
Microclimatic Influences on the Abundance of Three Non-Troglobiont Species




Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats, 2nd ed.; Oxford University Press: New York, NY, USA, 2019. [Google Scholar]
- Badino, G. Underground Meteorology—“What’s the Weather Underground”? Acta Carsologica 2010, 39, 427–448. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats: Ecology, Evolution, and Conservation; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Seasonal Variation in Microhabitat of Salamanders: Environmental Variation or Shift of Habitat Selection? PeerJ 2015, 3, e1122. [Google Scholar] [CrossRef]
- Mammola, S. Finding Answers in the Dark: Caves as Models in Ecology Fifty Years after Poulson and White. Ecography 2019, 42, 1331–1351. [Google Scholar] [CrossRef]
- Monro, A.K.; Bystriakova, N.; Fu, L.; Wen, F.; Wei, Y. Discovery of a Diverse Cave Flora in China. PLoS ONE 2018, 13, e0190801. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.L.; Martins, R.P. Trophic Structure and Natural History of Bat Guano Invertebrate Communities, with Special Reference to Brazilian Caves. Trop. Zool. 1999, 12, 231–252. [Google Scholar] [CrossRef]
- Lavoie, K.H.; Helf, K.L.; Poulson, T.L. The Biology and Ecology of North American Cave Crickets. J. Cave Karst Stud. 2007, 69, 114–134. [Google Scholar]
- Lunghi, E.; Valle, B.; Guerrieri, A.; Bonin, A.; Cianferoni, F.; Manenti, R.; Ficetola, G.F. Complex Patterns of Environmental DNA Transfers from Surface to Subterranean Soils Revealed by Analyses of Cave Insects and Springtails. Sci. Total Environ. 2022, 826, 154022. [Google Scholar] [CrossRef] [PubMed]
- Howarth, F.G.; Moldovan, O.T. The Ecological Classification of Cave Animals and Their Adaptations. In Cave Ecology; Moldovan, O.T., Kováč, Ľ., Halse, S., Eds.; Springer: Berlin, Germany, 2018; pp. 41–67. [Google Scholar]
- Fenolio, D.B.; Niemiller, M.L.; Bonett, R.M.; Graening, G.O.; Collier, B.A.; Stout, J.F. Life History, Demography, and the Influence of Caveroosting Bats on a Population of the Grotto Salamander (Eurycea Spelaea) from the Ozark Plateaus of Oklahoma (Caudata: Plethodontidae). Herpetol. Conserv. Biol. 2014, 9, 394–405. [Google Scholar]
- Simon, K.S.; Pipan, T.; Culver, D.C. A Conceptual Model of the Flow and Distribution of Organic Carbon in Caves. J. Cave Karst Stud. 2007, 69, 279–284. [Google Scholar]
- Ficetola, G.F.; Canedoli, C.; Stoch, F. The Racovitzan Impediment and the Hidden Biodiversity of Unexplored Environments. Conserv. Biol. 2019, 33, 214–216. [Google Scholar] [CrossRef]
- Bilandžija, H.; Ćetković, H.; Jeffery, W.R. Evolution of Albinism in Cave Planthoppers by a Convergent Defect in the First Step of Melanin Biosynthesis. Evol. Dev. 2012, 14, 196–203. [Google Scholar] [CrossRef]
- Gonzalez, B.C.; Worsaae, K.; Fontaneto, D.; Martínez, A. Anophthalmia and Elongation of Body Appendages in Cave Scale Worms (Annelida: Aphroditiformia). Zool. Scr. 2017, 47, 106–121. [Google Scholar] [CrossRef]
- Mack, K.L.; Jaggard, J.B.; Persons, J.L.; Roback, E.Y.; Passow, C.N.; Stanhope, B.A.; Ferrufino, E.; Tsuchiya, D.; Smith, S.E.; Slaughter, B.D.; et al. Repeated Evolution of Circadian Clock Dysregulation in Cavefish Populations. PLoS Genet. 2021, 17, e1009642. [Google Scholar] [CrossRef]
