
Two New Species of Elaphoidella (Copepoda, Harpacticoida) from Subterranean Waters in Northeast Thailand, with a Record of a Gynandromorphic Specimen and an Up-to-Date Key to Elaphoidella Species from Southeast Asia †



Abstract
1. Introduction
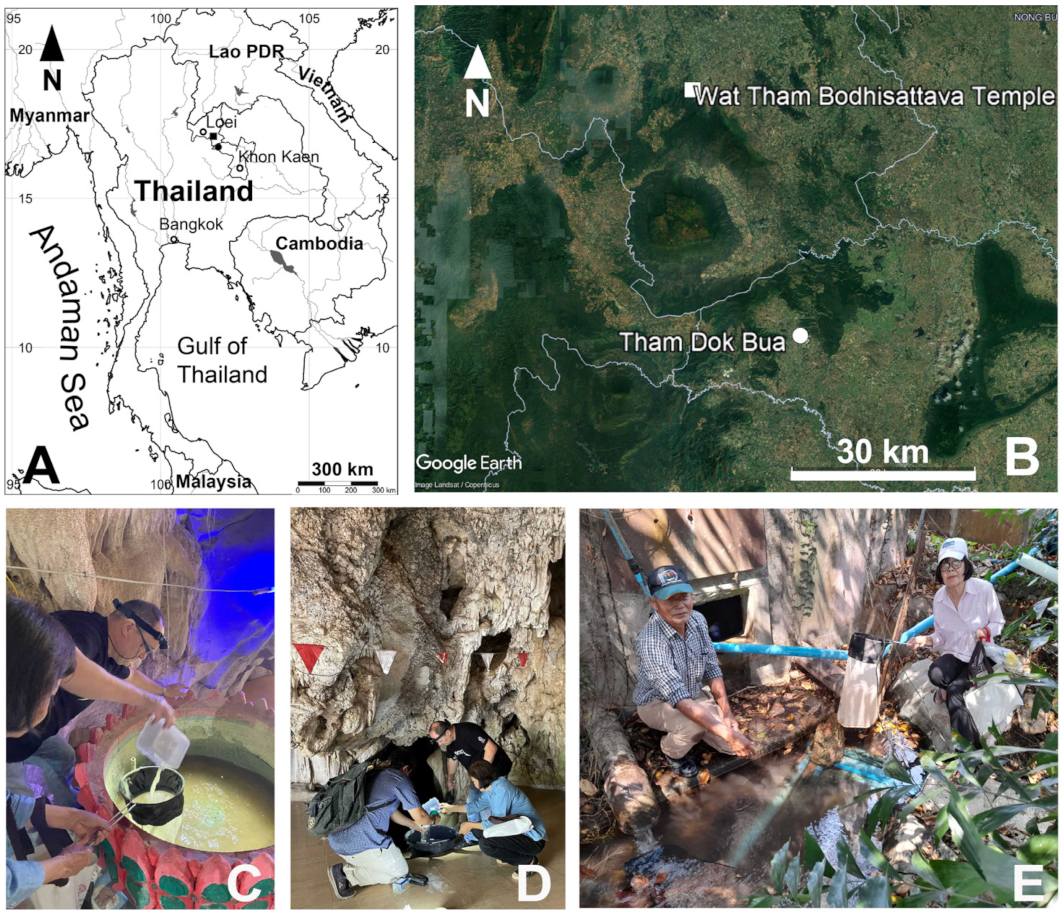
2. Materials and Methods
3. Results
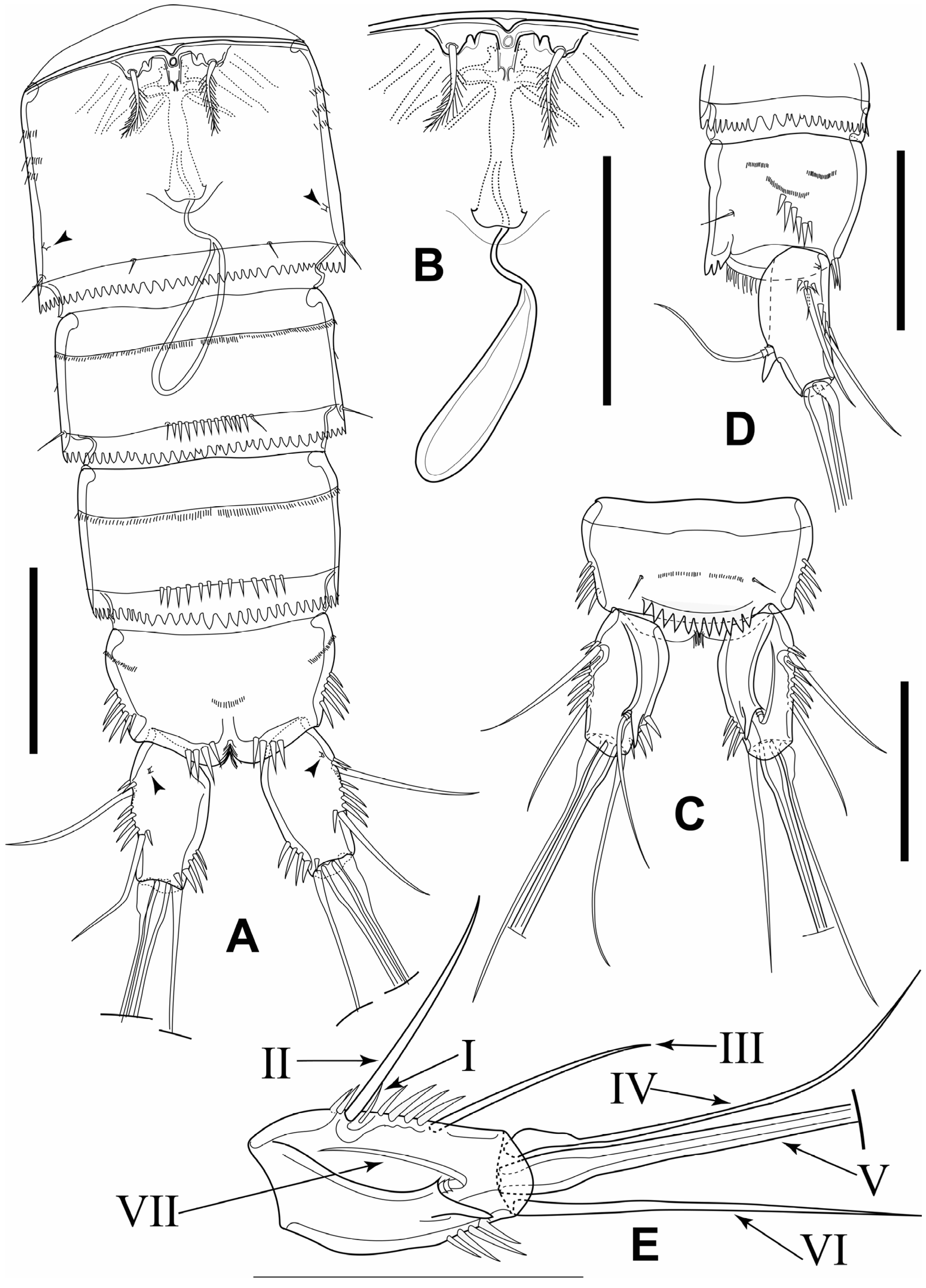
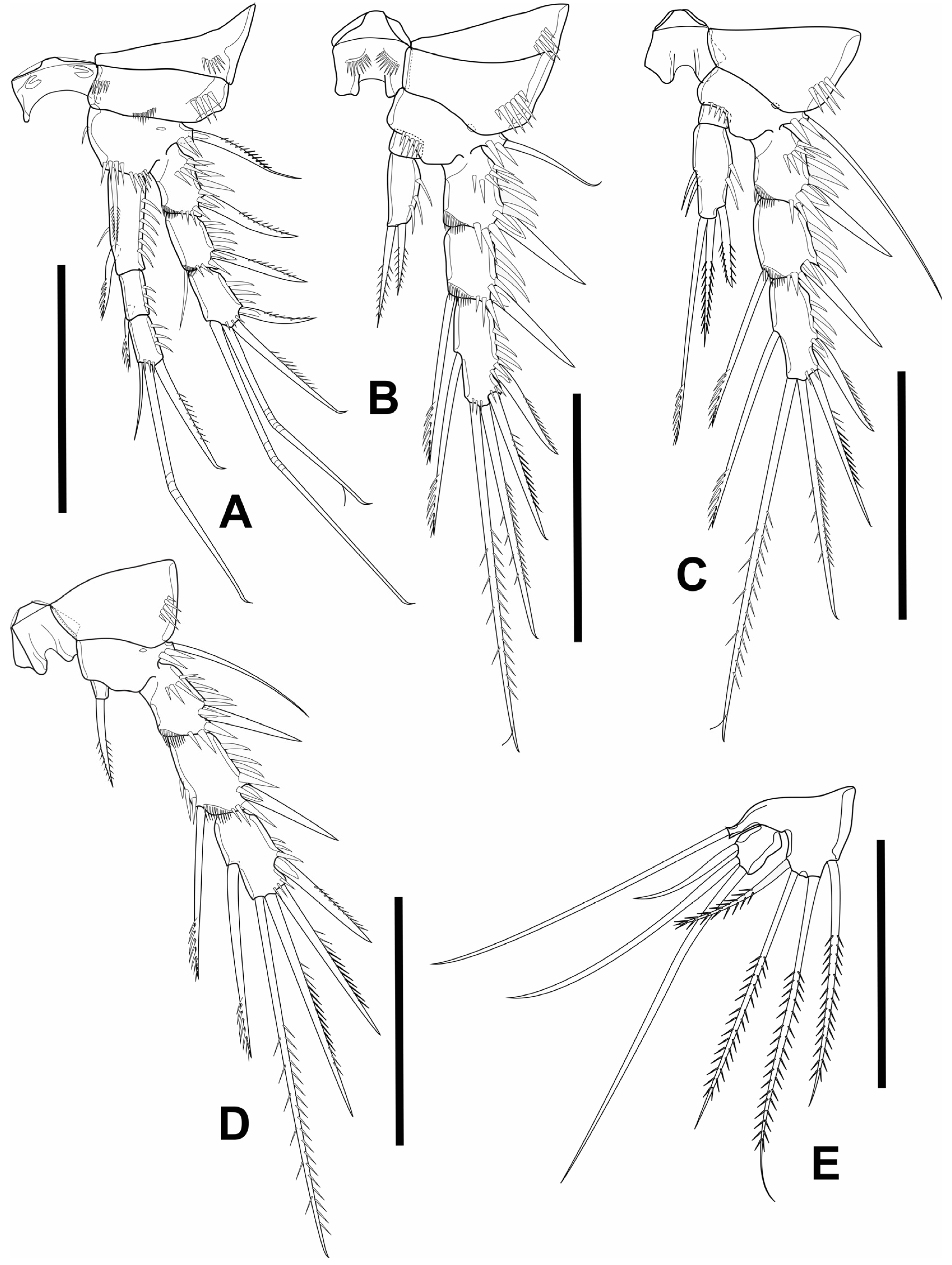
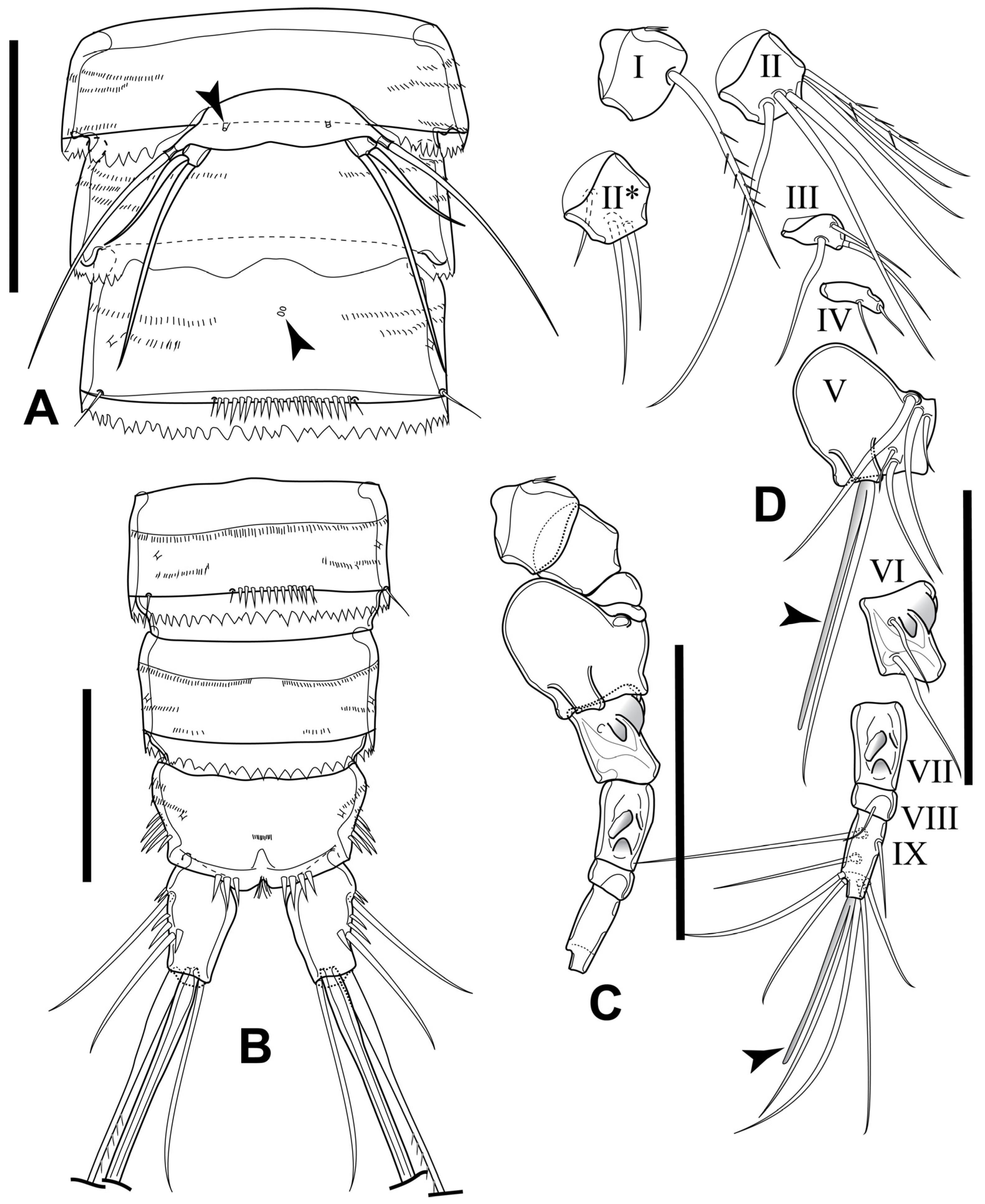
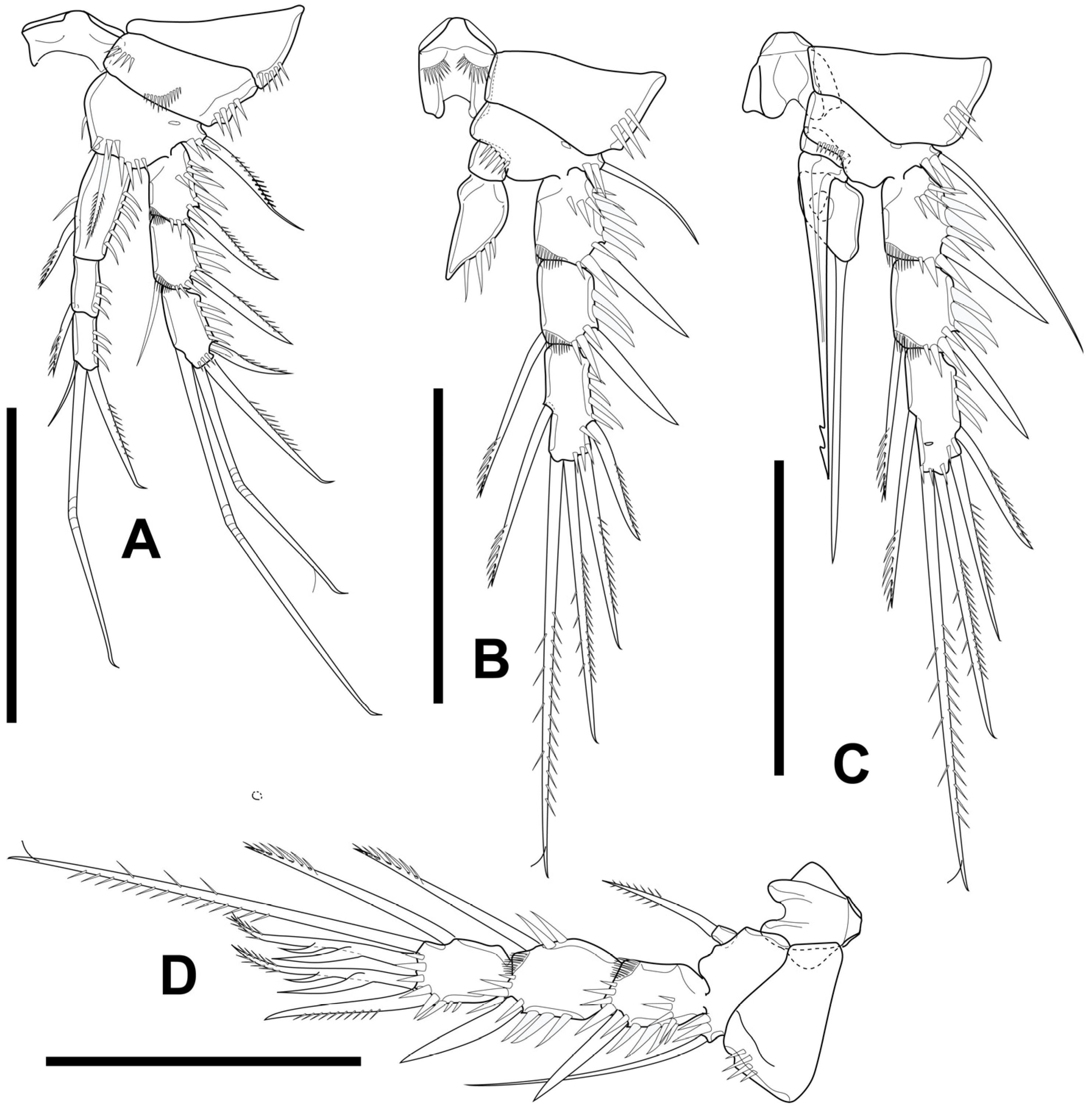
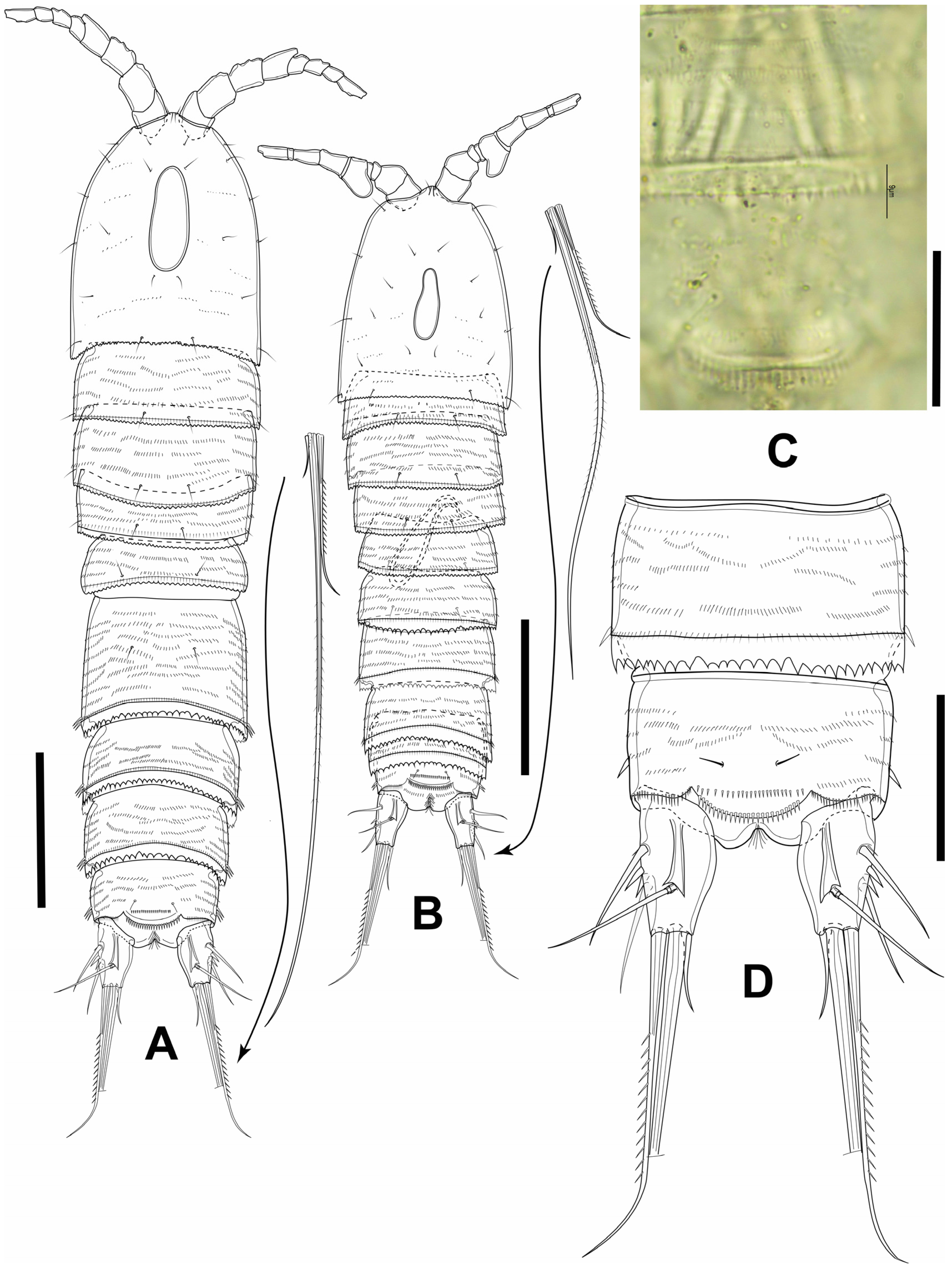
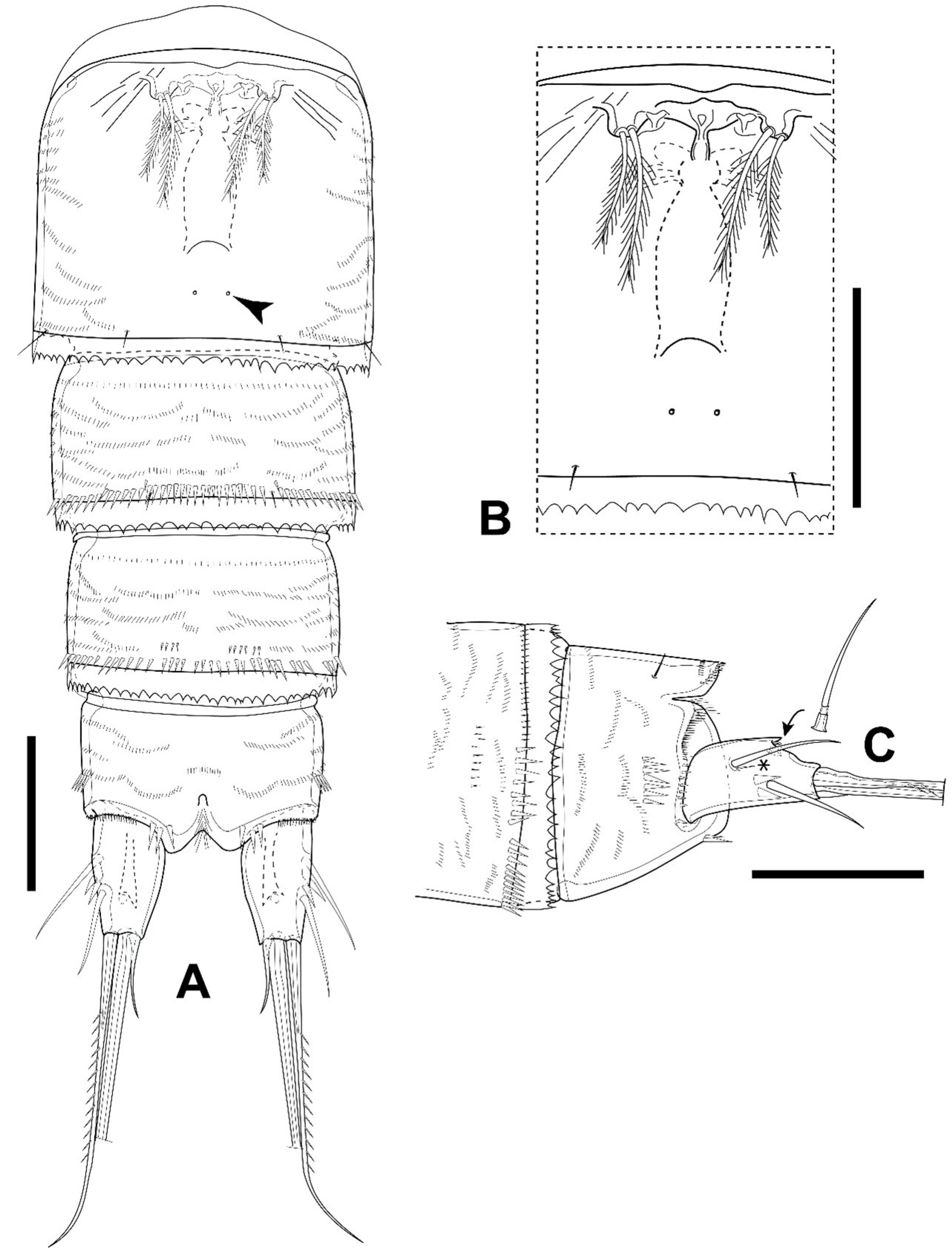
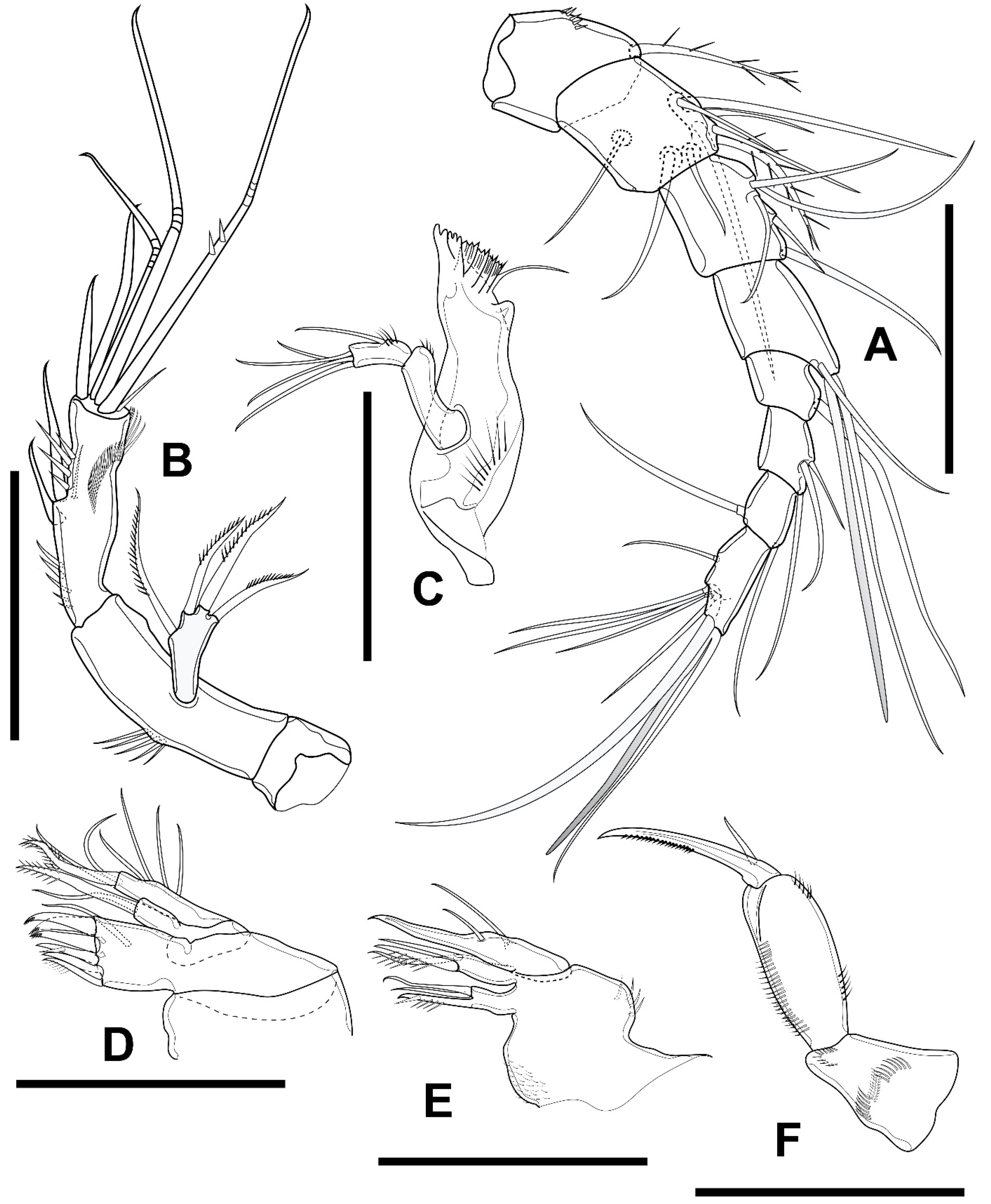
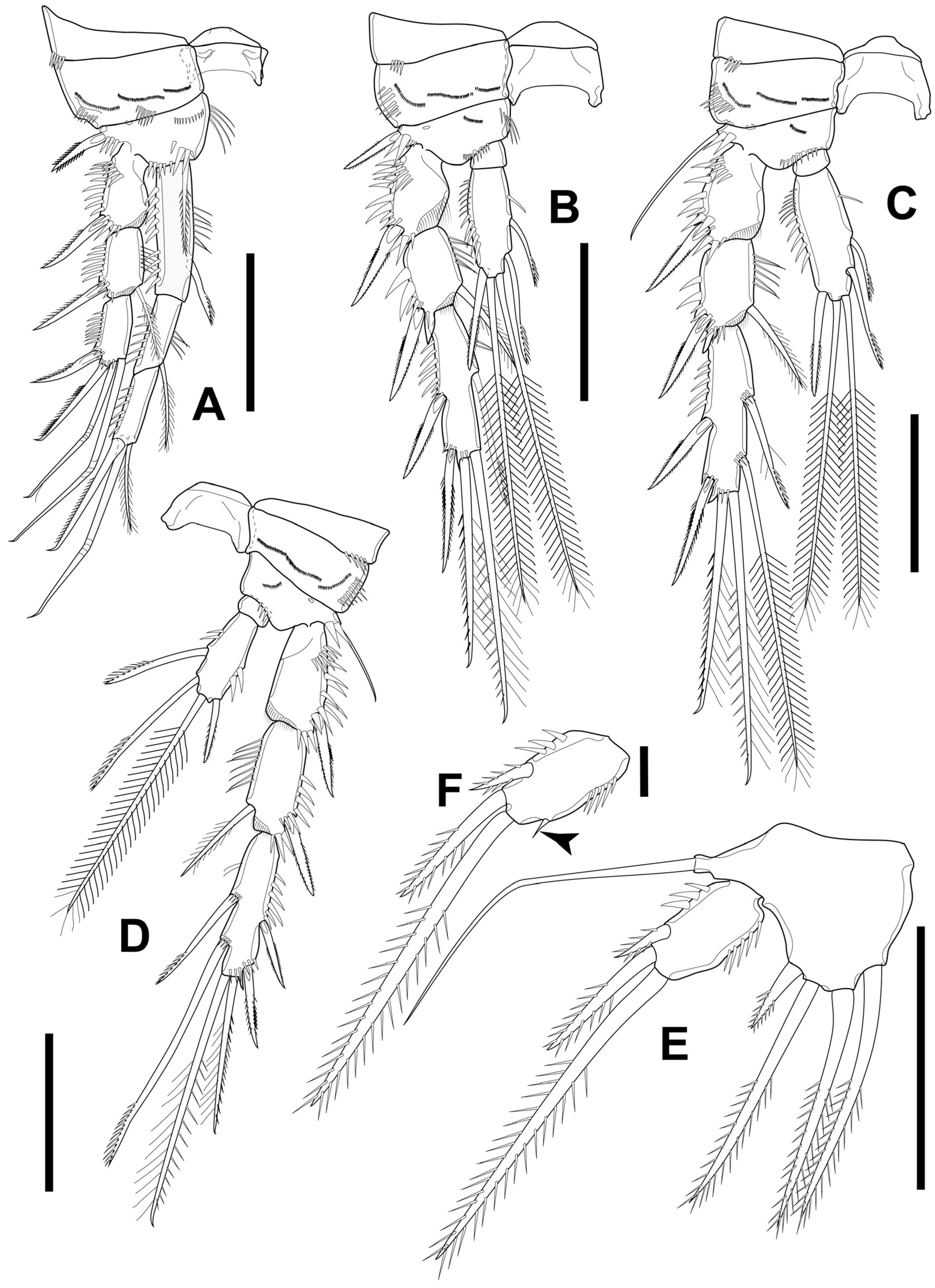
3.1. Elaphoidella phuphamanensis sp. nov.






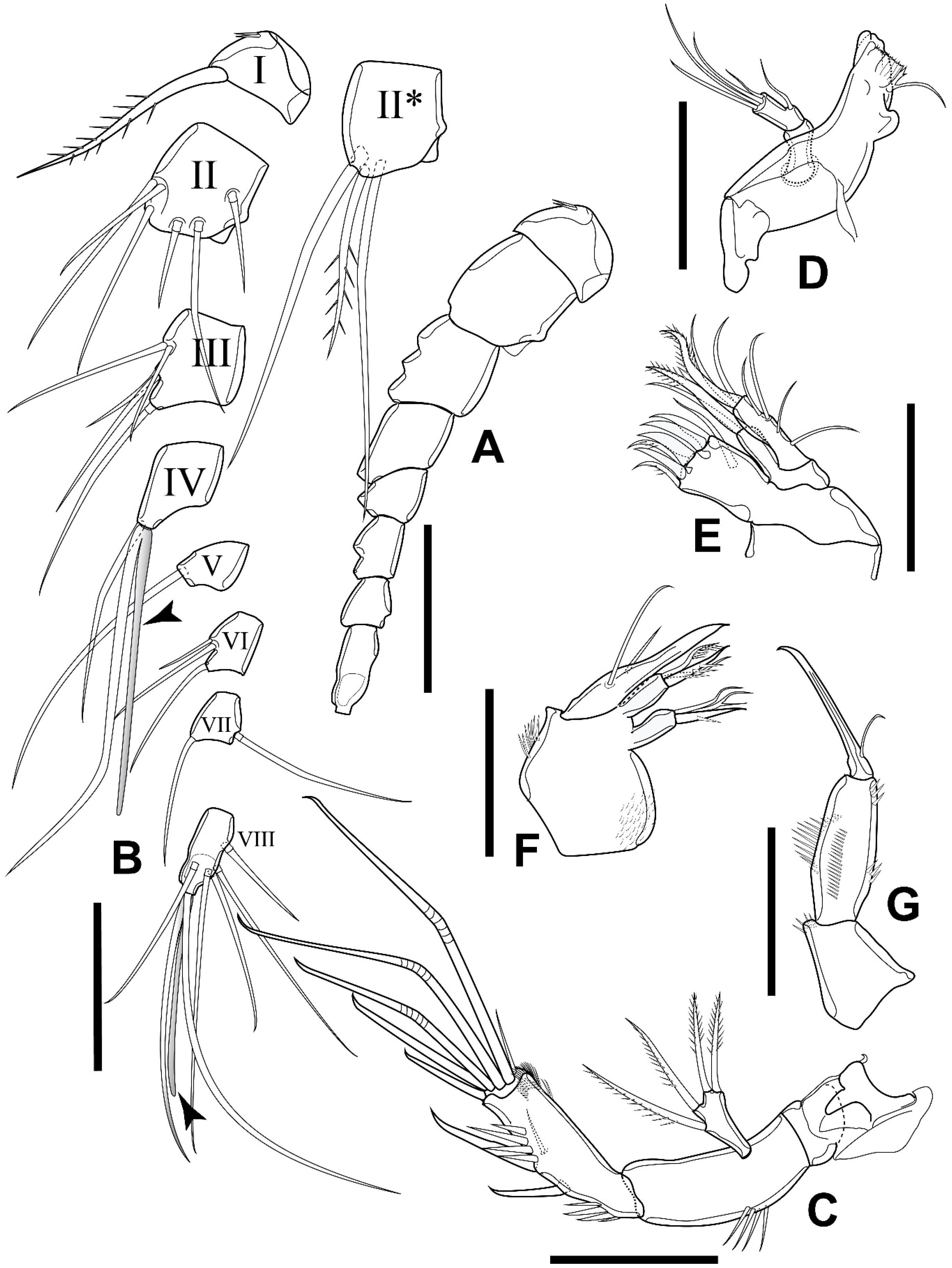
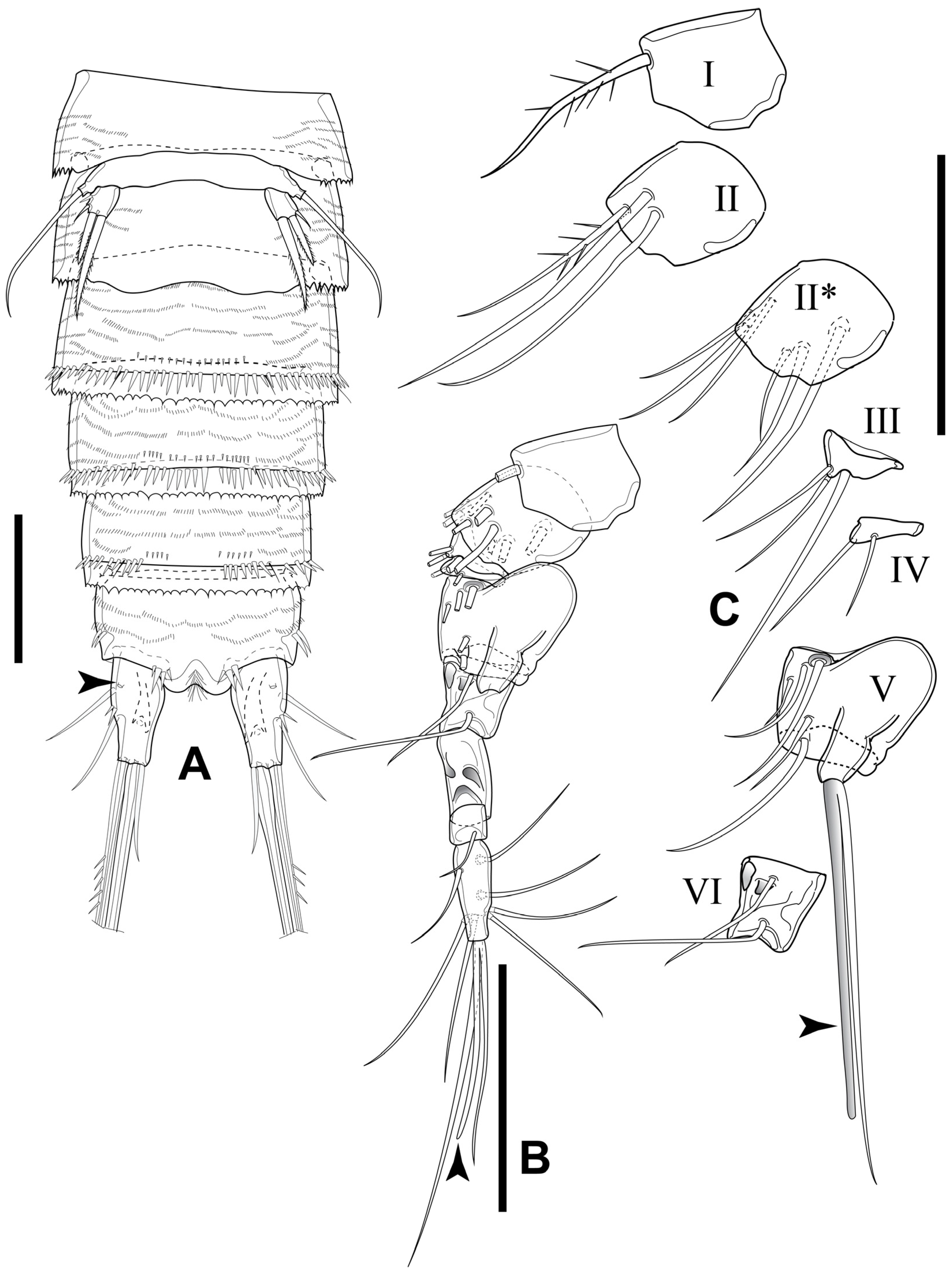
3.2. Elaphoidella propecabezasi sp. nov.






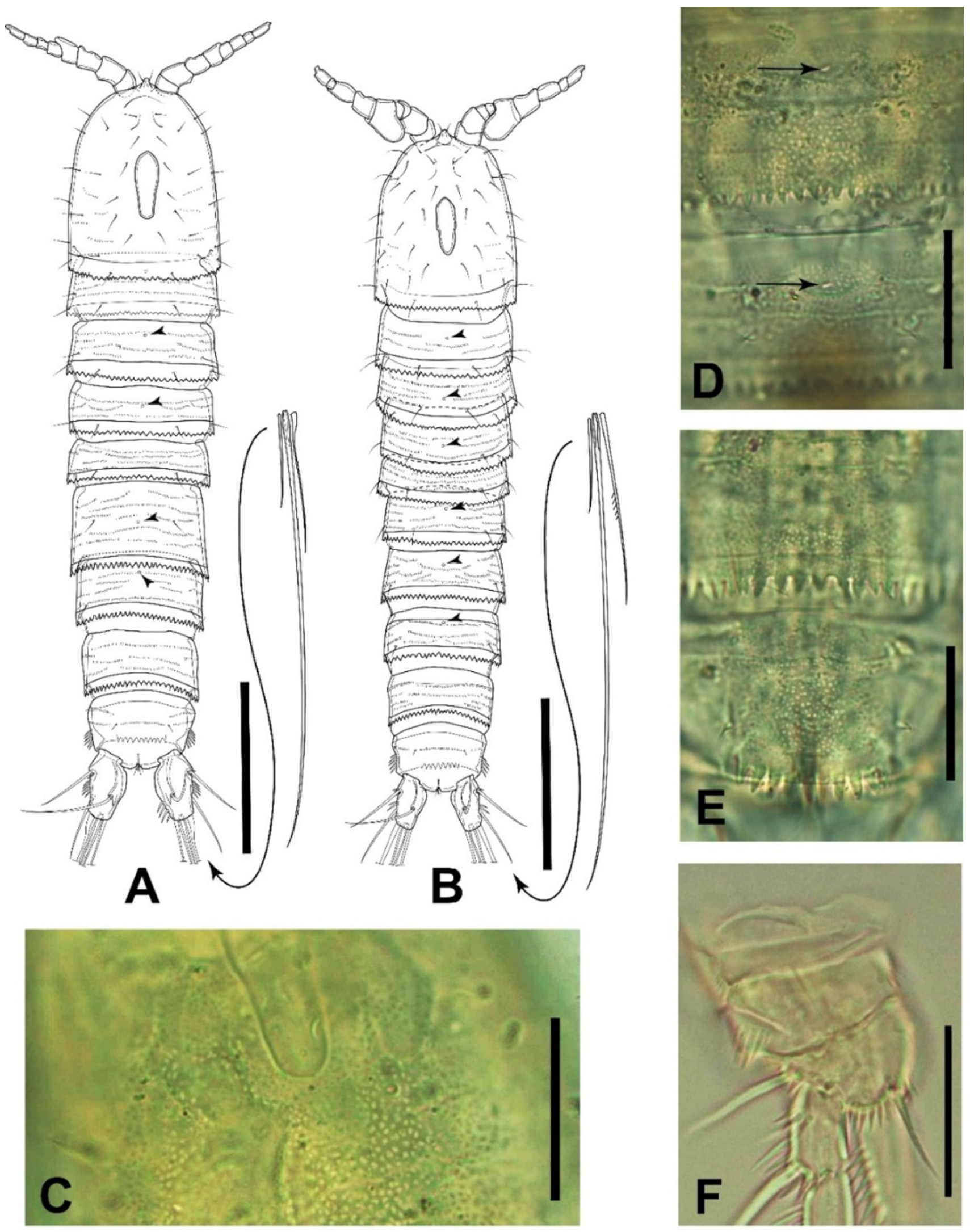
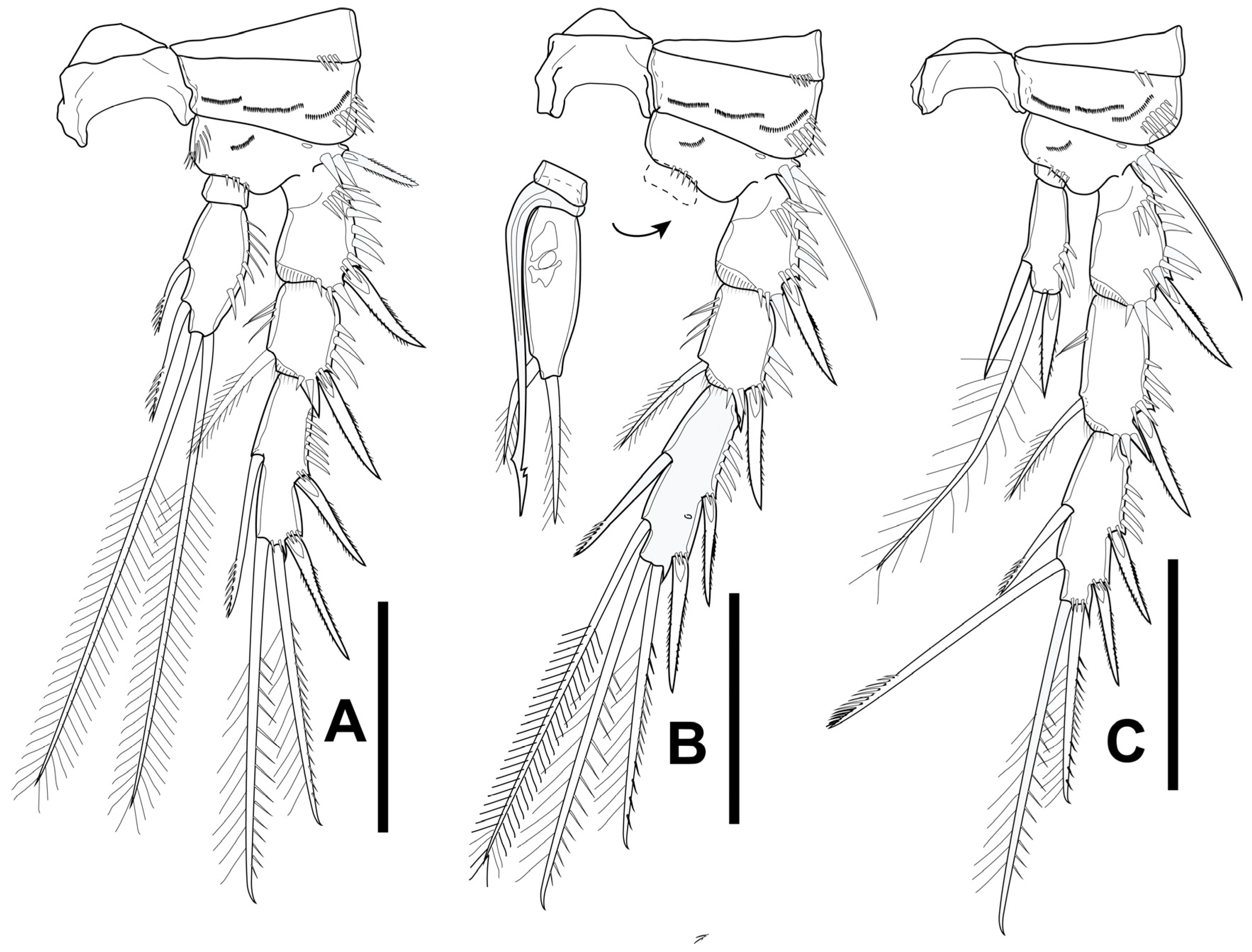
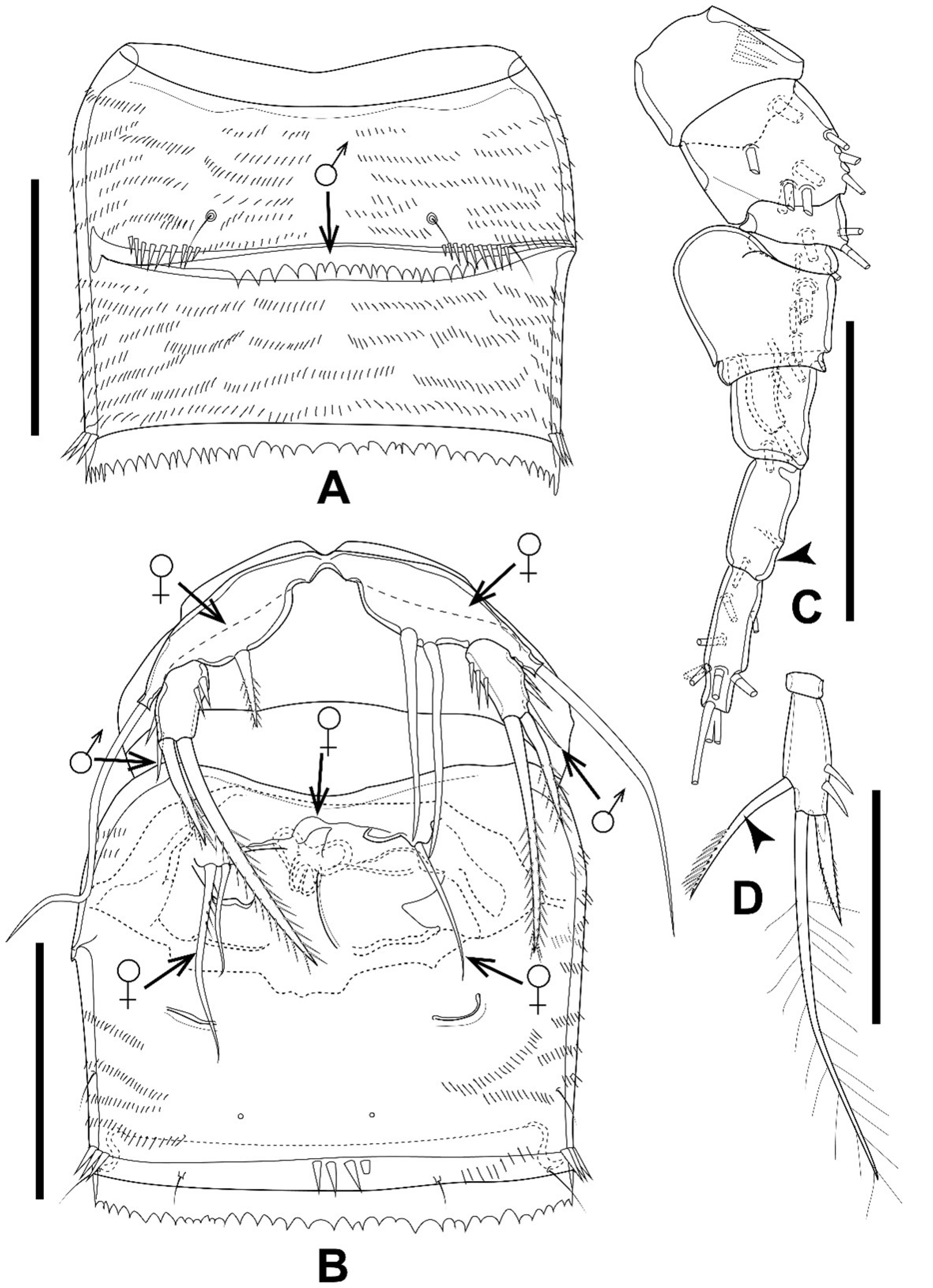
3.3. Gynandromorphic Specimen

4. Discussion
4.1. Presence of the Genus Elaphoidella in Thailand
4.2. Gynandromorphic Specimen
4.3. Amended Key to Elaphoidella Species from Southeast Asia
4.4. Key to the Females of Elaphoidella Chappuis, 1929, Recorded in Southeast Asia
| 1. - P1 Enp two-segmented …………………………….……..……………………………......….. 2 |
| - P1 Enp three-segmented ………………………….……………………………........................ 9 |
| 2. - P4 Enp completely reduced …………………………….……………...……………………… 3 |
| - P4 Enp two-segmented ……………………………….......…….……..…..………………….. 4 |
| 3. - P2–P3 Enp-2 each with two elements apically; inner seta reaching tip of Exp-3 at most …………………...........................................................….……………… E. thailandensis |
| - P2–P3 Enp-2 each with two and three elements apically, respectively: inner seta considerably surpassing tip of Exp-3 …............................……………….….. E. longiramus |
| 4. - P5 Enp lobe small, tip of Enp lobe reaching the middle of Exp at most; armature of P2–P4 Enp-1: 1.1.0 …………………………………………………………….......…. E. bidens bidens |
| - P5 Enp lobe, surpassing tip of Exp; armature of P2–P4 Enp-1: 0.0.0 ………………………………………….………………………………………………….. 5 |
| 5. - P5 Exp with three setae …………………….…………………..…………………….….….. 6 |
| - P5 Exp with four setae ………………………………….……………………………..…….. 7 |
| 6. - Caudal seta VI spiniform, curved inwards; seta IV (outermost lateral seta) on P5 Enp lobe minute, shorter than length of P5 Exp……………………..................................... E. cornuta |
