
Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities


Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subgenus | Group | Subgroup | Species | Subspecies |
|---|---|---|---|---|
| T. (Microtrechus) | T. vandykei | T. barberi | T. subtilis Barr, 1962 * | |
| T. pseudosubtilis Donabauer, 2009 * | ||||
| T. tonitru Barr, 1962 | ||||
| T. barberi Jeannel, 1931 * | ||||
| T. pseudobarberi Donabauer, 2009 * | ||||
| T. bowlingi | T. bowlingi Barr, 1962 * | |||
| T. vandykei | T. tusquitee Barr, 1979 | |||
| T. pisgahensis Barr, 1979 * | ||||
| T. vandykei Jeannel, 1927 * | ||||
| T. haoe Barr, 1979 * | ||||
| T. uncifer | T. inexpectatus Barr, 1985 * | |||
| T. thunderheadensis Donabauer, 2005 * | ||||
| T. uncifer Barr, 1962 | ||||
| T. plottbalsamensis Donabauer, 2005 * | ||||
| T. aduncus Barr, 1962 * | ||||
| T. cowensis Barr, 1962 | ||||
| T. toxawayi Barr, 1979 * | ||||
| T. howellae Barr, 1979 | ||||
| T. satanicus Barr, 1962 * | ||||
| T. tusquitensis Donabauer, 2005 * | ||||
| T. talequah Barr, 1962 * | ||||
| T. nebulosus | T. balsamensis Barr, 1962 * | |||
| T. cheoahensis Donabauer, 2005 | ||||
| T. clingmanensis Donabauer, 2005 * | ||||
| T. haoeleadensis Donabauer, 2005 | ||||
| T. luculentus | T. l. cheoahbaldensis Donabauer, 2005 | |||
| T. l. joannabaldensis Donabauer, 2005 * | ||||
| T. l. luculentus Barr, 1962 * | ||||
| T. l. wayahensis Barr, 1979 | ||||
| T. nantahalae Barr, 1979 | ||||
| T. nebulosus Barr, 1962 * | ||||
| T. novaculosus Barr, 1962 * | ||||
| T. pseudonovaculosus Donabauer, 2005 | ||||
| T. ramseyensis Donabauer, 2005 * | ||||
| T. rosenbergi Barr, 1962 | ||||
| T. snowbirdensis Donabauer, 2005 | ||||
| T. stefanschoedli Donabauer, 2005 | ||||
| T. stupkai Barr, 1979 | ||||
| T. tennesseensis | T. t. tauricus Barr, 1962 * | |||
| T. t. tennesseensis Barr, 1962 * | ||||
| T. thomasbarri Donabauer, 2005 | ||||
