.; Lu, S.; Gu, H. Karlodinium zhouanum, a new dinoflagellate species from China, and molecular phylogeny of Karenia digitata and Karenia longicanalis (Gymnodiniales, Dinophyceae). Phycologia 2018, 57, 401–412. [Google Scholar] [CrossRef]
- Li, Z.; Shin, H.H. Morphology and phylogeny of an unarmored dinoflagellate, Karlodinium jejuense sp. nov. (Gymnodiniales), isolated from the northern East China Sea. Phycol. Res. 2018, 66, 318–328. [Google Scholar] [CrossRef]
- Cen, J.; Wang, J.; Huang, L.; Lin, Y.; Ding, G.; Qi, Y.; Lü, S. Karlodinium elegans sp. nov. (Gymnodiniales, Dinophyceae), a novel species isolated from the East China Sea in a dinoflagellate bloom. J. Oceanol. Limnol. 2021, 39, 242–258. [Google Scholar] [CrossRef]
- Benico, G.; Takahashi, K.; Lum, W.M.; Yñiguez, A.T.; Iwataki, M. The harmful unarmored dinoflagellate Karlodinium in Japan and Philippines, with reference to ultrastructure and micropredation of Karlodinium azanzae sp. nov. (Kareniaceae, Dinophyceae). J. Phycol. 2020, 56, 1264–1282. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Sun, X.; Feng, J.; Yan, X. Microscopic observations and molecular identification of toxic unarmoured dinoflagellates Karlodinium micrum (Dinophyceae) from the East China Sea (ECS). Mar. Sci. Bull. 2008, 27, 32–37. [Google Scholar]
- Liu, Y.; Hu, Z.; Deng, Y.; Tang, Y.Z. Evidence for resting cyst production in the cosmopolitan toxic dinoflagellate Karlodinium veneficum and the cyst distribution in the China seas. Harmful Algae 2020, 93, 101788. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals: Proceedings—1st Conference on Culture of Marine Invertebrate Animals Greenport; Smith, W.L., Chanley, M.H., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Medlin, L.K.; Elwood, H.J.; Stickel, S.K.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of MAFFT for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Wang, J.; Kong, F.; Geng, H.; Zhao, Y.; Guan, W.; He, C.; Kang, Z.; Guo, W.; Zhou, Z.; Zhang, Q.; et al. Pigment characterization of the giant-colony-forming Haptophyte Phaeocystis globosa in the Beibu Gulf reveals blooms of different origins. Appl. Environ. Microbiol. 2022, 88, e01654-21. [Google Scholar] [CrossRef]
- Kong, F.; Yu, R.; Zhang, Q.; Yan, T.; Zhou, M. Pigment characterization for the 2011 bloom in Qinhuangdao implicated “brown tide” events in China. Chin. J. Oceanol. Limnol. 2012, 30, 361–370. [Google Scholar] [CrossRef]
- de Salas, M.F.; Laza-Martínez, A.; Hallegraeff, G.M. Novel unarmored dinoflagellates from the toxigenic family Kareniaceae (Gymnodiniales): Five new species of Karlodinium and one new Takayama from the australian sector of the Southern Ocean. J. Phycol. 2008, 44, 241–257. [Google Scholar] [CrossRef]
- Ballantine, D. Two new marine species of Gymnodinium isolated from the Plymouth area. J. Mar. Biol. Assoc. UK 1956, 35, 467–474. [Google Scholar] [CrossRef]
- de Salas, M.F.; Bolch, C.J.S.; Hallegraeff, G.M. Karlodinium australe sp. nov. (Gymnodiniales, Dinophyceae), a new potentially ichthyotoxic unarmoured dinoflagellate from lagoonal habitats of south-eastern Australia. Phycologia 2005, 44, 640–650. [Google Scholar] [CrossRef]
- Siano, R.; Kooistra, W.H.C.F.; Montresor, M.; Zingone, A. Unarmoured and thin-walled dinoflagellates from the Gulf of Naples, with the description of Woloszynskia cincta sp. nov. (Dinophyceae, Suessiales). Phycologia 2009, 48, 44–65. [Google Scholar] [CrossRef]
- Nézan, E.; Siano, R.; Boulben, S.; Six, C.; Bilien, G.; Chèze, K.; Duval, A.; Le Panse, S.; Quéré, J.; Chomérat, N. Genetic diversity of the harmful family Kareniaceae (Gymnodiniales, Dinophyceae) in France, with the description of Karlodinium gentienii sp. nov.: A new potentially toxic dinoflagellate. Harmful Algae 2014, 40, 75–91. [Google Scholar] [CrossRef]
- Gu, H.; Wu, Y.; Lü, S.; Lu, D.; Tang, Y.Z.; Qi, Y. Emerging harmful algal bloom species over the last four decades in China. Harmful Algae 2021, 102059. [Google Scholar] [CrossRef]
- Pan, Y.; Meng, R.; Li, Y.; Yang, L.; Mei, L.; Wu, Y.; Xu, J.; Zhou, C.; Yan, X. Changes in biochemical metabolites in manila clam after a temporary culture with high-quality microalgal feed mixed with the dinoflagellate species Karlodinium veneficum and K. zhouanum. Harmful Algae 2023, 125, 102422. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigment | Pigment Concentration (pg/cell) | Ratio to Chl a (w/w) |
|---|---|---|
| 19-butanoyloxyfucoxanthin | 0.93 | 0.29 |
| Fucoxanthin | 1.99 | 0.63 |
| 19′-Hexanoyloxyfucoxanthin | 1.73 | 0.55 |
| Diadinoxanthin | 0.28 | 0.09 |
| Diatoxanthin | 0.14 | 0.04 |
| Chl a | 3.16 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, J.; Fan, Y.; Chen, Q.; Chen, X.; Deng, Y.; Sun, Y.; Wang, J.; Hu, Z.; Tang, Y.Z. Characterization of the Unarmored Dinoflagellate Karlodinium decipiens (Dinophyceae) from Jiaozhou Bay, China. Diversity 2024, 16, 449. https://doi.org/10.3390/d16080449
Yao J, Fan Y, Chen Q, Chen X, Deng Y, Sun Y, Wang J, Hu Z, Tang YZ. Characterization of the Unarmored Dinoflagellate Karlodinium decipiens (Dinophyceae) from Jiaozhou Bay, China. Diversity. 2024; 16(8):449. https://doi.org/10.3390/d16080449
Chicago/Turabian StyleYao, Jialin, Yingyi Fan, Qiantong Chen, Xinxin Chen, Yunyan Deng, Yuanyuan Sun, Jinxiu Wang, Zhangxi Hu, and Ying Zhong Tang. 2024. "Characterization of the Unarmored Dinoflagellate Karlodinium decipiens (Dinophyceae) from Jiaozhou Bay, China" Diversity 16, no. 8: 449. https://doi.org/10.3390/d16080449
APA StyleYao, J., Fan, Y., Chen, Q., Chen, X., Deng, Y., Sun, Y., Wang, J., Hu, Z., & Tang, Y. Z. (2024). Characterization of the Unarmored Dinoflagellate Karlodinium decipiens (Dinophyceae) from Jiaozhou Bay, China. Diversity, 16(8), 449. https://doi.org/10.3390/d16080449