| - Caudal seta VI normally developed; seta IV (outermost lateral seta) on P5 Enp lobe as long as or longer than the length of P5 Exp............................................................ E. javaensis |
| 7. - P1 Enp-2 with two elements apically: one outer spine and one geniculate, inner seta …...…….………………………………………………..……………………. E. stygobiota |
| - P1 Enp-2 with three elements apically …………………………………………………….. 8 |
| 8. - All seta on caudal ramus normally developed; anal operculum with several short and small spinules on its free margin ………………………….……………………… E. elegans |
| - Caudal seta V and VI modified, swollen or bulbous at base; anal operculum with strong spinules on its free margin ………………………….…....……...………… E. sanoamuangae |
| 9. - P4 with one-segmented Enp ………..………………………… E. phuphamanensis sp. nov. |
| - P4 with two-segmented Enp …………………………………………………….…..…….. 10 |
| 10. - Armature of P2–P4 Enp-1: 1.1.1 or 1.0.0 ………………………………………..….……… 11 |
| - Armature of P2–P4 Enp-1: 1.1.0 or 0.1.0 ……………………………………..…..……….. 21 |
| - Armature of P2–P4 Enp-1: 0.0.0 …………………………….………………….……..…… 30 |
| 11. - Armature of P2–P4 Enp-1: 1.0.0 ………………………………………………….. E. ligorae |
| - Armature of P2–P4 Enp-1: 1.1.1 …………………………………………………………… 12 |
| 12. - Free margin of anal operculum with 10–14 strong spinules…………..………………… 13 |
| - Free margin of anal operculum with fine hairs or fine spinules or comb-like, (more than 18 hairs, spinules or indentations) ......................…………………………………………… 14 |
| 13. - Caudal ramus subquadrate, with acute extension dorsally ………….. E. bidens decorata |
| - Caudal ramus conical, without acute extension dorsally .……………..…......... E. fatimae |
| 14. - P5 Enp lobe with three elements ……………………....................................... E. trisaetosa |
| - P5 Enp lobe with four elements …………………………………………………..………. 15 |
| 15. - P5 Exp as long as or more than 3.0 times as long as wide ………..……..…..…………. 16 |
| - P5 Exp less than 2.5 times as long as wide ……………….…….…………….………….. 17 |
| 16. - P5 Exp less than 4.0 times as long as wide …….……………………………. E. longipedis |
| - P5 Exp more than 4.6 times as long as wide ……………………………… E. superpedalis |
| 17. - P5 Exp with three or four setae…………………………………….…….……….. E. affinis |
| - P5 Exp with six setae ………………….............................................................. E. bryophila |
| - P5 Exp with five setae …………………………………………………….….…………… 18 |