| T. tobiasi Donabauer, 2005 * | ||||
| T. tuckaleechee Barr, 1962 * | ||||
| T. unicoi Barr, 1962 | ||||
| T. valentinei Barr, 1979 | ||||
| T. verus Barr, 1962 * | ||||
| T. wayahbaldensis Donabauer, 2005 * | ||||
| T. (Trechus) | T. hydropicus | T. hydropicus | T. h. hydropicus (Horn, 1883) * | |
| T. h. avus Barr, 1962 * | ||||
| T. h. beutenmuelleri Jeannel, 1931 * | ||||
| T. h. canus Barr Barr, 1962 * | ||||
| T. schwarzi | T. s. schwarzi Jeannel, 1931 | |||
| T. s. scopulosus Barr, 1979 * | ||||
| T. s. saludae Barr, 1979 * | ||||
| T. cumberlandus Barr, 1962 * | ||||
| T. mitchellensis Barr, 1962 * | ||||
| T. carolinae Schaeffer, 1901 * | ||||
| T. roanicus Barr, 1962 * | ||||
| T. calignis Barr, 1985 * |
2. Materials and Methods
2.1. Specimen Collection and Identification
2.2. DNA Extraction and Amplification
2.3. Tree Building
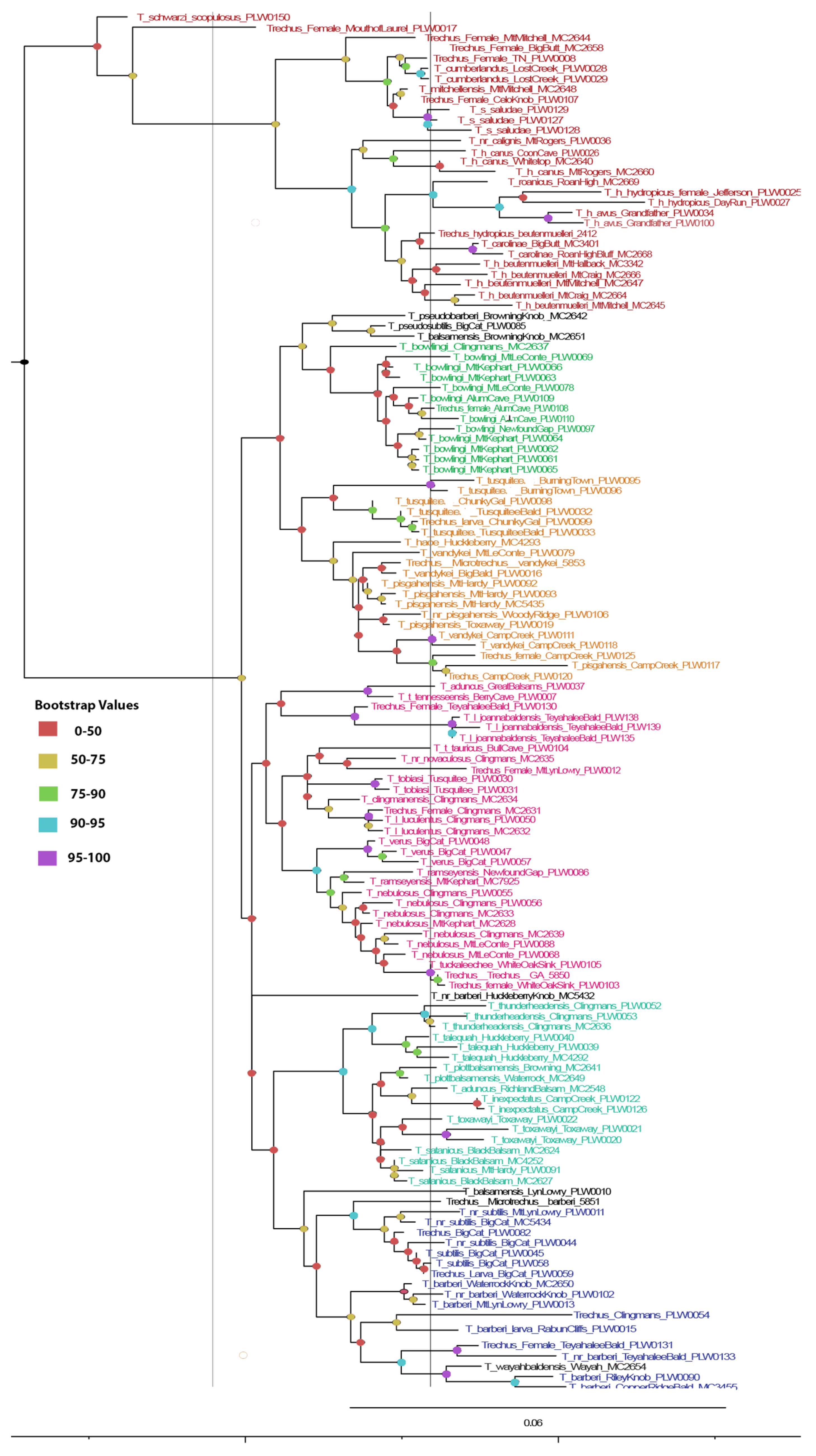
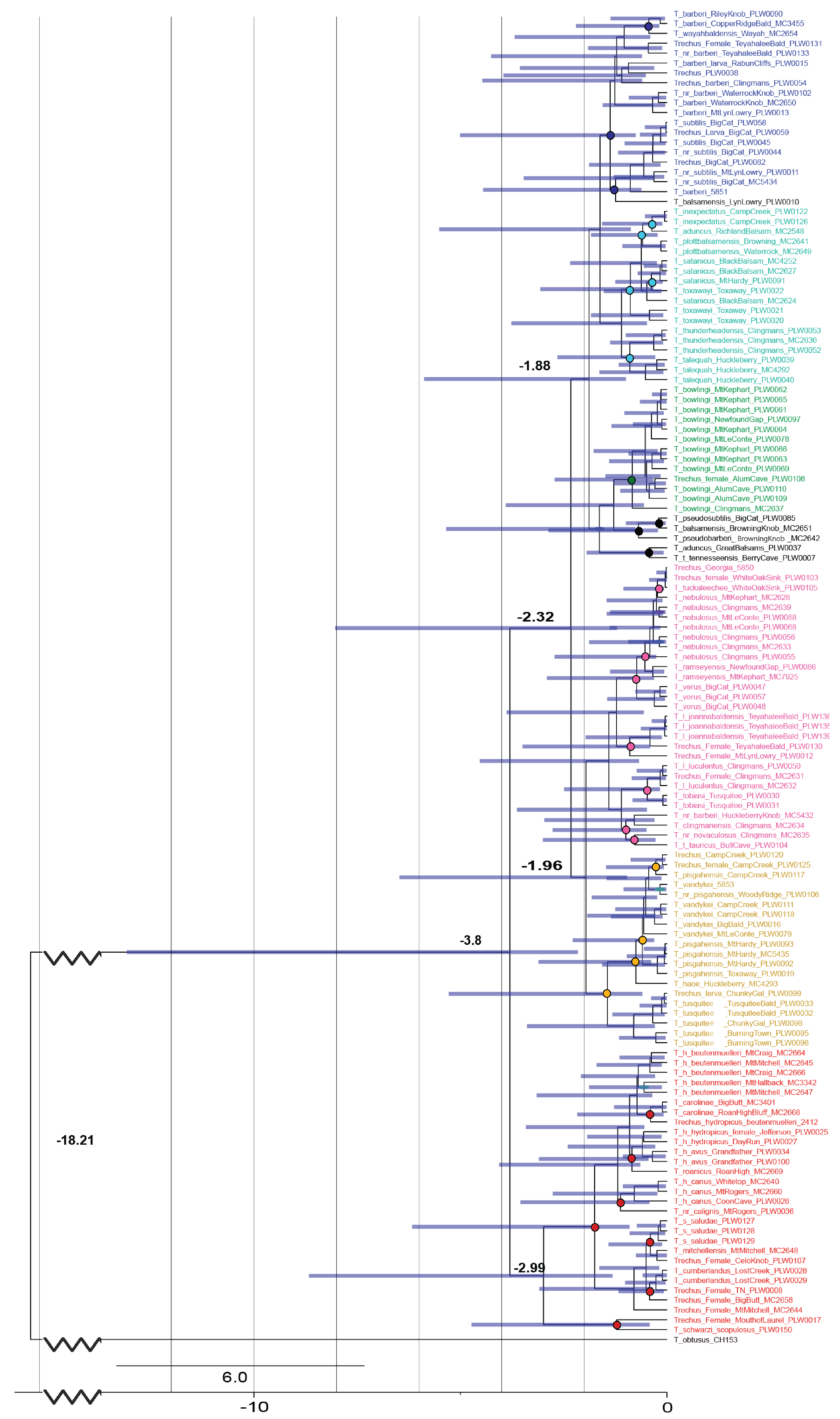
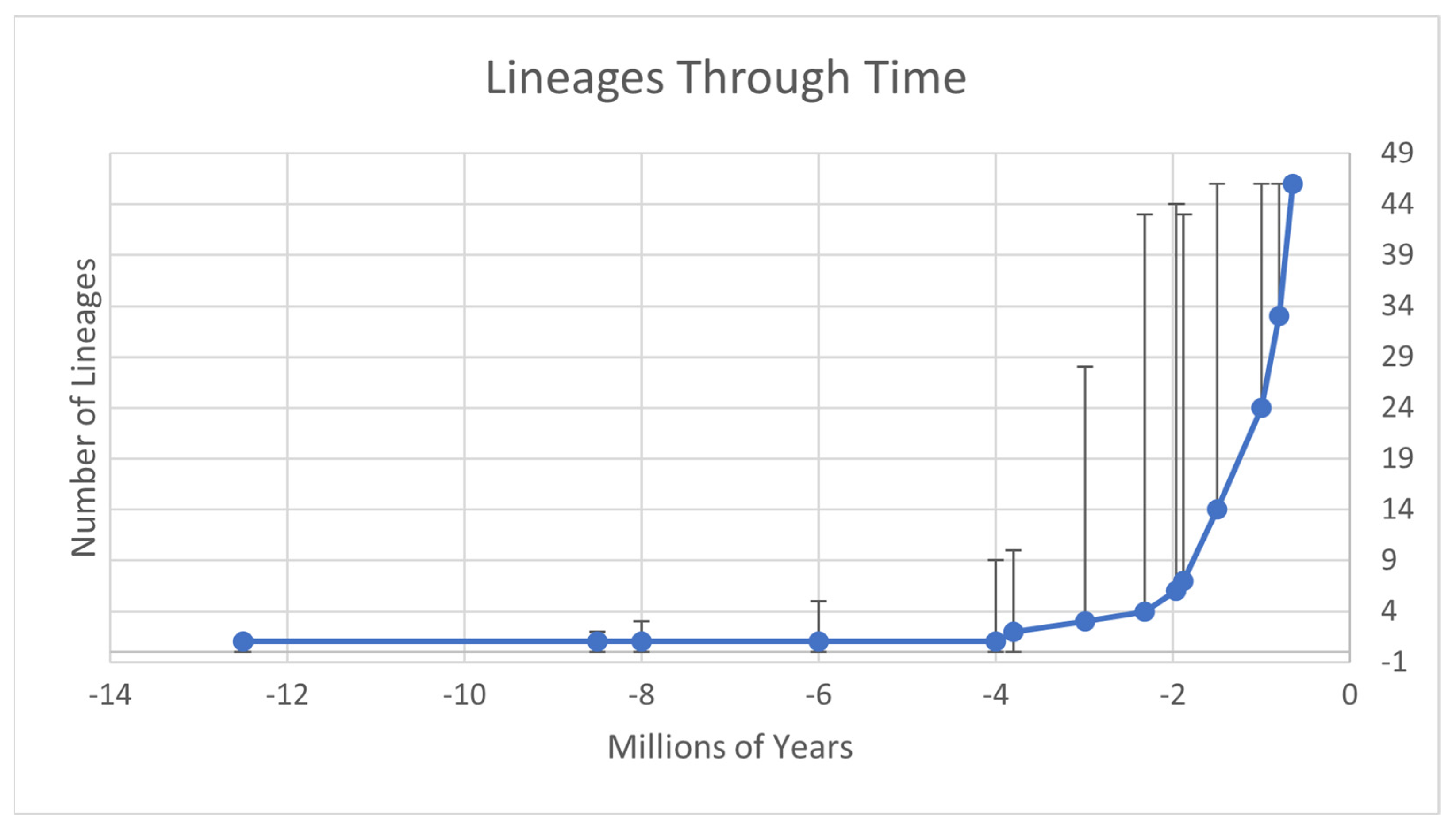
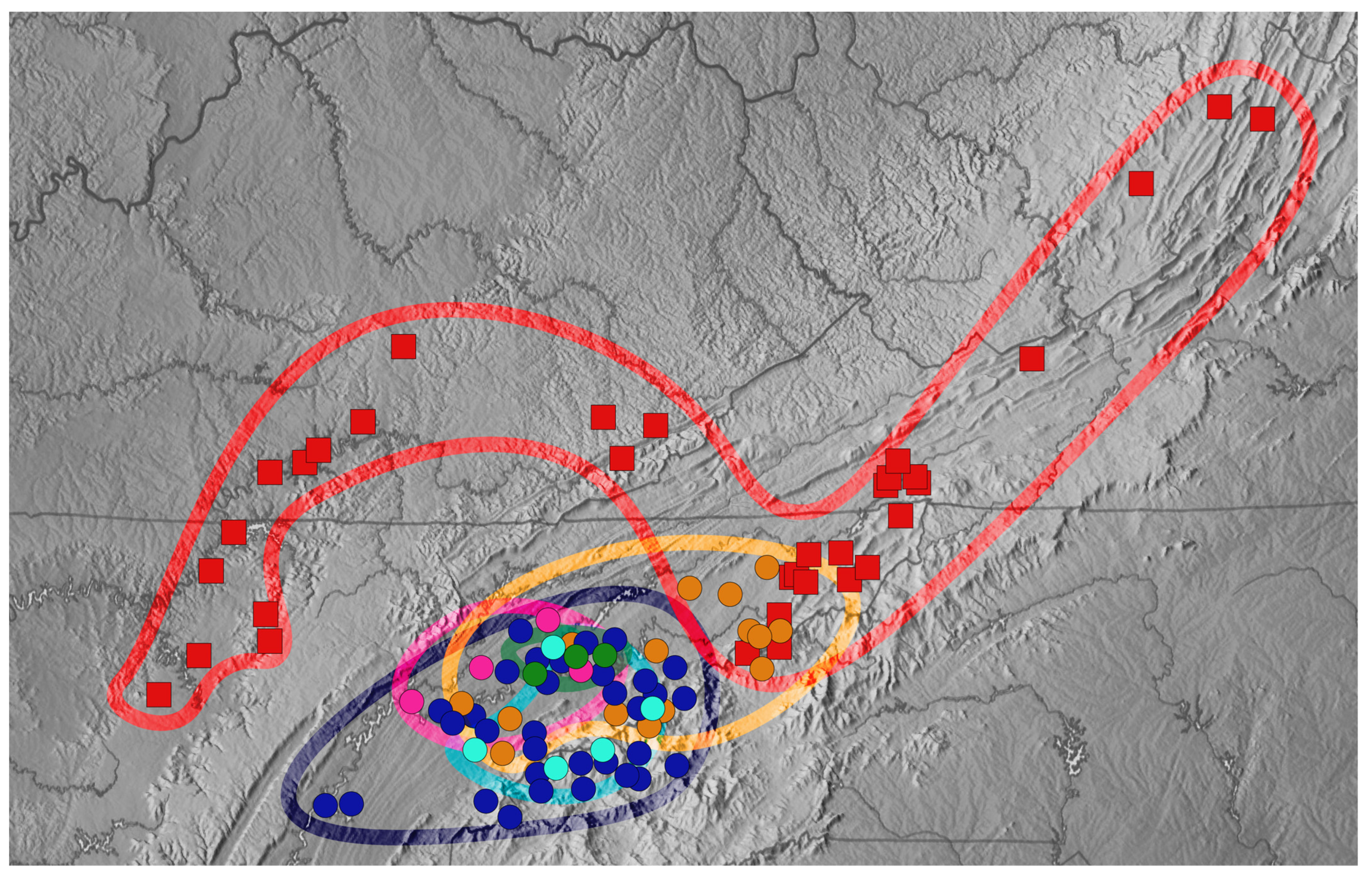
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yarnell, S.L. The Southern Appalachians: A History of the Landscape; US Department of Agriculture Forest Service: Asheville, NC, USA, 1998. [Google Scholar]
- Delcourt, P.A.; Delcourt, H.R. Paleoecological Insights on Conservation of Biodiversity: A Focus on Species, Ecosystems, and Landscapes. Ecol. Appl. 2017, 8, 921–934. [Google Scholar] [CrossRef]
- Hedin, M.; McCormack, M. Biogeographical evidence for common vicariance and rare dispersal in a southern Appalachian harvestman (Sabaconidae, Sabacon cavicolens). J. Biogeogr. 2017, 44, 1665–1678. [Google Scholar] [CrossRef]
- Thomas, S.M.; Hedin, M. Multigenic phylogeographic divergence in the paleoendemic southern Appalachian opilionid Fumontana deprehendor Shear (Opiliones, Laniatores, Triaenonychidae). Mol. Phylogen. Evol. 2008, 46, 645–658. [Google Scholar] [CrossRef]