- Pasquali, V.; Sbordoni, V. High Variability in the Expression of Circadian Rhythms in a Cave Beetle Population. Biol. Rhythm. Res. 2014, 45, 925–939. [Google Scholar] [CrossRef]
- Hyacinthe, C.; Attia, J.; Rétaux, S. Evolution of Acoustic Communication in Blind Cavefish. Nat. Commun. 2019, 10, 4231. [Google Scholar] [CrossRef]
- Plath, M.; Parzefall, J.; Körner, K.E.; Schlupp, I. Sexual Selection in Darkness? Female Mating Preferences in Surface- and Cave-Dwelling Atlantic Mollies, Poecilia mexicana (Poeciliidae, Teleostei). Behav. Ecol. Sociobiol. 2004, 55, 596–601. [Google Scholar] [CrossRef]
- Christiansen, K. Proposition Pour La Classification Des Animaux Cavernicoles. Spelunca 1962, 2, 76–78. [Google Scholar]
- Hervant, F. Starvation in Subterranean Species versus Surface-Dwelling Species: Crustaceans, Fish, and Salamanders. In Comparative Physiology of Fasting, Starvation, and Food Limitation; McCue, M.D., Ed.; Springer: Berlin, Germany, 2012; pp. 91–102. [Google Scholar]
- Lunghi, E.; Bilandžija, H. Longevity in Cave Animals. Front. Ecol. Evol. 2022, 10, 874123. [Google Scholar] [CrossRef]
- Lunghi, E.; Mammola, S.; Martinez, A.; Hesselberg, T. Behavioural Adjustments Enable the Colonization of Subterranean Environments. Zool. J. Linn. Soc. Lond. 2023, zlad133. [Google Scholar] [CrossRef]
- Kirn, T. Finds of Washed-out Proteus from the Pivka Intermittent Lakes and the Pivka River. Nat. Slov. 2018, 20, 51–56. [Google Scholar] [CrossRef]
- Manenti, R.; Di Nicola, M.R.; Zampieri, V.; Grassi, G.; Creanza, T.; Mauri, E.; Ficetola, G.F.; Barzaghi, B. Wandering Outside of the Styx: Surface Activity of an Iconic Subterranean Vertebrate, the Olm (Proteus anguinus). Ecology 2024, 105, e4252. [Google Scholar] [CrossRef] [PubMed]
- Buzzacott, P.L.; Zeigler, E.; Denoble, P.; Vann, R. American Cave Diving Fatalities 1969–2007. Int. J. Aquat. Res. Educ. 2009, 3, 162–177. [Google Scholar] [CrossRef]
- Iliffe, T.M.; Bowen, C. Scientific Cave Diving. Mar. Technol. Soc. J. 2001, 35, 36–41. [Google Scholar] [CrossRef]
- Zagmajster, M.; Culver, D.C.; Christman, M.C.; Sket, B. Evaluating the Sampling Bias in Pattern of Subterranean Species Richness: Combining Approaches. Biodivers. Conserv. 2010, 19, 3035–3048. [Google Scholar] [CrossRef]
- Rizzo, V.; Sánchez-Fernández, D.; Fresneda, J.; Cieslak, A.; Ribera, I. Lack of Evolutionary Adjustment to Ambient Temperature in Highly Specialized Cave Beetles. BMC Evol. Biol. 2015, 15, 10. [Google Scholar] [CrossRef]
- Fenolio, D.B.; Graening, G.O.; Collier, B.A.; Stout, J.F. Coprophagy in a Cave-Adapted Salamander; the Importance of Bat Guano Examined through Nutritional and Stable Isotope Analyses. Proc. R. Soc. B 2006, 273, 439–443. [Google Scholar] [CrossRef]
- Guillaume, O. Surface Newt Calotriton Asper Acclimation to Cave Conditions Improved Their Foraging Ability in Darkness. Front. Ecol. Evol. 2022, 10, 1057023. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Do Cave Features Affect Underground Habitat Exploitation by Non-Troglobite Species? Acta Oecologica 2014, 55, 29–35. [Google Scholar] [CrossRef]