| 18. - Caudal seta IV and VI modified; caudal seta VI short, wavy; caudal seta IV short, bulbous, pear-shaped …………………………………………………………………...…. E. cuspidata |
| - Caudal seta IV normally developed, setiform ……………..…………….…………….. 19 |
| 19. - All setae on P5 Exp normally developed, seta IV on P5 Exp as long as or longer than supporting segment ………………………………………..………………………………… 20 |
| - Two outer seta on P5 Exp short, slim, bare, seta IV much shorter than supporting segment ………………………………………………………………………………… E. isana |
| 20. - Caudal seta III inserted near seta IV, below the insertion of caudal seta IV; spinule on lateral surface of urosomite 4 relatively large and strong……..…………...……. E. similis |
| - Caudal seta III inserted on lateral surface, below the insertion of caudal seta II; spinule on lateral surface of urosomite 4 slim .....……..………………………………. E. intermedia |
| 21. - Armature of P2–P4 Enp-1: 0.1.0 ……………………………………………….………..… 22 |
| - Armature of P2–P4 Enp-1: 1.1.0 ……………….……………………………..……………. 23 |
| 22. - Inner marginal and apical setae on P4 Enp-2 less than three times as long as supporting segment; caudal rami without spinules at base of caudal seta V and VI ventrally………… E. malayica |
| - Inner marginal and apical setae on P4 Enp-2 relatively long, more than three times as long as the segment bearing them; caudal rami with or without spinules at base of caudal seta V and VI ventrally…...…..………………………………………………………………... E. labani |
| 23. - P5 Enp lobe relatively short, insertion of seta II on P5 Enp lobe reaching the middle of Exp at most ……………………………………………………………………..………...…………. 24 |
| - P5 Enp lobe relatively long, insertion of seta II on P5 Enp lobe reaching or surpassing tip of Exp ……………………………………………………..…………………………..………… 26 |
| 24. - Anal operculum short, reaching distal margin of anal somite, free margin with strong spinules; dorsal keel on caudal ramus with acute extension distally …... E. bidens coronata |
| - Anal operculum surpassing distal margin of anal somite, semicircular or semi-oval in shape, free margin comb-like; dorsal keel on caudal ramus without acute extension distally …………….……………………………………………………………..……………… 25 |
| 25. - Free margin of anal operculum with about 40 spinules…….…………….. E. grandidieri |
| - Free margin of anal operculum with about 20 oblong spinules ………… E. namnaoensis |
| 26. - P4 Enp-2 with three (exceptionally two) setae and spines …………..…..…...………… 27 |
| - P4 Enp-2 with four setae and spines ….………...………………………..…..…………… 28 |