- Church, S.A.; Kraus, J.M.; Mitchell, J.C.; Church, D.R.; Taylor, D.R. Evidence for multiple pleistocene refugia in the postglacial expansion of the eastern tiger salamander, Ambystoma tigrinum tigrinum. Evolution 2008, 57, 372–383. [Google Scholar] [CrossRef]
- Walker, M.J.; Stockman, A.K.; Marek, P.E.; Bond, J.E. Pleistocene glacial refugia across the appalachian mountains and coastal plain in the millipede genus Narceus: Evidence from population genetic, phylogeographic, and paleoclimatic data. BMC Evol. Biol. 2009, 9, 25. [Google Scholar] [CrossRef]
- Wheeler, Q. Notes on Host Associations and Habitats of Dasyceridae (Coleoptera) in the Southern Appalachian Mountains. Coleopt. Bull. 1984, 38, 227–231. [Google Scholar]
- Estes, D. Carex fumosimontana (Cyperaceae), a new endemic from the Great Smoky Mountains National Park, North Carolina and Tennessee. Brittonia 2013, 65, 200–207. [Google Scholar] [CrossRef]
- Dirnböck, T.; Essl, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Mccain, C.M.; Colwell, R.K. Assessing the threat to montane biodiversity from discordant shifts in temperature and precipitation in a changing climate. Ecol. Lett. 2011, 14, 1236–1245. [Google Scholar] [CrossRef]
- Wiens, J.J.; Camacho, A.; Goldberg, A.; Jezkova, T.; Kaplan, M.E.; Lambert, S.M.; Miller, E.C.; Streicher, J.W.; Walls, R.L. Climate change, extinction, and Sky Island biogeography in a montane lizard. Mol. Ecol. 2019, 28, 2610–2624. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.C. The Pseudanophthalmus of the Appalachian Valley (Coleoptera: Carabidae). Am. Midl. Nat. 1965, 73, 41–72. [Google Scholar] [CrossRef]
- Hedin, M.; Milne, M.A. New species in old mountains: Integrative taxonomy reveals ten new species and extensive short-range endemism in Nesticus spiders (Araneae, Nesticidae) from the southern Appalachian Mountains. ZooKeys 2023, 1145, 1. [Google Scholar] [CrossRef]
