Abstract
During the study of diversity in filamentous cyanobacteria in China, two strains (WZU0719 and WZU0723) with the form of thin filaments were isolated from the surface of Qiandao Lake, a large freshwater lake in Zhejiang Province, China. A comprehensive analysis was conducted, incorporating morphological, ecological, and molecular data. The morphological examination provided an initial identification as a Leptolyngbya-like cyanobacterium. Genetic characterization was also performed by amplifying the 16S rRNA gene and the 16S-23S rRNA internal transcribed spacer (ITS) region. The phylogenetic grouping based on the 16S rRNA gene demonstrates that the examined strain is unequivocally assigned to the Pegethrix genus. However, it possesses distinct phylogenetic divergence from the six described Pegethrix species. Additionally, discrepancies in habitat further differentiate it from other members of this genus. Employing the polyphasic approach, we present a comprehensive account of the newly discovered taxa: Pegethrix qiandaoensis sp. nov. The novel taxonomic finding in this research significantly contributes to enhancing the comprehension of Pegethrix diversity across various habitats.
1. Introduction
The wide distribution of cyanobacteria, with the nature of microorganisms, makes them an integral component of various ecosystems, including soil and aquatic environments [1,2]. In taxonomic history, cyanobacteria had long been classified based on their morphological characteristics but recent advancements in studying their molecular phylogenies have led to significant revisions [3,4,5]. Many of the morphological characteristics that were traditionally used as indicators of phylogeny have been proven to be variable. Additionally, phylogenetic reconstructions demonstrated that a large number of genera previously regarded to have consistent morphological traits were polyphyletic [6,7]. These polyphyletic genera, sometimes referred to as cryptogenera, may have resulted from multiple lineages of cyanobacteria undergoing frequent evolutionary convergence [8,9,10].
The 16S rRNA gene is commonly used as a molecular marker for identifying prokaryotes [11,12], especially in taxonomic studies to determine their assignment at genus level. Moreover, the practical use of secondary structures in the 16S–23S rRNA internal transcribed spacer (ITS) is considered advantageous for discovering new species of cyanobacteria [13,14]. After several major revisions, the current cyanobacterial classification has changed greatly [15,16], including new orders and families proposed to satisfy the requirement of monophyly at all taxonomic ranks. The newly proposed family Oculatellaceae has been demonstrated to make outstanding contributions to further clarifying the relationship between Leptolyngbyales and Pseudanabaenales [17]. Oculatellaceae, primarily consisting of diverse subaerophytic taxa found in wet rocks and soils, was phylogenetically grouped as a sister clade to Pseudanabaenales, with high support [17].
Among the recently described aerophytic genera, Pegethrix stands out with its six characterized species. These species have been thoroughly examined in terms of morphology, genetics, and ecology. Interestingly, most of these species could form nodules in their filaments under low light conditions. Furthermore, a species recently discovered in China has been found to exhibit an additional capability of inducing chlorophyll f synthesis under far-red light (FRL) [18]. At present, all species in the genus come from wet rocks and soil crusts near streams [17,18,19].
In this study, we isolated two thin filamentous cyanobacterial strains (WZU0719 and WZU0723) from the surface of a large freshwater lake in Zhejiang Province, China. We used a polyphasic approach, including molecular characteristics, morphology, and ecological data, to evaluate the taxonomic status of the strains. Initial morphological examination temporarily identified the strains as a Leptolyngbya-related morphotype. Molecular analyses, however, determined their position within the genus Pegethrix and further supported them as a new species. The main objectives of this study were to clarify the phylogenetic position of the two strains. In addition, the novel species possesses a distinctive habitat, setting it apart from other species within the genus. To differentiate it from other Pegethrix species and establish it as a new species, we have named it Pegethrix qiandaoensis sp. nov. based on the nomenclature of Algae, Fungi, and Plants. This study presents a taxonomic description of the newly discovered species.
2. Materials and Methods
2.1. Strain Collection and Cultivation
In June 2020, samples were collected from the surface of Qiandao Lake in Zhejiang Province, China (118°34′ E, 29°22′ N). Samples were transported back to the laboratory and placed in a sampling refrigerator at 4 °C to maintain freshness. Clusters of filaments were separated within the field of view of a dissecting microscope (Carl Zeiss STEMI 508, Oberkochen, Germany) at 40× magnification using a laboratory-made Pasteur pipette. The isolated cell clusters were then placed in 24-well plates containing sterile modified liquid medium BG-11 [20]. After three to four weeks, pure culture was inspected for contaminants and transferred to a screw-cap tube containing 10 mL of the same medium to obtain a unialgal culture. Cultures were exposed to a 12 h light/12 h dark cycle at 25 °C and a photon flux density of 35 μmol·m−2·s−1. These cultures were deposited at the Algae Culture Collection Center, College of Life and Environmental Science, Wenzhou University, China. The accession numbers of these strains are WZU0719 and WZU0723, respectively.
2.2. Morphological Characterization
Live cultures were detected using a LEICA DM2000 LED microscope (LEICA, Wetzlar, Germany) and photomicrographs were taken using a LEICA DMC 5400 photomicrography system (digital camera attached to the microscope) (LEICA, Wetzlar, Germany). Images were analyzed using Leica Application Suite X 3.7.4. Cell size was measured from digital images obtained at 1000× magnification and the average vegetative cell width of more than 60 filaments was calculated.
2.3. DNA Extraction, PCR Amplification, and Sequencing
To avoid contamination by other bacteria, the cultured cells were centrifuged and washed three times with sterile phosphorus-free modified liquid medium BG-11. Total genomic DNA of these strains was extracted using a modified cetyltrimethylammonium bromide (CTAB) method [21]. Primers used for 16S-23S rRNA amplification include PA (5′-AGA GTT TGATCC TGG CTC AG-3′) and B23s (5′-CTT CGC CTC TGT GTG CCT AGG T-3′) [22,23]. The 16S rRNA PCR reaction volume was 50 μL, including 2 μL template DNA and 1 μL (10 μmol/L) of each primer, 22 μL sterile water, and 25 μL 2×Taq mix (Dye Plus) (Vazyme Biotech Co. Ltd., Nanjing, China). PCR amplification was performed using a SimpliAmp™ thermal cycler (San Francisco, CA, USA). The PCR profile consisted of an initial denaturation at 95 °C for 5 min, followed by 30 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 2 min, and a final extension step of 10 min at 72 °C. PCR products were purified using TIANgel Midi (Tiangen Biotechnology Co., Ltd., Beijing, China) purification kit. The purified PCR products were then cloned using the versatile simple vector kit pClone007 (Beijing Tsingke Biotechnology Co., Ltd., Beijing, China). The insert containing the plasmid in E. coli Trelidf™ 5α was replicated using chemically competent cells and incubated at 37 °C for 12–14 h. The clones that contained the target fragment were sequenced bidirectionally using standard plasmid primers M13F (5′-TGT AAA ACG ACG GCC AGT-3′) and M13R (5′-CAG GAA ACA GCT ATG ACC-3′), by Wuhan Tianyi Huayu Gene Technology Co., Ltd. (located in Wuhan, China).