| 27. - P3 Enp-2 with four elements; inner lateral seta on P4 Enp-2 relatively short, less than three times as long as supporting segment; caudal rami without spinules at base of caudal seta V and VI ventrally……..……….…………………………………………………. E. sewellis. str. |
| - P3 Enp-2 with five elements; inner lateral seta on P4 Enp-2 relatively long, more than three times as long as supporting segment; caudal rami with or without spinule at base of caudal seta V and VI ventrally .….…… ………………………………………………………. E. labani |
| 28. - P3 Enp-2 with three elements ……………………………………………. E. bromeliaecola |
| - P3 Enp-2 with five elements ……….……………………….…...……… E. thienemanni (29) |
| 29. - - Two apical elements on P4 Enp-2 spiniform setae ………...................... E. t. thienemanni |
| - Two apical elements apically on P4 Enp-2 comprisingg one spiniform and one longer seta……………………………………………………………..…….…………….. E. t. serrulata |
| 30. - P5 Exp with two–four elements; P3 Enp-2 with four–five elements ………………….. 31 |
| - P5 Exp with five elements; P3 Enp-2 with six elements …………….… ….. E. jaesornensis |
| 31. - P3 Enp-2 with four setae and spines ……………………………………………………… 32 |
| - P3 Enp-2 with five setae and spines ……………………………………….……………… 33 |
| 32. - Armature formula of P2–P4 Enp-2: 5.4.3………………………………………. E. brancelji |
| - Armature formula of P2–P4 Enp-2: 4.4.3………...…………………………… E. vietnamica |
| 33. - Armature formula of P2–P4 Enp-2: 5.5.3; P5 Exp with two setae …………. E. margaritae |
| - Armature formula of P2–P4 Enp-2: 5.5.4; P5 Exp with three or four setae……………. 34 |
| 34. - P5 Exp with three elements ……………………………………. E. propecabezasi sp. nov. |
| - P5 Exp with four elements ………………..…………………….……………E. paraaffinis |
4.5. Key to the Males of Elaphoidella Chappuis, 1929, Recorded in Southeast Asia
| 1. - P1 Enp two-segmented ……………….……………………………………..………………. 2 |
| - P1 Enp three-segmented ……………………..……………………………………………… 6 |
| 2. - P4 Enp completely reduced; outer apical element of P4 Exp-3 transformed, antler-like spine ……………………………………………..……………………...……..….…………….. 3 |
| - P4 Enp two-segmented ………………………………….………….…………….…………. 4 |
| 3. - P2 Enp-2 with two elements: outer short and spiniform, inner one seta; segment 5 of A1 with spinous process in median anterior margin and accompanied by one seta…………………...................................................................……………... E. thailandensis |