- Graham, C.H.; Carnaval, A.C.; Cadena, C.D.; Zamudio, K.R.; Roberts, T.E.; Parra, J.L.; Mccain, C.M.; Bowie, R.C.K.; Moritz, C.; Baines, S.B.; et al. The origin and maintenance of montane diversity: Integrating evolutionary and ecological processes. Ecography 2014, 37, 711–719. [Google Scholar] [CrossRef]
- Caterino, M.S.; Recuero, E. Shedding light on dark taxa in sky-island Appalachian leaf litter: Assessing patterns of endemicity using large-scale, voucher-based barcoding. Insect Conserv. Divers. 2023, 17, 16–30. [Google Scholar] [CrossRef]
- Jenkins, M.A. Impact of the Balsam Woolly Adelgid (Adelges piceae Ratz.) on an Abies fraseri (Pursh) Poir. Dominated Stand near the Summit of Mount LeConte, Tennessee. Castanea 2003, 68, 109–118. [Google Scholar]
- McManamay, R.H.; Resler, L.M.; Campbell, J.B.; McManamay, R.A. Assessing the impacts of balsam woolly adelgid (Adelges piceae Ratz.) and anthropogenic disturbance on the stand structure and mortality of Fraser fir [Abies fraseri (pursh) Poir.] in the Black Mountains, North Carolina. Castanea 2011, 76, 1–19. [Google Scholar] [CrossRef]
- Dukes, C.D.; Janssens, F.; Recuero, E.; Caterino, M.S. Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity 2022, 14, 847. [Google Scholar] [CrossRef]
- Caterino, M.S.; Recuero, E. Molecular diversity of Protura in southern High Appalachian leaf litter. Biodivers. Data J. 2023, 11, e113342. [Google Scholar] [CrossRef]
- Graham, M.R.; Flint, W.D.; Powell, A.M.; Fet, V.; Pauley, T.K. Phylogeography of the Cow Knob Salamander (Plethodon punctatus Highton): Populations on isolated Appalachian mountaintops are disjunct but not divergent. Front. Amphib. Reptile Sci. 2023, 1, 1175492. [Google Scholar] [CrossRef]
- Herman, T.A.; Bouzat, J.L. Range-wide phylogeography of the four-toed salamander: Out of Appalachia and into the glacial aftermath. J. Biogeogr. 2016, 43, 666–678. [Google Scholar] [CrossRef]