2.4. Phylogenetic Analysis
The online tool Nucleotide BLAST was used to compare the 16S-23S rRNA gene sequences obtained in this study. Highly similar 16S-23S rRNA gene sequences were downloaded from the NCBI GenBank database for phylogenetic analysis. Downloaded and captured sequences were aligned using MAFFT v7.463 and trimmed using BioEdit v7.0.9 [24,25]. The phylogenetic tree of 16S rRNA gene outgroup was established using the sequence of Gloeobacter violaceus PCC7421. After multiple rounds of sequence comparison, a data matrix containing 1162 sequences and 79 nucleotide sites of the 16S rRNA gene was obtained. The best alternative model for the phylogenetic analysis was selected using the ModelFinder program in the molecular phylogenetic analysis platform PhyloSuite v1.2.2 [26]. Bayesian inference (BI) and maximum-likelihood (ML) analyses were performed based on the Akaike information criterion (AIC). For the 16S rRNA gene matrix, the nucleic acid substitution model (GTR + R4 + F) was chosen for optimal ML analysis, and the recommended fitting model (GTR + I + G + F) was used for BI analysis. The Pseudanabaena foetida (LC016779.1) sequence was used as the outgroup to construct the ITS gene phylogenetic tree. After multiple rounds of sequence comparison, a data matrix containing 1080 sequences and 48 nucleotide sites of the ITS gene was obtained. ITS gene matrix analysis used the nucleic acid substitution model (TVM + R5 + F) of maximum-likelihood analysis (ML) and the adaptation model (GTR + G + F) proposed by Bayesian analysis (BI) for phylogenetic analysis. The specific operating parameters of the surrogate model were estimated using IQ-TREE v2.1.3 and Mr-Bayes v3.2.7, respectively [27,28]. For ML analysis, bootstrapping was performed with 1000 pseudo-replicates on default options for relative support. The BI analysis consisted of two parallel runs of 10 million generations, with sampling every 100 generations. The first 25% of sampled data were discarded as burn-in. Neighbor-joining (NJ) analysis was performed using a two-parameter Kimura model with 1000 bootstrap replicates created in MEGA 11. To estimate the genetic distance of the 16S rRNA gene sequence similarity matrix, MEGA 11 used the Kimura 2-parameter model to calculate the p distance of the pairwise deletion gaps. Phylogenetic trees were visualized in Fig Tree v1.4.3, edited in Tree View 1.6.6 [29,30], and adjusted in Adobe Illustrator 2022 software. The sequences of 16S-23S rRNA gene obtained in this study were deposited in the GenBank database with accession numbers OR815386 and PP084669.
2.5. Morphological Ultrastructural Characterization
Samples were set with 2.5% glutaraldehyde in 0.1 M phosphate buffer with a pH of 7.2 and a temperature of 4 °C for a duration of three days to stabilize their filaments. Once completed, they underwent three rinses in 0.1 M phosphate buffer. Then, they were subjected to a post-fixation process involving 1% osmium tetroxide for a period of 2 h. Once fixed, the materials went through a dehydration process utilizing an ethanol gradient that progressed from 30% to 50%, then to 70%, 90%, and ultimately 100%. Lastly, the specimens were integrated into Spurr’s resin as per the procedure outlined by Spurr in 1969. To facilitate transmission electron microscope (TEM) examination, the samples were sectioned into 80 nm sections using a Leica UC6 ultramicrotome. These sections were subsequently stained with a combination of 2% uranyl acetate and lead citrate. Microscopic observation of the specimen was conducted using a HT7700 transmission electron microscope, operated at 80 kV on a Hitachi TEM system control from Japan.
2.6. Analyses of 16S–23S Internal Transcribed Spacer (ITS)
The tRNA gene sequences were detected using the tRNAscan SE 2.0 web server [31] and compared with related taxa by analyzing the 16S-23S rRNA ITS sequences of cyanobacterial strains. To simulate the secondary structure of conserved regions, namely D1-D1′, Box-B, and V3 helices, RNAfold webserver was used under standard conditions [32].
3. Results
3.1. Morphological Description
Pegethrix qiandaoensis K. Gao, Y. Cheng et R. Li sp. nov. (Figure 1).

Figure 1.
Light microscopy of Pegethrix qiandaoensis strains. A single trichome with a sheath (a,h,i). Morphological photos confirm that the strains that were studied have no obvious nodules cells and no knots have been observed in the filaments (a–i). Observed obvious cell wall structure (e,g,i). Macro-culture status of Pegethrix qiandaoensis strains (j). Scale bars: 10 µm.
The colony appears bright blue-green or olive-green. The filaments are long and vary in width between young and mature trichomes. Morphological images confirmed that no obvious nodule cells or knots were seen in the filaments of the studied strains. Occasionally, the filaments may have single or double false branches, and, in rare cases, the strains show several separate trichomes (Figure 1c). The width of the trichomes is typically between 2.3 and 4.0 µm. The sheath is firm and usually attached to the trichome, although it may occasionally widen or exhibit irregular and stratified patterns (Figure 1h,i). The sheath is absent in immature filaments or hormogonia. The trichomes themselves are untapered, with no or slight constriction at distinct cross walls (Figure 1e,i), and do not form nodules. This trait sets it apart from other Pegethrix species. The hormogonia are short (Figure 1g). Cells are usually isodiametric, often shorter than wide, exclusively in meristematic zones and slightly longer than wide in young trichomes. The length of the cells ranges from (1.3)–1.7 to 2.7 µm. End cells were rounded. The thylakoids are parietally arranged (four or five per cell) (Figure 2).