| - P2 Enp-2 with one, long seta; A1 normally developed as characteristic of the genus ………………………………...…………………….……………………. E. longiramus |
| 4. - P4 Exp-3 with distal outer and outer apical antler-like, transformed spines; anal operculum with about nine strong spinules on free margin ………….. E. sanoamuangae |
| - P4 Exp-3 without transformed spines …………………….…………….…………………. 5 |
| 5. - P5 Exp with two elements; P3 Enp-3 with two elements; P2 Enp-2 with two or three elements ……….………………………….………………………………...……... E. javaensis |
| - P5 Exp with three elements; P3 Enp-2 with one element; P2 Enp-2 with one apical element ……………………….………………….………………………………. E. stygobiota |
| 6. - P4 Exp-3 normally developed, more than 1.5 times as long as wide, without transformed elements; P3 Enp-3 with two elements………………………………………….................... 7 |
| - P4 Exp-3 normally developed, more than 1.5 times as long as wide; proximal outer spine transformed and shorter than distal outer one…...…..…....….…………………………… 11 |
| - P4 Exp-3 shortened; one or two elements on outer half of P4 Exp-3 transformed into antler-like spine …...…..............................................................................................………… 13 |
| 7. - P5 Exp with two elements ………………….…….………...…..……..……… E. margaritae |
| - P5 Exp with three elements, two inner ones spiniform, outermost one short, slender seta …………………………………………...…………………….. E. propecabezasi sp. nov. |
| - P5 Exp with four elements ……………………………….…………….………….……….. 8 |
| 8. - P4 Enp-2 with two elements ………………………………..…………….….……. E. similis |
| - P4 Enp-2 with three elements ……………………………….…………….……………….. 9 |
| 9. - P2 Enp-2 with five elements ……………………………………………......…….. E. ligorae |
| - P2 Enp-2 with four elements ………………………………………..…………………….. 10 |
| 10. - Inner apical element on P3 Enp-3 modified, obtuse, with long hairs……… E. cuspidata |
| - Inner apical element on P3 Enp-3 normally developed, setiform ……….. E. paraaffinis |
| 11. - P5 Exp with three setae; anal operculum reaching mid of caudal ramus, free margin with about 40 spinules……………….……………………………………………….. E. grandidieri |
| - P5 Exp with four setae ……………………………………..………………..…….………. 12 |
| 12. - Caudal ramus without dorsal keel, about 1.5 times as long as wide; P5 Exp shorter than wide, with three spiniform elements on inner margin; length of seta II (inner to outer) less than two times of innermost one …………………….……..………………….. E. bryophila |