- Hedin, M.; Carlson, D.; Coyle, F. Sky island diversification meets the multispecies coalescent—Divergence in the spruce-fir moss spider (Microhexura montivaga, Araneae, Mygalomorphae) on the highest peaks of southern Appalachia. Mol. Ecol. 2015, 24, 3467–3484. [Google Scholar] [CrossRef]
- Ortiz, C.C.; Browne, R.A. Carabidae (ground beetle) species composition of southern Appalachian spruce-fir forests. Southeast. Nat. 2011, 10, 591–608. [Google Scholar] [CrossRef]
- Harden, C.W.; Caterino, M.S.; Malabad, T.E. First Records of the Palearctic Species Trechus obtusus Erichson from the Appalachian Region of the Southeastern United States (Coleoptera: Carabidae: Trechinae: Trechini). Coleopt. Bull. 2022, 76, 61–69. [Google Scholar] [CrossRef]
- Kane, C.; Barr, T.C.; Stratton, E. Genetic patterns and population structure in Appalachian Trechus of the vandykei group. Brimleyana 1990, 16, 133–150. [Google Scholar]
- Faille, A.; Ribera, I.; Deharveng, L.; Bourdeau, C.; Garnery, L.; Queinnec, E.; Deuve, T. A molecular phylogeny shows the single origin of the Pyrenean subterranean Trechini ground beetles (Coleoptera: Carabidae). Mol. Phylogen. Evol. 2010, 54, 97–106. [Google Scholar] [CrossRef]
- Huang, S.; Tian, M.; Faille, A. A preliminary phylogeny of cave trechine beetles from China (Coleoptera: Carabidae: Trechini). In ARPHA Conference Abstracts; Pensoft Publishers: Sofia, Bulgaria, 2020; Volume 3. [Google Scholar] [CrossRef]
- Erwin, T.L.; House, G.N. A catalogue of the primary types of Carabidae (incl. Cicindelinae) in the collections of the United States National Museum of Natural History (USNM) (Coleoptera). Coleopt. Bull. 1978, 32, 231–256. [Google Scholar]
- Barr, T.C. The genus Trechus (Coleoptera: Carabidae: Trechini) in the southern Appalachians. Coleopt. Bull. 1962, 16, 65–92. [Google Scholar]