- Lunghi, E. Occurrence of the Black Lace-Weaver Spider, Amaurobius Ferox, in Caves. Acta Carsologica 2020, 49, 119–124. [Google Scholar] [CrossRef]
- Venarsky, M.P.; Huntsman, B.M.; Huryn, A.D.; Benstead, J.P.; Kuhajda, B.R. Quantitative Food Web Analysis Supports the Energy Limitation Hypothesis in Cave Stream Ecosystems. Oecologia 2014, 176, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Mulargia, M.; Veith, M.; Corti, C.; Ficetola, G.F. Environmental Suitability Models Predict Population Density, Performance and Body Condition for Microendemic Salamanders. Sci. Rep. 2018, 8, 7527. [Google Scholar] [CrossRef]
- Barzaghi, B.; Ficetola, G.F.; Pennati, R.; Manenti, R. Biphasic Predators Provide Biomass Subsidies in Small Freshwater Habitats: A Case Study of Spring and Cave Pools. Freshw. Biol. 2017, 62, 1637–1644. [Google Scholar] [CrossRef]
- Chapin, K.J. Guano Deposition Predicts Top Predator (Amblypygi: Phrynidae) Abundance in Subtropical Caves. J. Arachnol. 2019, 47, 385–388. [Google Scholar] [CrossRef]
- Salgado, S.S.; Motta, P.C.; De Souza Aguiar, L.M.; Nardoto, G.B. Tracking Dietary Habits of Cave Arthropods Associated with Deposits of Hematophagous Bat Guano: A Study from a Neotropical Savanna. Austral Ecol. 2014, 39, 560–566. [Google Scholar] [CrossRef]
- Acevedo, P.; Escudero, M.A.; Muńoz, R.; Gortázar, C. Factors Affecting Wild Boar Abundance across an Environmental Gradient in Spain. Acta Theriol. 2006, 51, 327–336. [Google Scholar] [CrossRef]
- Amarasekare, P.; Coutinho, R.M. Effects of Temperature on Intraspecific Competition in Ectotherms. Am. Nat. 2014, 184, E50–E65. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E. Ecology and Life History of Meta bourneti (Araneae: Tetragnathidae) from Monte Albo (Sardinia, Italy). PeerJ 2018, 6, e6049. [Google Scholar] [CrossRef]
- Lunghi, E.; Bruni, G.; Ficetola, G.F.; Manenti, R. Is the Italian Stream Frog (Rana italica Dubois, 1987) an Opportunistic Exploiter of Cave Twilight Zone? Subterr. Biol. 2018, 25, 49–60. [Google Scholar] [CrossRef]
- Lipovšek, S.; Leitinger, G.; Janžekovič, F.; Kozel, P.; Dariš, B.; Perc, M.; Devetak, D.; Weiland, N.; Novak, T. Towards Understanding Partial Adaptation to the Subterranean Habitat in the European Cave Spider, Meta menardi: An Ecocytological Approach. Sci. Rep. 2019, 9, 9121. [Google Scholar] [CrossRef]
- Campbell Grant, E.H.; Lowe, W.H.; Fagan, W.F. Living in the Branches: Population Dynamics and Ecological Processes in Dendritic Networks. Ecol. Lett. 2007, 10, 165–175. [Google Scholar] [CrossRef]
- Mammola, S.; Isaia, M. Spiders in Cave. Proc. R. Soc. B 2017, 284, 20170193. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Cave Features, Seasonality and Subterranean Distribution of Non-Obligate Cave Dwellers. PeerJ 2017, 5, e3169. [Google Scholar] [CrossRef] [PubMed]
- Bank, R.A.; Neubert, E. MolluscaBase. Checklist of the Land and Freshwater Gastropoda of Europe. Available online: https://www.marinespecies.org/traits./aphia.php?p=sourcedetails&id=279050 (accessed on 1 March 2024).