| - Caudal ramus with dorsal keel, about twice as long as wide; P5 Exp as long as wide, three inner elements setiform, length of seta II more than two times the innermost one …………………………….……………………….……..…………………... E. intermedia |
| 13. - Only outer apical element of P4 Exp-3 transformed …...……………………………….. 14 |
| - Both distal outer and outer apical elements of P4 Exp-3 transformed…….................... 16 |
| 14. - P2 and P4 Enp-2 with four and three elements, respectively…………… E. sewellis. str. |
| - P2 and P4 Enp-2 each with three elements………………………………...…………….. 15 |
| 15. - Caudal ramus subquadrate; free margin of anal operculum with about six strong spinules; apical seta on P2 Enp-2 short and slim, as long as or slightly longer than supporting segment; distal inner seta on P2 Enp-2 reaching the tip of the apical seta.………................................................................................................................. E. malayica |
| - Caudal ramus slightly longer than wide, conical; free margin of anal operculum with 18–20 short spinules ………………………..……………..……………………….. E. vietnamica |
| - Caudal ramus about twice as long as wide, crescent, outer margin strongly concaved; free margin of anal operculum with more than 25 spinules; apical seta on P2 Enp-2 more than twice as long as distal inner seta ………………………………………………… E. brancelji |
| 16. - P4 Enp-2 one-segmented; P4 Exp-3 subquadrate; P2 Enp-2 without armature ………………………………………..…… E. phuphamanensis sp. nov. |
| - P4 Enp-2 two-segmented ……………..…………………………..……………………….. 17 |
| 17. - P2 and P4 Enp-1 each without inner seta ………………..………………………. E. labani |
| - P2 and P4 Enp-1 with and without inner seta, respectively …………………………… 18 |
| 18. - P3 Enp-3 with two short setae apically, subequal in length; P5 Exp wider than long; length of apical seta on P2 Enp-2 less than twice length of distal inner seta; outer spine on P3 Exp-2 just reaching insertion of proximal spine of P3 Exp-3 on outer margin ………………………….………………………..…………………… E. bromeliaecola |
| - P3 Enp-3 with thick blunt spine and much longer seta; P5 Exp as long as wide; length of apical seta on P2 Enp-2 more than twice length of distal inner seta; outer spine on P3 Exp-2 reaching tip of P3 Exp-3 …………..………………….…………………..… E. thienemanni |
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walter, T.C.; Boxshall, G. World of Copepods Database. Available online: https://marinespecies.org/aphia.php?p=taxdetails&id=1080 (accessed on 28 February 2024).