- Barr, T.C. Revision of Appalachian Trechus (Coleoptera: Carabidae). Brimleyana 1979, 2, 29–75. [Google Scholar]
- Barr, T.C. New Trechine Beetles (Coleoptera: Carabidae) from the Appalachian Region. Brimleyana 1985, 11, 119–132. [Google Scholar]
- Barr, T.C. Pattern and process in speciation of trechine beetles in eastern North America (Coleoptera: Carabidae: Trechinae). In Taxonomy, Phylogeny and Zoogeography of Beetles and Ants; Ball, G.E., Ed.; Dr W Junk Publishers: The Hague, The Netherlands, 1985. [Google Scholar]
- Donabauer, M. New Species and Subspecies of the Trechus (Microtrechus) nebulosus-group from the Southern Appalachians (Coleoptera: Carabidae: Trechinae). Z. Der Arbeitsgemeinschaft Der Osterr. Entomol. 2005, 57, 65–92. [Google Scholar]
- Donabauer, M. New species of the Trechus (Microtrechus) uncifer-group from the Southern Appalachians (Coleoptera: Carabidae). Z. Der Arbeitsgemeinschaft Der Osterr. Entomol. 2005, 57, 51–62. [Google Scholar]
- Donabauer, M. New species of the Trechus (Microtrechus) vandykei group from the southern Appalachians (Coleoptera: Carabidae: Trechinae). Z. Der Arbeitsgemeinschaft Der Osterr. Entomol. 2009, 61, 129–141. [Google Scholar]
- Donabauer, M. A taxonomic reorganization of European Trechus CLAIRVILLE, 1806 (Coleoptera: Carabidae: Trechinae). Z. Der Arbeitsgem. Osterr. Entomol. 2019, 71, 87–117. [Google Scholar]
- Möst, M.H.; Donabauer, M.; Arthofer, W.; Schlick-Steiner, B.C.; Steiner, F.M. Towards an evolutionary history of European-Alpine Trechus ground beetles: Species groups and wing reduction. Mol. Phylogen. Evol. 2020, 149, 106822. [Google Scholar] [CrossRef]
- Maddison, D.R.; Porch, N. A preliminary phylogeny and review of the genus Tasmanitachoides, with descriptions of two new species (Coleoptera, carabidae, bembidarenini). ZooKeys 2021, 1044, 153–196. [Google Scholar] [CrossRef]