- Lunghi, E.; Corti, C.; Mulargia, M.; Zhao, Y.; Manenti, R.; Ficetola, G.F.; Veith, M. Cave Morphology, Microclimate and Abundance of Five Cave Predators from the Monte Albo (Sardinia, Italy). Biodivers. Data J. 2020, 8, e48623. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling. Inferring Patterns and Dynamics of Species Occurrence; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- Crump, M.L.; Scott, N.J. Visual Encounter Surveys. In Measuring and Monitoring Biological Diversity: Standard Methods for Amphibians; Heyer, W.R., Donnelly, M.A., McDiarmid, R.W., Hayek, L.C., Foster, M.S., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1994; pp. 84–92. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference. Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria.
- Douglas, B.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barke, R.J.; Schofield, M.R.; Link, W.A.; Sauer, J.R. On the Reliability of N-Mixture Models for Count Data. Biometrics 2017, 74, 369–377. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Barzaghi, B.; Melotto, A.; Muraro, M.; Lunghi, E.; Canedoli, C.; Lo Parrino, E.; Nanni, V.; Silva-Rocha, I.; Urso, A.; et al. N-Mixture Models Reliably Estimate the Abundance of Small Vertebrates. Sci. Rep. 2018, 8, 10357. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Ficetola, G.F.; Zhao, Y.; Manenti, R. Are the Neglected Tipuloidea Crane Flies (Diptera) an Important Component for Subterranean Environments? Diversity 2020, 12, 333. [Google Scholar] [CrossRef]
- Manenti, R.; Lunghi, E.; Ficetola, G.F. Distribution of Spiders in Cave Twilight Zone Depends on Microclimatic Features and Trophic Supply. Invertebr. Biol. 2015, 134, 242–251. [Google Scholar] [CrossRef]