- Lang, K. Monographie der Harpacticiden II; Nordiska Bokhandeln: Stockholm, Sweden, 1948. [Google Scholar]
- Gaviria, S. Zwei Canthocamptidae (Copepoda, Harpacticoida) aus kolumbianischen Andengewässern. Ann. Naturhist. Mus. Wien B Bot. Zool. 1993, 94–95, 361–375. [Google Scholar]
- Mori, N.; Brancelj, A. Distribution and habitat preferences of species within the genus Elaphoidella Chappuis, 1929 (Crustacea: Copepoda: Harpacticoida) in Slovenia. Zool. Anz. 2008, 247, 85–94. [Google Scholar] [CrossRef]
- Rundle, S.D.; Bilton, D.T.; Shiozawa, D.K. Global and regional patterns in lotic meiofauna. Freshw. Biol. 2000, 44, 123–134. [Google Scholar] [CrossRef]
- Chappuis, P.A. Neue Harpacticiden aus Java. Treubia 1928, 10, 271–283. [Google Scholar]
- Dumont, H.J.; Maas, S. Five new species of leaf litter harpacticoids (Crustacea, Copepoda) from Nepal. Zool. Scr. 1988, 17, 55–68. [Google Scholar] [CrossRef]
- Reid, J. The harpacticoid and cyclopoid copepod fauna in the Cerrado Region of central Brazil. 1. Species composition, habitats and zoogeography. Acta Limnol. Bras. 1993, 6, 56–68. [Google Scholar]
- Galassi, D.M.P. Groundwater copepods: Diversity patterns over ecological and evolutionary scales. Hydrobiologia 2001, 453/454, 227–253. [Google Scholar] [CrossRef]
- Brancelj, A. Fauna of an unsaturated karstic zone in central Slovenia: Two new species of Harpacticoida (Crustacea: Copepoda), Elaphoidella millenni n. sp. and E. tarmani n. sp., their ecology and morphological adaptations. Hydrobiologia 2009, 621, 85–104. [Google Scholar] [CrossRef]
- Watiroyram, S.; Sanoamuang, L.; Brancelj, A. Two new species of Elaphoidella (Copepoda, Harpacticoida) from caves in southern Thailand and a key to the species of Southeast Asia. Zootaxa 2017, 4282, 501–525. [Google Scholar] [CrossRef]
- Daday, E. Untersuchungen über die Copepodenfauna von Hinerindien, Sumatra und Java, nebst einem Beitrag zur Copepodenkenntnis der Hawaii-Inseln. (Reise von Dr. Walter Volz). Zool. Jahrb. Abt. Syst. Georgr. Biol. Tiere. 1906, 24, 175–206. [Google Scholar]
- Pesce, G.L.; Apostolov, A.M. Elaphoidella margaritae sp. n. a new phreatobitic harpacticoid from subterranean waters of Thailand (Crustacea, Copepoda, Canthocamptidae. Acta Zool. Bulg. 1985, 28, 70–75. [Google Scholar]
- Deharveng, L.; Bedos, A. The cave fauna of Southeast Asia: Origin, evolution and ecology. In Ecosystems of the World; Wilkens, H., Culver, D.C., Humphreys, W.F., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2000; Volume 30, pp. 603–632. [Google Scholar]
- Deharveng, L.; Ellis, M.; Bedos, A.; Jantarit, S. Tham Chiang Dao: A hotspot of subterranean biodiversity in northern Thailand. Diversity 2023, 15, 1076. [Google Scholar] [CrossRef]
- Brancelj, A.; Watiroyram, S.; Sanoamuang, L. The first record of cave-dwelling Copepoda from Thailand and description of a new species: Elaphoidella namnaoensis, sp. nov. (Copepoda, Harpacticoida). Crustaceana 2010, 83, 779–793. [Google Scholar] [CrossRef]
- Watiroyram, S.; Wongduan, J.; Sanoamuang, L. Elaphoidella longiramus sp. nov. (Copepoda: Harpacticoida, Canthocamptidae), a new species of cave-dwelling copepod from central Thailand. Zootaxa 2022, 5138, 152–166. [Google Scholar] [CrossRef]
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991. [Google Scholar]
- Petkovski, T.K. Subterrane Süßwasser-Harpacticoida von Kuba. In Résultats des Expéditions Biospéologiques Cubano-Roumaines à Cuba; Orghidan, T.N., Ed.; Editura Academiei Republicii Socialiste Romania: Bucharest, Romania, 1973; Volume 1, pp. 125–141. [Google Scholar]
- Chappuis, P.A. Copepoda Harpaticoida der Deutschen Limnologischen Sunda-Expedition. In Tropische Binnengewässer Band 8; Ruttner, F., Ed.; Schweizerbart Science Publishers: Stuttgart, Germany, 1931; pp. 512–584. [Google Scholar]
- Löffler, H. Die Harpacticidenfauna des Mt. Kinabalu (Borneo) mit besonderer Berücksichtigung der Gattung Maraenobiotus nebst Angaben zur Harpacticidenfauna des Gebietes Nuwara (Hochplateau Ceylon). In Hochgebirgsforschung Heft 3: Beiträge zur Kenntnis Einiger Kleinorganismen Tropischer Hochgebirgsseen; Helmich, W., Ed.; Universitatsverlag Wagner: Innsbruck, Austria, 1973; pp. 5–28. [Google Scholar]
- Watiroyram, S.; Brancelj, A. A new species of the genus Elaphoidella Chappuis (Copepoda, Harpacticoida) from a cave in the south of Thailand. Crustaceana 2016, 89, 459–476. [Google Scholar] [CrossRef]
- Wells, J.B.J. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef]
- Apostolov, A.M. Étude sur quelques copépodes harpacticoïdes du genre Elaphoidella Chappuis, 1929 de Bulgarie avec une révision du genre. Acta Mus. Maced. Sci. Nat. 1985, 17, 133–163. [Google Scholar]
- Petkovski, T.K. Weitere neue Elaphoidella-Arten (Copepoda, Harpacticoida) aus den subterranen Binnengewässern von Kuba. Acta Mus. Maced. Sci. Nat. 1982, 16, 139–174. [Google Scholar]
- Young, S.S. Four new species of freshwater harpacticoid copepods (Canthocamptidae: Copepoda) from mountain lakes of Taiwan. Taiwan J. Biodivers. 2010, 12, 327–340. [Google Scholar]
- Gaviria, S.; Defaye, D. Description of Elaphoidella paramuna n. sp. (Canthocamptidae), a new harpacticoid copepod from Colombia. Crustaceana 2015, 88, 1003–1029. [Google Scholar] [CrossRef]
- Fefilova, E.B.; Alekseev, V.R. A new species and new records of harpacticoids (Crustacea: Copepoda: Harpacticoida) from North-Eastern Borneo. Zoosyst. Ross. 2018, 27, 205–217. [Google Scholar] [CrossRef]
- Cottarelli, V.; Bruno, M.C.; Berera, R. First record of Parastenocarididae from Thailand and description of a new genus (Copepoda: Harpacticoida). J. Crustac. Biol. 2010, 30, 478–494. [Google Scholar] [CrossRef]
- Boonyanusith, C.; Sanoamuang, L.; Brancelj, A. A new genus and two new species of cave-dwelling cyclopoids (Crustacea, Copepoda) from the epikarst zone of Thailand and up-to-date keys to genera and subgenera of the Bryocyclops and Microcyclops groups. Eur. J. Taxon. 2018, 431, 1–30. [Google Scholar] [CrossRef]
- Brancelj, A.; Boonyanusith, C.; Sanoamuang, L. A new cyclopoid genus (Copepoda, Crustacea) from a deep aquifer in northeastern Thailand with comments on peculiar sampling sites and local fauna. Raffles Bull. Zool. 2024, 72, 71–83. Available online: https://lkcnhm.nus.edu.sg/wp-content/uploads/sites/10/2024/02/RBZ-2024-0005.pdf (accessed on 28 February 2024).
- Pesce, G.L. Pesce’s Web Portal. Available online: http://www.luciopesce.net/copepods/arpa/elaph.htm (accessed on 28 December 2023).
- Rocha-Olivares, A.J.; Fleeger, J.W.; Foltz, D.W. Decoupling of molecular and morphological evolution in deep lineages of a meiobenthic harpacticoid copepod. Mol. Biol. Evol. 2001, 18, 1088–1102. [Google Scholar] [CrossRef]
- Gilbert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional biodiversity. BioScience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Deharveng, L.; Stoch, F.; Gibert, J.; Bedos, A.; Galassi, D.M.P.; Zagmajster, M.; Brancelj, A.; Camacho, A.; Giani, N.; Magniez, G.; et al. Groundwater biodiversity in Europe. Freshw. Biol. 2009, 54, 709–726. [Google Scholar] [CrossRef]
- Boonyanusith, C.; Brancelj, A.; Sanoamuang, L. First representatives of the genus Fierscyclops Karanovic, 2004 (Copepoda, Cyclopidae) from South East Asia. J. Limnol. 2013, 72 (Suppl. 2), 275–289. [Google Scholar] [CrossRef]
- Bruno, M.C.; Cottarelli, V. First record of Kinnecaris (Copepoda: Harpacticoida: Parastenocarididae) from Turkey and Thailand; description of three new species and emended definition of the genus. Ital. J. Zool. 2015, 82, 69–94. [Google Scholar] [CrossRef]
- Narita, S.; Pereira, R.A.S.; Kjellberg, F.; Kageyama, D. Gynandromorphs and intersexes: Potential to understand the mechanism of sex determination in arthropods. Terr. Arthropod Rev. 2010, 3, 63–96. [Google Scholar] [CrossRef]
- Fusco, G.; Minelli, A. Descriptive versus causal morphology: Gynandromorphism and intersexuality. Theory Biosci. 2023, 142, 1–11. [Google Scholar] [CrossRef]
- Gusmão, L.F.M.; McKinnon, A.D. Sex ratios, intersexuality and sex change in copepods. J. Plankton Res. 2009, 31, 1101–1117. [Google Scholar] [CrossRef]
- Moore, C.G.; Stevenson, J.M. The occurrence of intersexuality in harpacticoid copepods and its relationship with pollution. Mar. Pollut. Bull. 1991, 22, 72–74. [Google Scholar] [CrossRef]
- Moore, C.G.; Stevenson, J.M. Intersexuality in benthic harpacticoid copepods in the Firth of Forth, Scotland. J. Nat. Hist. 1994, 28, 1213–1230. [Google Scholar] [CrossRef]
- Klie, W. Ein gynandromorpher Amphiascus (Cop. Harp.) von Helgoland. Zool. Anz. 1944, 145, 77–79. [Google Scholar]
- Huys, R.; Gee, M.G. A revision of Danielssenia Boeck and Psammis Sars with the establishment of two new genera Archisenia and Bathypsammis (Harpacticoida: Paranannopidae). Bull. Br. Mus. Nat. Hist. Zool. 1993, 59, 83–94. [Google Scholar]