- Maddison, D.R.; Kanda, K.; Boyd, O.F.; Faille, A.; Porch, N.; Erwin, T.L.; Rogi-Junent, S. Phylogeny of the beetle supertribe Trechitae (Coleoptera: Carabidae): Unexpected clades, isolated lineages, and morphological convergence. Mol. Phylogen. Evol. 2019, 132, 151–176. [Google Scholar] [CrossRef]
- Harden, C.W.; Davidson, R.L.; Malabad, T.E.; Caterino, M.S.; Maddison, D.R. Biology Phylogenetic systematics of the enigmatic genus Horologion Valentine, 1932 (Coleoptera, Carabidae, Trechinae, Horologionini), with description of a new species from Bath County, Virginia. Subterr. Biol. 2024, 49, 1–49. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Elbrecht, V.; Leese, F. Validation and development of freshwater invertebrate metabarcoding COI primers for environmental impact assessment. Front. Environ. Sci. 2017, 5, 1–11. [Google Scholar]
- Wild, A.L.; Maddison, D.R. Evaluating nuclear protein-coding genes for phylogenetic utility in beetles. Mol. Phylogen. Evol. 2008, 48, 877–891. [Google Scholar] [CrossRef]
- Shull, V.L.; Vogler, A.P.; Baker, M.D.; Maddison, D.R.; Hammond, P.M. Sequence alignment of 18S ribosomal RNA and the basal relationships of Adephagan beetles: Evidence for monophyly of aquatic families and the placement of Trachypachidae. Syst. Biol. 2011, 50, 945–969. [Google Scholar] [CrossRef]
- Maddison, D.R. Systematics of the North American beetle subgenus Pseudoperyphus (Coleoptera: Carabidae: Bembidion) based upon morphological, chromosomal, and molecular data. Ann. Carnegie Mus. 2008, 77, 147–193. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore sequencing technology, bioinformatics and applications. Nat. Biotechnol. 2021, 39, 1348–1365. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Molecular Analysis. 2001. Available online: https://www.mesquiteproject.org/home.html (accessed on 10 January 2024).