Reference strains: WZU0719 and WZU0723 (The Algae Culture Collection of the College of Life and Environmental Science, Wenzhou University, China).
Type locality: Isolated from the water surface of Qiandao Lake in Zhejiang Province, China (118°34′ E, 29°22′ N).
Holotype designated here: Specimen No. WZUH-ZLYN202312, containing dry material of the strain Pegethrix qiandaoensis WZU 0719, has been preserved in the Herbarium of Wenzhou University (WZUH) located in Wenzhou, Zhejiang Province, China.
Etymology: qiandaoensis refers to the Qiandao lake where the strain was isolated, transliterated into Latin.
Habitat: Free-living in water surface.
3.2. Molecular and Phylogenetic Analyses
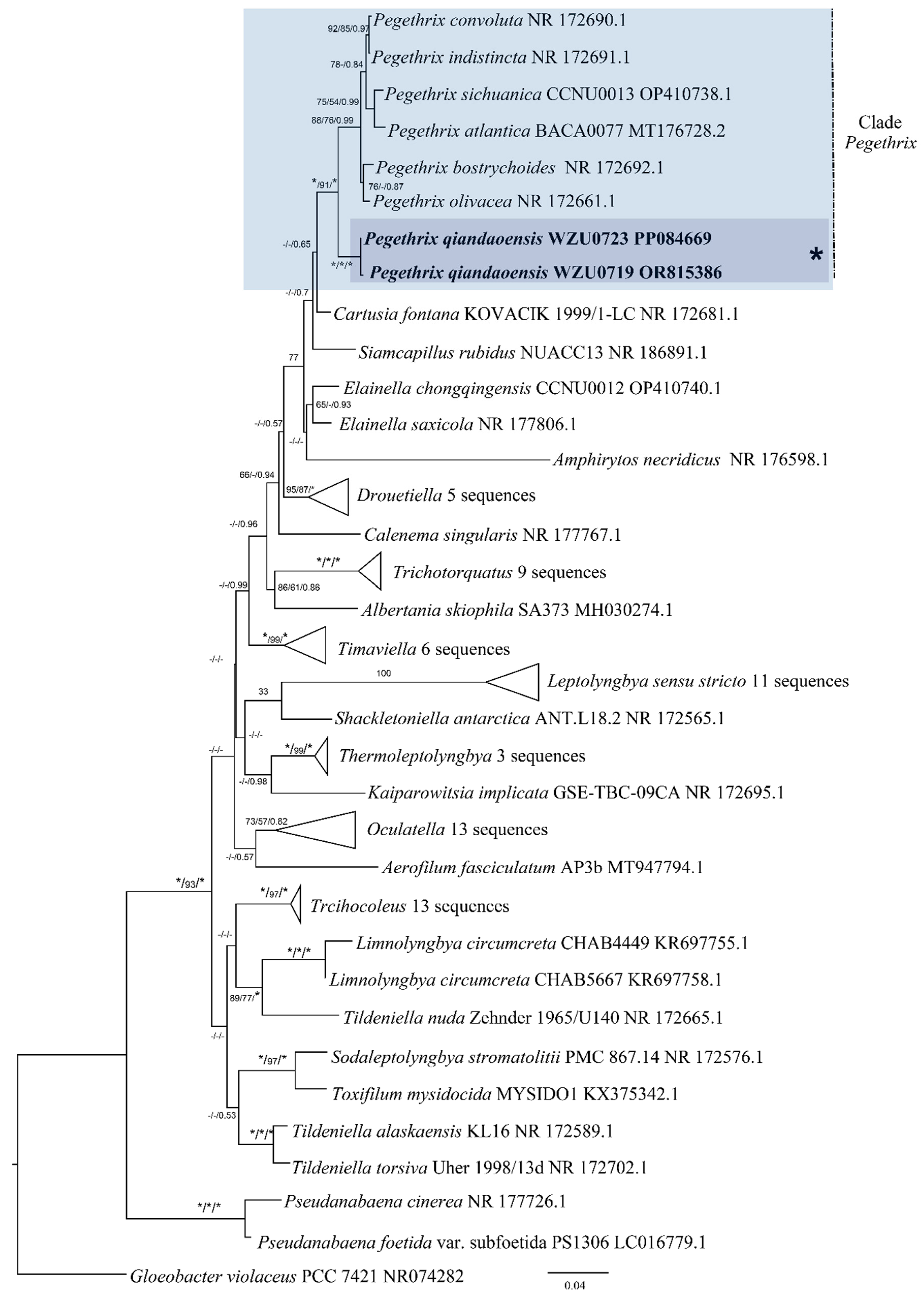
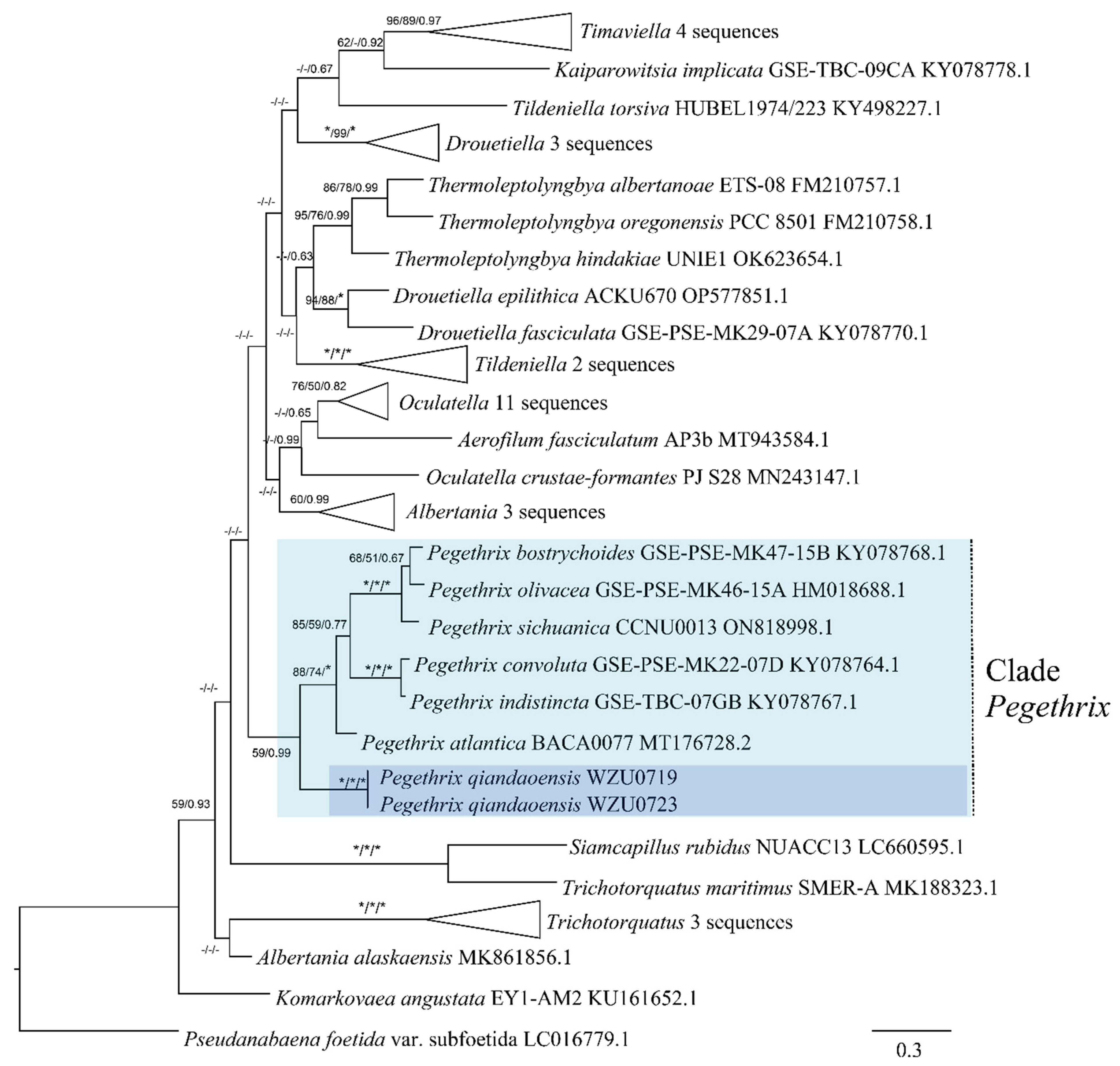
Based on the 16S rRNA gene sequences, a distance matrix revealed that the Pegethrix qiandaoensis strains had a resemblance of 96.73–97.36% to other Pegethrix species (Table 1). A total of 79 representative taxa sequences were examined in the phylogenetic analysis to evaluate the placement of the Pegethrix clade among the cyanobacteria (Figure 3). The results of phylogenetic analyses, including MP, ML, and BI analyses, exhibited consistent tree