- Curry, P.A.; Yeung, N.W. Predation on Endemic Hawaiian Land Snails by the Invasive Snail Oxychilus alliarius. Biodivers. Conserv. 2013, 22, 3165–3169. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, B.; Christensen, H.B. lmerTest: Tests in Linear Mixed Effects Models; R Package, Version 2.0–2.9; 2016. Available online: www.r-project.org (accessed on 10 December 2023).
- Ficetola, G.F.; Romano, A.; Salvidio, S.; Sindaco, R. Optimizing Monitoring Schemes to Detect Trends in Abundance over Broad Scales. Anim. Conserv. 2017, 21, 221–231. [Google Scholar] [CrossRef]
- Pekár, S.; Líznarová, E.; Řezáč, M. Suitability of woodlice prey for generalist and specialist spider predators: A comparative study. Ecol. Entomol. 2016, 41, 123–1300. [Google Scholar] [CrossRef]
- Nyffeler, M.; Moor, H.; Foelix, R.F. Spiders feeding on earthworms. J. Arachnol. 2001, 29, 119–124. [Google Scholar] [CrossRef]
- Ferreira, R.L.; Mendes Martins, V.; Arantes Paixão, E.; Souza Silva, M. Spatial and Temporal Fluctuations of the Abundance of Neotropical Cave-Dwelling Moth Hypena sp. (Noctuidae, Lepidoptera) Influenced by Temperature and Humidity. Subterr. Biol. 2015, 16, 47–60. [Google Scholar] [CrossRef]
- Hesselberg, T.; Simonsen, D.; Juan, C. Do Cave Orb Spiders Show Unique Behavioural Adaptations to Subterranean Life? A Review of Evidence from Cave Orb Spiders. Behaviour 2019, 156, 969–996. [Google Scholar] [CrossRef]
- Novak, T.; Tkavc, T.; Kuntner, M.; Arnett, A.E.; Lipovšek Delakorda, S.; Perc, M.; Janžekovič, F. Niche Partitioning in Orbweaving Spiders Meta menardi and Metellina merianae (Tetragnathidae). Acta Oecologica 2010, 36, 522–529. [Google Scholar] [CrossRef]
- Novak, T.; Sambol, J.; Janžekovič, F. Faunal Dynamics in the Železna Jama Cave. Acta Carsologica 2004, 33, 249–267. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What Shapes the Trophic Niche of European Plethodontid Salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef] [PubMed]