- Gómez, S. On some new species of Stenheliinae Brady, 1880 (Copepoda, Harpacticoida, Miraciidae) from north-western Mexico, with the proposal of Lonchoeidestenhelia gen. nov. ZooKeys 2020, 987, 41–79. [Google Scholar] [CrossRef]
- Hamond, R. Non-marine harpacticoid copepods of Auatralia. I. Camthocamptidae of the genus Camthocamptus Westwood s. lat. and Fibulacamtus, gen. nov. and including the description of a related new species of Camthocamptus from New Caledonia. Invertebr. Taxon. 1987, 1, 1023–1247. [Google Scholar] [CrossRef]
- Gutierrez-Aguirre, M.; Suarez-Morales, E.; Cervantes-Martínez, A.; Sarma, N.; Sarma, S.S.S. Morphology of Elaphoidella grandidieri (Guerne and Richard, 1893) (Copepoda, Harpacticoida) from Mexico with notes on fecundity in culture conditions. In Studies on Freshwater Copepoda: A Volume in Honour of Bernard Dussart; Defaye, D., Suarez-Morales, E., Von Vaupel Klein, C., Eds.; Koninklijke Brill: Leiden, The Netherlands, 2011; pp. 227–244. [Google Scholar] [CrossRef]
- Fuentes-Reines, J.M.; de Roa, E.Z. Occurrence of Elaphoidella grandidieri (Guerne and Richard, 1893) (Crustacea: Copepoda: Harpacticoida) in Ciénaga Grande de Santa Marta, Colombia. Check List 2013, 9, 1580–1583. [Google Scholar] [CrossRef][Green Version]
- Guerne, J.; Richard, J. Canthocamptus grandidieri, Alona cambouei, nouveaux Entomostracés d’eau douce de Madagascar. Mém. Soc. Zool. Fr. 1893, 6, 214–224. [Google Scholar]
- Ishida, T. Copepods in the mountain waters of Kyushu, Tsushima and Ryukyu Islands, southwestern Japan. Sci. Rep. Hokkaido Fish Hatch. 1990, 44, 39–51. [Google Scholar]
- Shen, C.J.; Tai, A.Y.; Li, A.Y.; Song, D.X.; Chang, C.Z.; Song, Y.Z.; Chen, G.X. Fauna Sinica, Crustacea: Freshwater Copepoda; Science Press: Beijing, China, 1979. [Google Scholar]
- Defaye, D. Contribution à la connaissance des Crustacés Copépodes d’Ethiopie. Hydrobiologia 1988, 164, 103–147. [Google Scholar] [CrossRef]
- Sars, G.O. Pacifische Plankton-Crustaceen. (Ergebnisse einer Reise nach dem Pacific Schaunsland 1896–1897). Zool. Jahrb. Abt. Syst. Oekol. Geogr. Tiere 1904, 19, 629–646. [Google Scholar]
- Kikuchi, Y. Redescription of a freshwater harpacticoid copepod, Elaphoidella grandidieri (Guerne & Richard, 1893), from a swamp at Itako, central Japan. Publ. Itako Hydrobiol. Stat. 1985, 2, 1–8. [Google Scholar]
- Watiroyram, S.; Sanoamuang, L.; Brancelj, A. A new species of Elaphoidella Chappuis, 1929 and Schizopera Sars, 1905 (Copepoda: Harpacticoida) from two caves in northeastern and southern Thailand. Zootaxa 2021, 5051, 550–569. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basis | Exopod | Endopod | |||||
|---|---|---|---|---|---|---|---|
| Leg | 1 | 2 | 3 | 1 | 2 | 3 | |
| P1 (female and male) | 1-I | I-0 | I-1 | I-I,2-0 | 0-1 | 0-1 | 0-I,1-1 |
| P2 | 1-0 | I-0 | I-1 | II-2-1 | 0-0 | 0-2-0 | |
| P2 (male) | 1-0 | I-0 | I-1 | II-2-1 | 0-0 | 0-0-0 | |
| P3 | 1-0 | I-0 | I-1 | II-2-1 | 0-0 | I-I,1-2 | |
| P3 (male) | 1-0 | I-0 | I-1 | II-2-1 | 0-0 | 0-0 (apophysis) | 0-I-0 |
| P4 | 1-0 | I-0 | I-1 | II-2-1 | 0-1-0 | ||
| P4 (male) | 1-0 | I-0 | I-1 | II-I,1-1 | 0-1-0 | ||
| Taxa | Habitat | Distribution |
|---|---|---|
| E. botosaneanui Petkovski, 1973 | cave | Cuba |
| E. bromeliaecola (Chappuis, 1928) | phytotelmata, cave | Indonesia (Java, Sumatra), Thailand |
| E. crenobia Petkovski, 1973 | cave | Cuba |
| E. einslei Petkovski, 1973 | cave | Cuba |
| E. humboldti Löffler, 1963 | phytotelmata | Ecuador, Colombia |
| E. jochenmartensi Dumont and Maas, 1988 | leaf litter | Nepal |
| E. kieferi Löffler, 1968 | wet moss | Nepal |
| E. labani Löffler, 1973 | creek, wet moss | Malaysia (Borneo) |
| E. negroensis Kiefer, 1967 | groundwater (interstitial) | Brazil |
| E. prohumboldti Petkovski, 1980 | cave | Cuba |
| E. sanoamuangae Watiroyram and Brancelj, 2016 | cave | Thailand |
| E. schubarti Chappuis, 1936 | wet moss, spring | Brazil, Colombia |
| E. subcrenobia Petkovski, 1980 | cave | Cuba |
| E. thienemanni thienemanni Chappuis, 1931 | phytotelmata (near stream) | Indonesia (Sumatra) |
| E. thienemanni serrulata Chappuis, 1931 | floating-leaf plant | Indonesia (Sumatra) |
| E. turgisetosa Petkovski, 1980 | cave | Cuba |
| Character | E. phuphamanensis sp. nov. | E. sanoamuangae | E. bromeliaecola | E. thienemanni | E. laboni |
|---|---|---|---|---|---|
| Ornamentation of free margin of anal operculum (both sexes) | 10–12 robust spinules | 8–10 strong spinules | 10–14 strong spinules | 12–14 strong spinules | numerous fine spinules |
| Number of segments of P1 Enp (both sexes) | 3 | 2 | 3 | 3 | 3 |
| Number of segments of P4 Enp (both sexes) | 1 | 2 | 2 | 2 | 2 |
| Armature of P2–P4 Enp-1 (female) | 0.0.0 | 0.0.0 | 1.1.0 | 1.1.0 | 1.1.0 |
| Armature of P2–P4 Enp-2 (female) | 2.3.1 | 3.5.3 | 4.3.4 | 4.5.4 | 4.5.2–3 |
| Armature of P2–P4 Exp-3 (female) | 5.5.5 | 5.6.6 | 5.6.6 | 5.6.6 | 5.6.6 |
| Armature of P5 baseoendopodal lobe and Exp (female) | 4.3 | 4.4 | 4.4 | 4.4 | 4.4 |
| Prominence at tip of baseoendopod of P5 where seta II inserts (female) | absent | present | present | present | present |
| Armature of distal segment of P2–P4 Enp (male) | 0.1.1 | 2.2.3 | 3.2.3 | 3.2.3 | 2.2.2 |
| Armature of P3 Enp-3 (male) | sword-like seta | two setae, subequal in length | two setae, subequal in length | thick blunt spine and one longer seta | two setae, subequal in length |
| Number of elements on the P4 Exp-3 (male) | 5 | 6 | 6 | 6 | 6 |
| Number of elements on the P5 Exp (male) | 2 | 3 | 3 | 3 | 2–3 |
| Basis | Exopod | Endopod | |||||
|---|---|---|---|---|---|---|---|
| Leg | 1 | 2 | 3 | 1 | 2 | 3 | |
| P1 (female and male) | I-1 | I-0 | I-1 | I-I,1-1 | 0-1 | 0-1 | 0-I,1-1 |
| P2 | I-0 | I-0 | I-1 | II-2-1 | 0-0 | I-2-2 | |
| P2 (male) | I-0 | I-0 | I-1 | II-2-1 | 0-0 | 0-2-2 | |
| P3 | 1-0 | I-0 | I-1 | II-2-2 | 0-0 | I-2-2 | |
| P3 (male) | 1-0 | I-0 | I-1 | II-2-2 | 0-0 | 0-0 (apophysis) | 0-1-1 |
| P4 | 1-0 | I-0 | I-1 | II-2-2 | 0-0 | I-1-2 | |
| P4 (male) | 1-0 | I-0 | I-1 | II-2-2 | 0-0 | 0-I,1-1 | |
| Character | E. propecabezasi sp. nov. | E. cabezasi | E. paraaffinis | E. ligorae | E. margaritae |
|---|---|---|---|---|---|
| Free margin of hyaline frill on urosomites 2–4 | serrated | smooth | serrated | serrated | smooth? |
| Ornamentation of free margin of anal operculum | 20–25 short spinules | smooth | 24–27 oblong spinules | numerous fine spinules | 13–16 strong spinules |
| Shape of caudal rami | conical; about 1.7 times as long as wide | conical; about 1.8 times as long as wide | rectangular; as long as wide | conical; about 1.5 times as long as wide | subconical about 1.4 times as long as wide |
| Relative length of P1 Exp | reaching tip of P1 Enp-2 | reaching tip of P1 Enp-2 | reaching 1/2 length of P1 Enp-3 | reaching tip of P1 Enp-2 | reaching 1/2 length of P1 Enp-2 |
| Armature of P2–P4 Enp-1 (female) | 0.0.0 | 0.0.0 | 0.0.0 | 1.0.0 | 0.0.0 |
| Armature of P2–P4 Exp-3 (female) | 5.6.6 | 5.6.6 | 5.6.6 | 5.6.6 | 5.6.6 |
| Armature of P2–P4 Enp-2 (female) | 5.5.4 | 5.5.4 | 5.5.4 | 5.5.4 | 5.5.3 |
| Number of elements on P5 baseoendopod Exp (female) | 4.3 | 4.4 | 4.4 | 4.4 | 4.2 |
| Relative length of seta IV on P5 baseoendopod (female) | >1/2 length of seta II | >1/2 length of seta II | <1/2 length of seta II | >1/2 length of seta II | >1/2 length of seta II |
| Armature of P2–P4 distal segment of Enp (male) | 4.2.3 | 4.2.3 | 4.2.3 | 5.2.3 | 3.2.3 |
| Number of elements on P5 Exp (male) | 3 | 4 | 4 | 4 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonyanusith, C.; Brancelj, A.; Sanoamuang, L. Two New Species of Elaphoidella (Copepoda, Harpacticoida) from Subterranean Waters in Northeast Thailand, with a Record of a Gynandromorphic Specimen and an Up-to-Date Key to Elaphoidella Species from Southeast Asia. Diversity 2024, 16, 185. https://doi.org/10.3390/d16030185
Boonyanusith C, Brancelj A, Sanoamuang L. Two New Species of Elaphoidella (Copepoda, Harpacticoida) from Subterranean Waters in Northeast Thailand, with a Record of a Gynandromorphic Specimen and an Up-to-Date Key to Elaphoidella Species from Southeast Asia. Diversity. 2024; 16(3):185. https://doi.org/10.3390/d16030185
Chicago/Turabian StyleBoonyanusith, Chaichat, Anton Brancelj, and Laorsri Sanoamuang. 2024. "Two New Species of Elaphoidella (Copepoda, Harpacticoida) from Subterranean Waters in Northeast Thailand, with a Record of a Gynandromorphic Specimen and an Up-to-Date Key to Elaphoidella Species from Southeast Asia" Diversity 16, no. 3: 185. https://doi.org/10.3390/d16030185
APA StyleBoonyanusith, C., Brancelj, A., & Sanoamuang, L. (2024). Two New Species of Elaphoidella (Copepoda, Harpacticoida) from Subterranean Waters in Northeast Thailand, with a Record of a Gynandromorphic Specimen and an Up-to-Date Key to Elaphoidella Species from Southeast Asia. Diversity, 16(3), 185. https://doi.org/10.3390/d16030185