topologies. The phylogeny constructed using the 16S rRNA gene clearly demonstrates that our strain resided within the genus Pegethrix (MP and ML bootstrap percentages of 100% and 100% (BP) and posterior probability (PP) 1.00). The Pegethrix qiandaoensis clade occupies an independent phylogenetic position in Pegethrix. For a separate ITS phylogenetic tree (Figure 4), the Pegethrix qiandaoensis clade can also occupy an independent phylogenetic site within the genus. The differentiation of it has been widely used as molecular evidence of species differentiation at the genus level.

Table 1.
Comparison of the 16S rRNA gene sequence similarity between Pegethrix qiandaoensis and closely related species. Similarity = [1 − (p-distance)] × 100.
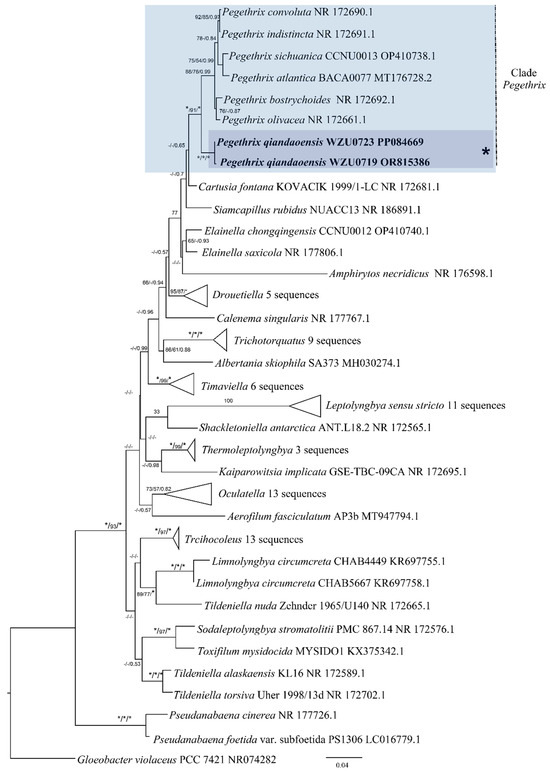
Figure 3.
Maximum-likelihood (ML) phylogenetic tree of Pegethrix qiandaoensis strains based on 16S rRNA gene sequences. For MP/ML methods and Bayesian posterior probabilities, bootstrap values above 50% are displayed in the BI tree. * Indicates a bootstrap value of 100 and a posterior probability of 1.00 for MP, ML, and BI. The novel filamentous strains of this study are indicated in bold.
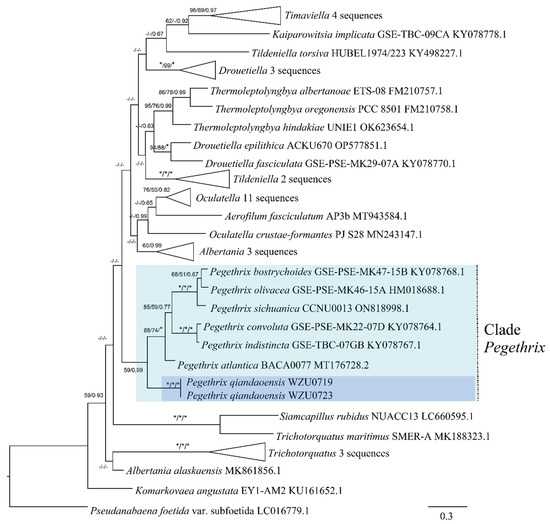
Figure 4.