- Andújar, C.; Serrano, J.; Gámez-Zurita, J. Winding up the molecular clock in the genus Carabus (Coleoptera: Carabidae): Assessment of methodological decisions on rate and node age estimation. BMC Evol. Biol. 2012, 12, 40. [Google Scholar] [CrossRef]
- Faille, A.; Hofmann, S.; Merene, Y.; Hauth, D.; Opgenoorf, L.; Woldehawariat, Y.; Schmidt, J. Explosive radiation versus old relicts: The complex history of Ethiopian Trechina, with description of a new genus and a new subgenus. Dtsch. Entomol. Z. 2023, 70, 311–335. [Google Scholar] [CrossRef]
- Schmidt, J.; Belousov, I.; Michalik, P. X-ray microscopy reveals endophallic structures in a new species of the ground beetle genus Trechus Clairville, 1806 from Baltic amber (Coleoptera, Carabidae, Trechini). Zookeys 2016, 614, 113–127. [Google Scholar] [CrossRef]
- Faille, A.; Fresneda, J.; Bourdeau, C. Reconciling morphological and molecular data in a highly convergent group: The Pyrenean radiation of hypogean Trechini (Coleoptera: Carabidae). Integr. Syst. 2023, 6, 9–37. [Google Scholar] [CrossRef]
- Crespi, E.J.; Rissler, L.J.; Browne, R.A. Testing Pleistocene refugia theory: Phylogeographical analysis of Desmognathus wrighti, a high-elevation salamander in the southern Appalachians. Mol. Ecol. 2003, 12, 969–984. [Google Scholar] [CrossRef]
- Hermans, G.C.D.; Moya, O.; Oromi, P.; Juan, C. Evolution and diversification of the forest and hypogean ground-beetle genus Trechus in the Canary Islands. Mol. Phylogen. Evol. 2007, 42, 687–699. [Google Scholar]
- Leray, V.L.; Caravas, J.; Friedrich, M.; Zigler, K.S. Mitochondrial sequence data indicate “Vicariance by Erosion” as a mechanism of species diversification in North American Ptomaphagus (Coleoptera, Leiodidae, Cholevinae) cave beetles. Subterr. Biol. 2019, 29, 35–57. [Google Scholar] [CrossRef]
- Whitaker, M.; Procter, T.; Fontanella, F. Phylogeography and demographic expansion in the widely distributed horned passalus beetle, Odontotaenius disjunctus (Coleoptera: Passalidae). Mitochondrial DNA Part A 2021, 32, 85–97. [Google Scholar] [CrossRef]
- Anderson, W.A. A new species of Glyceria from the Great Smoky Mountains. Rhodora 1933, 35, 320–322. [Google Scholar]





| Gene | Forward Primer | Reverse Primer | Reference | Denaturing | Annealing | Cycle # | Sample Frequency |
|---|---|---|---|---|---|---|---|
| COI | LCO1490 | HCO2198 | [41] | 94 °C 0:30 | 50 °C 0:30 | 35 | Every Individual * |
| BF2 | BR2 | [42] | |||||
| Topo | TP643F | TP932R | [43] | 94 °C 0:30 | 57°/52°/45° 0:30 | 6/6/36 | Species Group |
| TP675F | TP932R | [43] | |||||
| CAD4 | CD806F | CD1098R2 | [43] | 94 °C 0:20 | 60 °C 0:20 | 39 | Species Group |
| CD821F | CD1098R2 | [43] | 94 °C 0:20 | 55 °C 0:20 | 37 | ||
| 18S | 18S5′ | 18Sb5.0 | [44] | 94 °C 0:30 | 50 °C 0:30 | 35 | Species Group |
| 28S | NLF184/21 | LS1041R | [45] | 94 °C 0:30 | 50 °C 0:30 | 35 | Species Group |
| Wingless | wg550F | wgAbRZ | [43] | 94 °C 0:20 | 52 °C 0:20 | 37 | Species |
| wg578F | wgAbR | [43] | 94 °C 0:20 | 54 °C 0:20 | 35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wooden, P.L.S.; Caterino, M.S. Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities. Diversity 2024, 16, 212. https://doi.org/10.3390/d16040212
Wooden PLS, Caterino MS. Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities. Diversity. 2024; 16(4):212. https://doi.org/10.3390/d16040212
Chicago/Turabian StyleWooden, Patricia L. S., and Michael S. Caterino. 2024. "Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities" Diversity 16, no. 4: 212. https://doi.org/10.3390/d16040212
APA StyleWooden, P. L. S., & Caterino, M. S. (2024). Trechus (Coleoptera: Carabidae) of Appalachia: A Phylogenetic Insight into the History of High Elevation Leaf Litter Communities. Diversity, 16(4), 212. https://doi.org/10.3390/d16040212