- Manenti, R. Dry stone walls favour biodiversity: A case-study from the Appennines. Biodiv. Conserv. 2014, 23, 1879–1893. [Google Scholar] [CrossRef]
- Ferreira, A.F.; Martins, A.M.d.F.; da Cunha, R.T.; Melo, P.J.; Rodrigues, A.R. Reproductive Biology of Oxychilus (Atlantoxychilus) Spectabilis (Milne-Edwards, 1885) (Gastropoda: Pulmonata): A Gametogenic Approach. Arquipélago Life Mar. Sci. 2013, 30, 11–17. [Google Scholar]
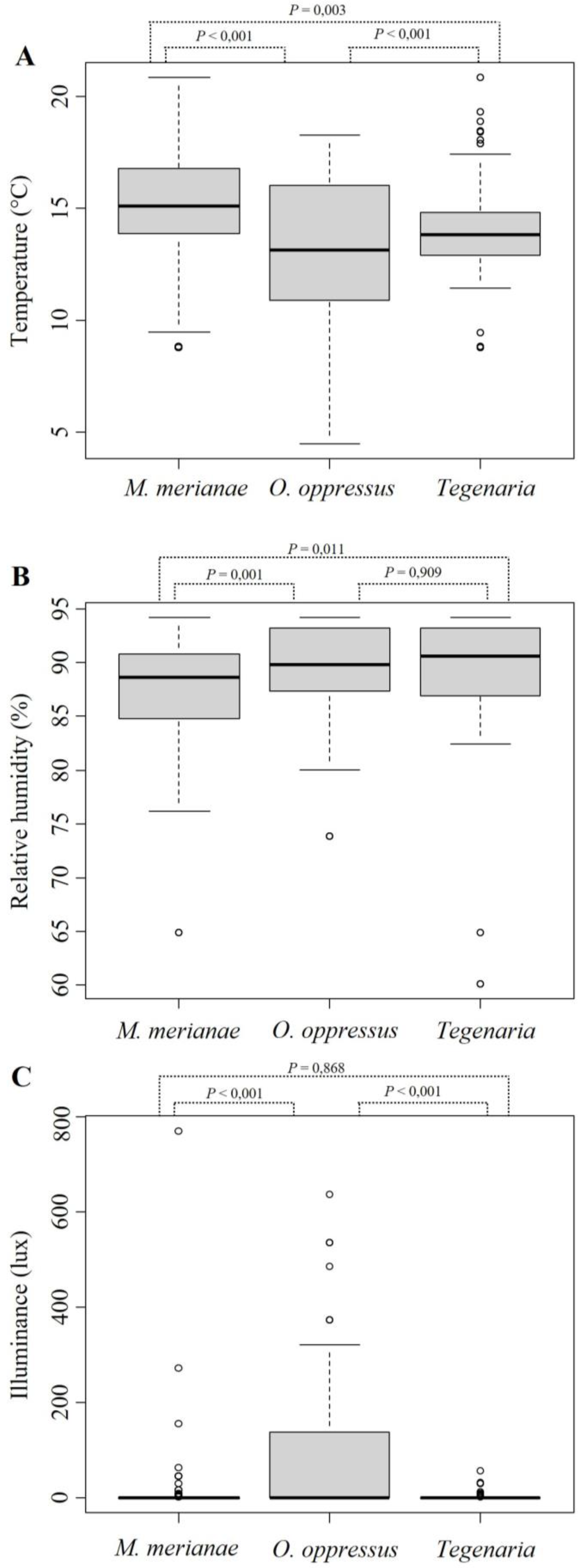

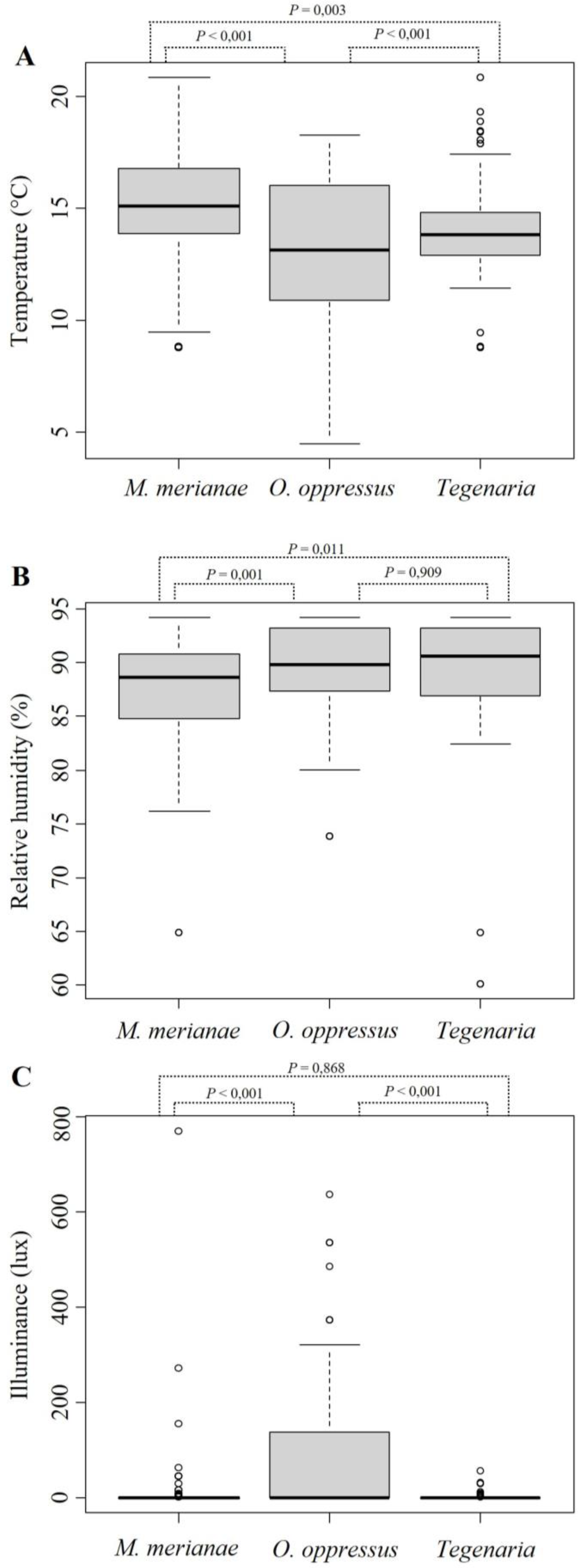{kind=link}
| Species | Model | Type of Covariate | AICc | Covariate Effect (±SE) | Detection Probability (±SE) |
|---|---|---|---|---|---|
| Metellina merianae | 1 | None | 879.14 | ||
| 2 | Sector depth | 853.59 | −0.05 (±0.01) | 0.195 (±0.02) | |
| 3 | Season | 864.93 | |||
| Tegenaria sp. | 1 | None | 786.11 | ||
| 2 | Sector depth | 775.8 | −0.02 (±0.01) | 0.209 (±0.04) | |
| 3 | Season | 785.24 | |||
| Oxychilus oppressus | 1 | None | 842.68 | 0.358 (±0.05) | |
| 2 | Sector depth | 843.19 | |||
| 3 | Season | 845.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppari, L.; Manenti, R.; Lunghi, E. Microclimatic Influences on the Abundance of Three Non-Troglobiont Species. Diversity 2024, 16, 186. https://doi.org/10.3390/d16030186
Coppari L, Manenti R, Lunghi E. Microclimatic Influences on the Abundance of Three Non-Troglobiont Species. Diversity. 2024; 16(3):186. https://doi.org/10.3390/d16030186
Chicago/Turabian StyleCoppari, Luca, Raoul Manenti, and Enrico Lunghi. 2024. "Microclimatic Influences on the Abundance of Three Non-Troglobiont Species" Diversity 16, no. 3: 186. https://doi.org/10.3390/d16030186
APA StyleCoppari, L., Manenti, R., & Lunghi, E. (2024). Microclimatic Influences on the Abundance of Three Non-Troglobiont Species. Diversity, 16(3), 186. https://doi.org/10.3390/d16030186