Bayesian inference (BI) phylogenetic tree of Pegethrix qiandaoensis strains based on ITS rRNA gene sequences. For MP/ML methods and Bayesian posterior probabilities, bootstrap values above 50% are displayed in the BI tree. * Indicates a bootstrap value of 100 and a posterior probability of 1.00 for MP, ML, and BI. The novel filamentous strains of this study are indicated in bold.
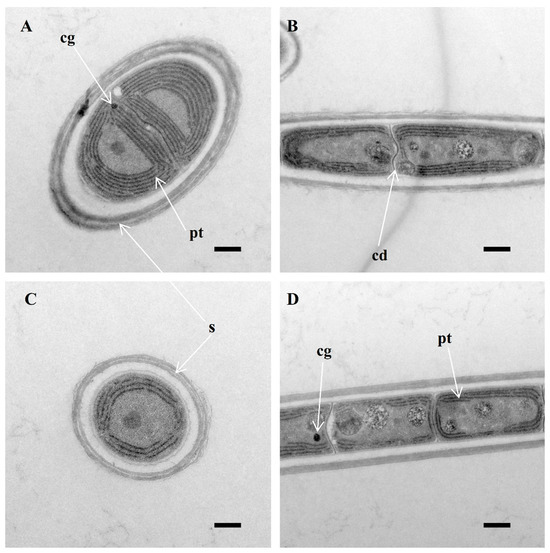
Figure 2.
Longitudinal section of vegetative cell (TEM), showing sheath (s) and parietal thylakoids (pt) (A,C,D), cyanophycin granules (cg) (A,D) and cell division—formation of new cell wall (cd) (B), and a lack of cell constriction. Scale bar: 0.5 μm.
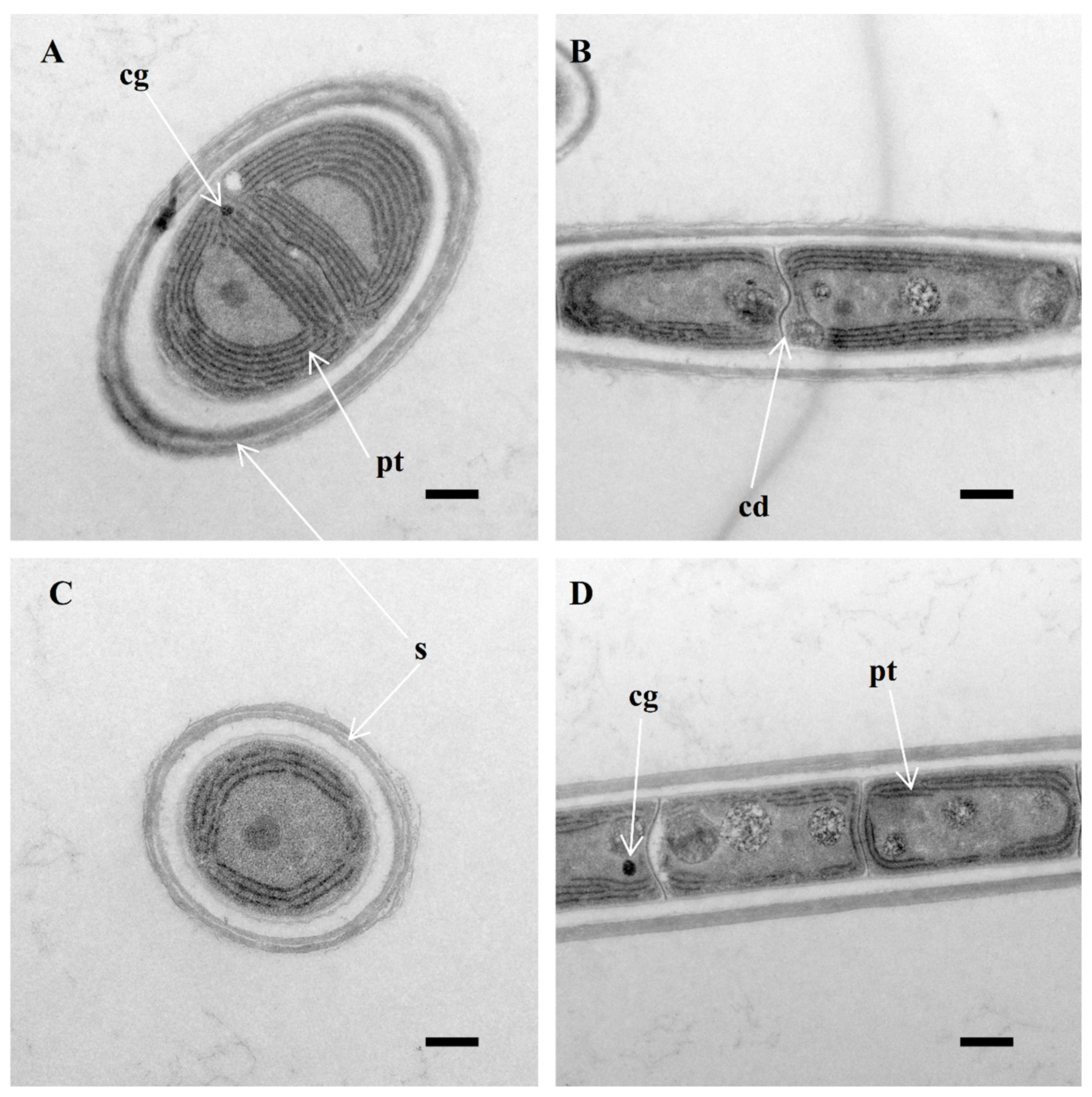
Figure 2.
Longitudinal section of vegetative cell (TEM), showing sheath (s) and parietal thylakoids (pt) (A,C,D), cyanophycin granules (cg) (A,D) and cell division—formation of new cell wall (cd) (B), and a lack of cell constriction. Scale bar: 0.5 μm.
3.3. Molecular Analyses on the ITS between 16S–23S rRNA Genes
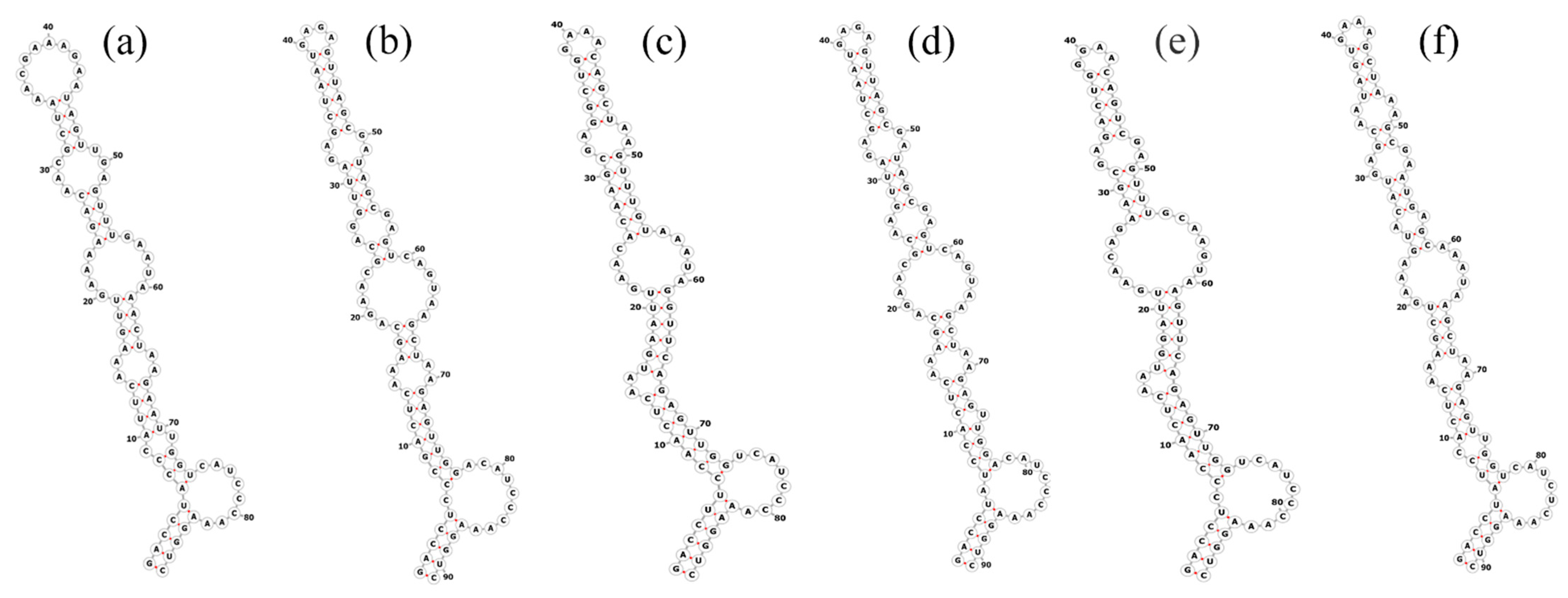
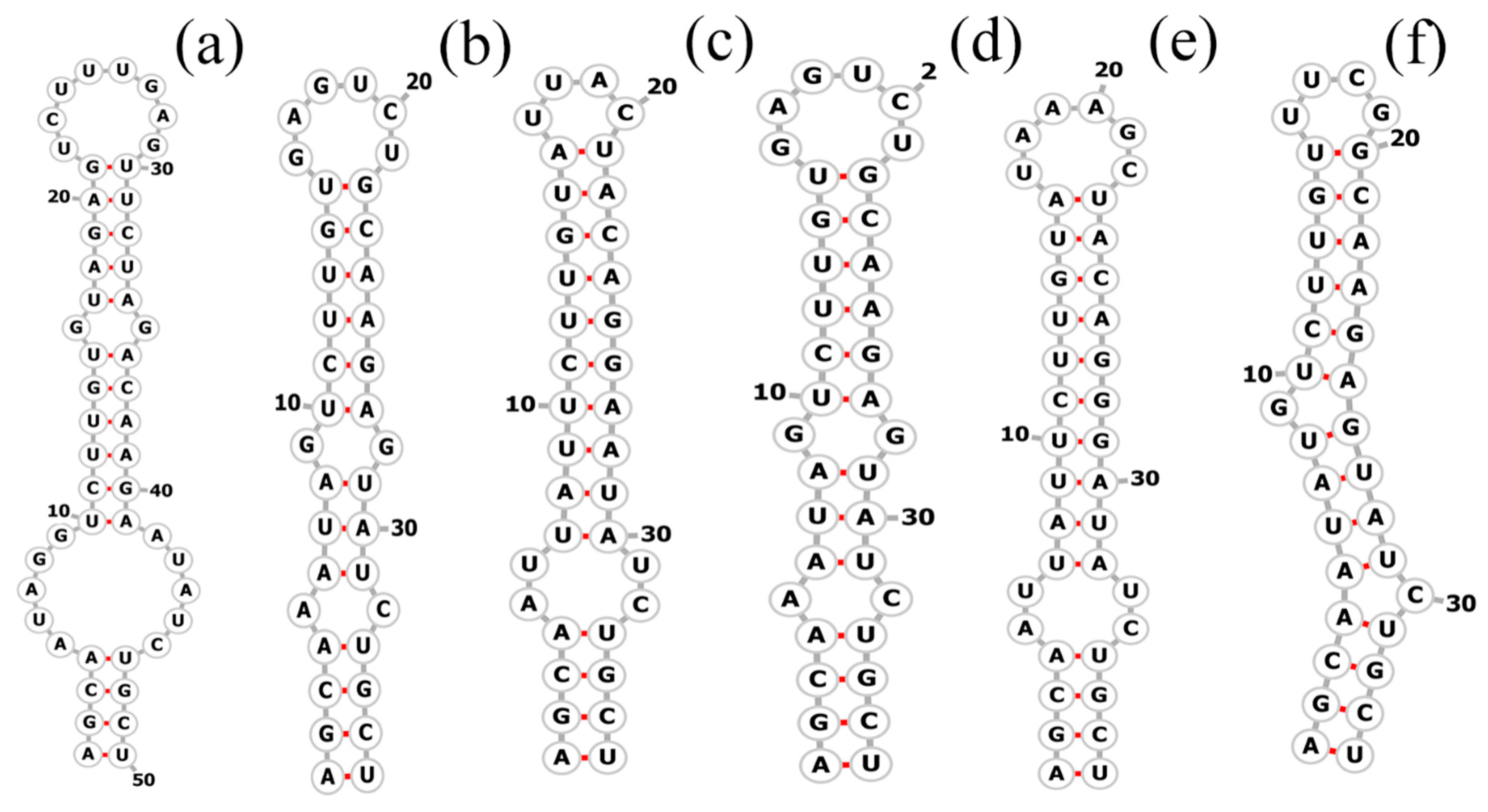
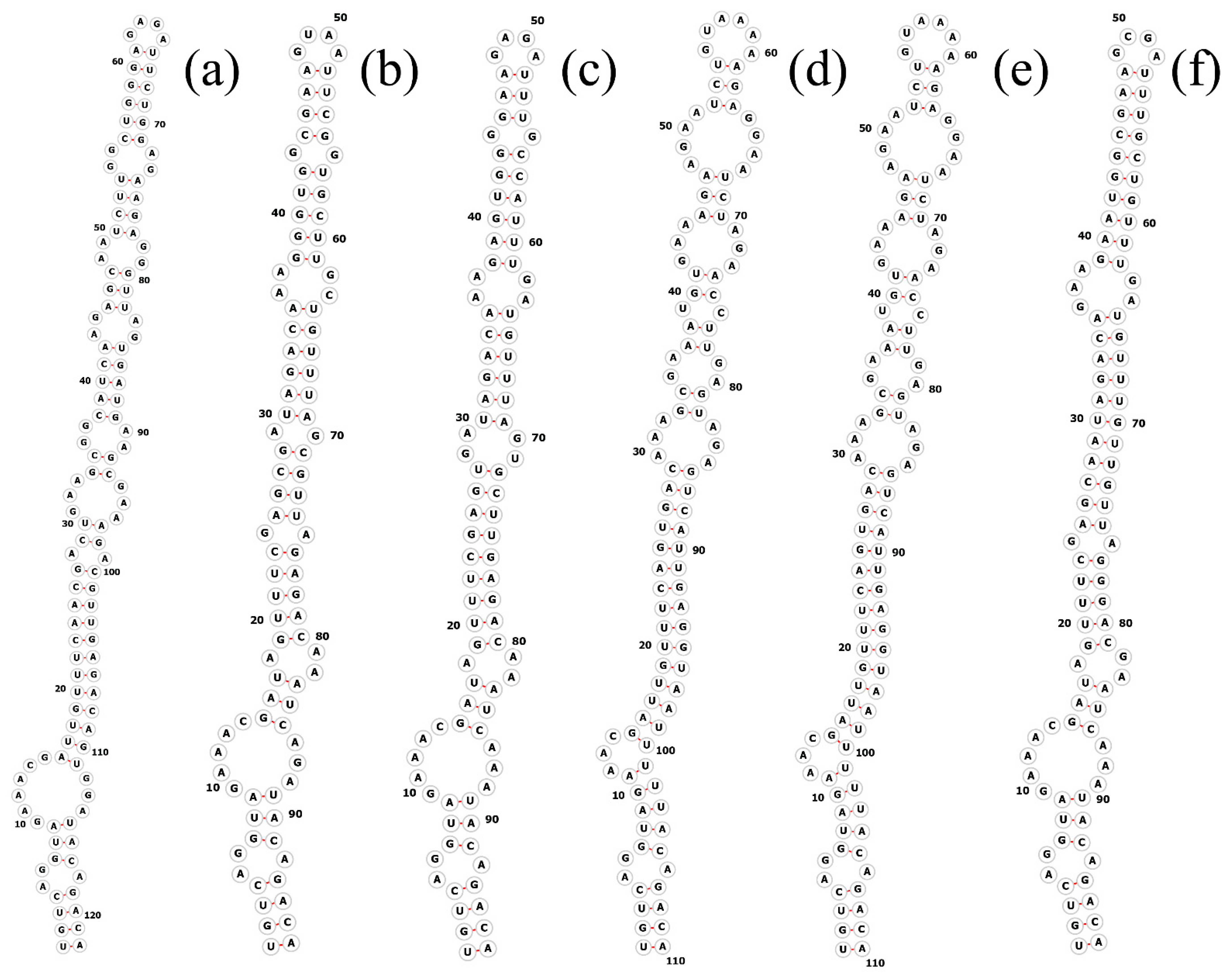
The total length of the Pegethrix qiandanensis strains ITS clones obtained in this study was 649 bp (Table 2), and all sequences contained tRNA Ile and tRNA Ala. The 16S–23S rRNA ITS secondary structures show some similarity among species, even for the D1–D1′ helix (Figure 5) and Box-B helix (Figure 6), with the highest variability found in the V3 region (Figure 7). Pegethrix qiandaoensis has the largest top ring structure (5′-AACGAAUAA-3′). P. atlantica and P. bostrychoides exhibit similar basolateral bulges on the D1–D1′ helix (Figure 5), a feature unique to these species, although there are some differences in the central inner loop. The terminal hairpin of Pegethrix contains four-nucleotide residues (5′-GGAA-3′) found in P. convoluta, P. atlantica, and P. bostrychoides, and the same nucleotide residue is found in P. convoluta an P. indistincta (5′-GAGA-3′). P. sichuanica has a unique nucleotide residue (5′-GGAA-3′).
Table 2.
Analysis of the length (number of nucleotides) of the 16S-23S ITS region of Pegethrix strains.
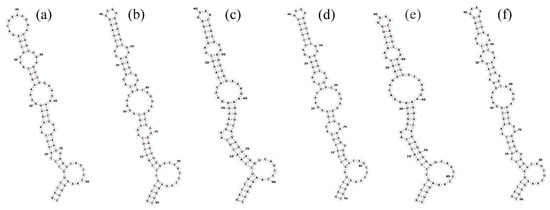
Figure 5.
Secondary structures of the D1–D1′ helix in Pegethrix species. (a–f) D1–D1′ helices: (a) Pegethrix qiandaoensis WZU0719. (b) Pegethrix convoluta. (c) Pegethrix bostrychoides. (d) Pegethrix indistincta. (e) Pegethrix sichuanica. (f) Pegethrix atlantica.
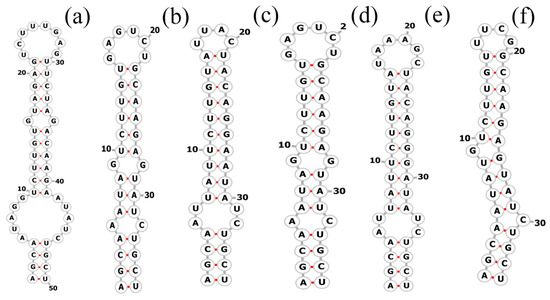
Figure 6.
Secondary structures of the Box–B helix in Pegethrix species. (a–f) D1–D1′ helices: (a) Pegethrix qiandaoensis WZU0719. (b) Pegethrix convoluta. (c) Pegethrix bostrychoides. (d) Pegethrix indistincta. (e) Pegethrix sichuanica. (f) Pegethrix atlantica.
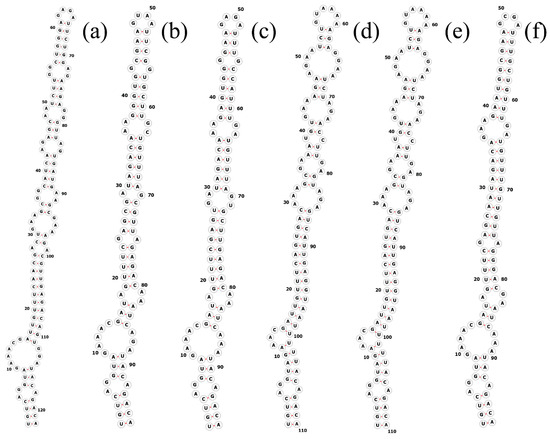
Figure 7.
Secondary structures of the V3 helix in Pegethrix species. (a–f) D1–D1′ helices: (a) Pegethrix qiandaoensis WZU0719. (b) Pegethrix convoluta. (c) Pegethrix bostrychoides. (d) Pegethrix indistincta. (e) Pegethrix sichuanica. (f) Pegethrix atlantica.
4. Discussion
To establish the cyanobacterial taxonomic system consisting of monophyletic groups at all ranks is becoming an ideal aim for studying the true diversity of cyanobacteria [33,34]. Further, with the usage of the polyphasic approach, a large number of cyanobacterial new genera and species has been described, which subsequently led to substantial taxonomic revisions, especially at the order and family levels [4,14,16]. In particular, thin filamentous cyanobacteria, previously only belonging to Oscillatoriales, have been divided into several orders in the updated taxonomic system [16], including Nodosilineales, Oculatellales and Leptolyngbyales. In this study, we present a scientific investigation that focuses on examining and categorizing two strains of thin filamentous cyanobacteria through the polyphasic approach. Based on a comprehensive analysis, we have identified them within the genus Pegethrix, Oculatellaceae, and further established them as a novel species. This conclusion is reached by comprehensively considering the sequence threshold of the 16S rRNA gene, the phylogeny based on the 16S rRNA gene, and the secondary structure and morphological characteristics of ITS [35,36]. The results of this study also revealed that species with similar phylogenetic positions can exhibit significant differences in habitat performance.
Since its establishment in 2018, the genus Pegethrix has been shown to exhibit remarkable similarities among species. Because of the uncomplicated morphological traits within this genus, some overlapping characteristics exist among the species. Nonetheless, specific morphological attributes provide valuable diagnostic clues for identifying Pegethrix qiandaoensis in this study. These attributes comprise the development of elongated, loosely coiled filaments under typical lighting conditions, as well as the presence of a stratified and expanded sheath. Moreover, the filaments in P. qiandaoensis hardly knot to form nodules, revealing the obvious difference from other Pegethrix species. Phylogenetically, the close clade formed by the strains of P. qiandaoensis and the other six Pegethrix species based on the 16S rRNA gene, with high support values, represents the strong monophyletic group for the genus Pegethrix (Figure 4). Moreover, 96.73–97.36% of the 16S rRNA similarities among P. qiandaoensis and the other Pegethrix species, lower than all the proposed cut-off values of bacterial species separation [37,38], strongly support the establishment of Pegethrix qiandaoensis sp. nov. From the initial establishment of the genus Pegethrix described in Mai et al. (2018) [17], the sequence identities of the 16S rRNA gene sequences among the six species of the genus, ranging from 98.6 to 99.9%, did not provide evidence of species separation in this genus using only these sequence identities. However, the addition of Pegethrix qiandaoensis sp. nov. in this study can help considerably to molecularly provide a high validity of the genus Pegethrix. The 16S–23S ITS region plays a crucial role in distinguishing between different species [8,39]. The results from ITS analyses indicate that each species has distinct secondary structures in the 16S-23S ITS region, specifically in the D1-D1’ and Box-B helices. These structural differences, including the significantly different Box-B helix observed in P. qiandaoensis compared to the other Pegethrix species (Figure 6 and Figure 7), provide further evidence of significant genetic divergence among various lineages [8,40,41]. This observation holds true for all the genera examined and discussed in this research, highlighting the consistent reliability and sensitivity of the ITS gene in distinguishing between genera.
Last, from the ecological viewpoint, the occurrence of this species in a freshwater ecosystem in China further distinguished it from previously documented species, which primarily inhabit soil, stones, rocks, and thermal springs. This study also revealed that species with a similar phylogenetic development relationship can exhibit significant differences in habitat performance. Apparently, P. qiandaoensis in this study was shown to stand as the most distinctive member in the genus Pegethrix.
In conclusion, the present study provided a typical case with the focus on diversity of a cyanobacterial genus with thin filaments, based on the polyphasic approach. In recent years, a rapidly growing number of new taxa of the thin filamentous cyanobacteria have been intensively reported and progress in the taxonomy of such cyanobacterial groups has led to considerable revisions in cyanobacterial taxonomy. Although only one species new to science was recognized in this study from a genus of the thin filamentous cyanobacteria, it is clear that more species, genera, or high categories in these cyanobacterial forms remain to be named and defined. With enhancement of the polyphasic culture-dependent approach, more results on the diversity of thin filamentous cyanobacteria from less explored regions in China will be highly expected.
Author Contributions
Conceptualization, K.G. and Y.C.; methodology, Y.C.; software, Y.C. and P.X.; formal analysis, Y.C. and R.G.; investigation, K.G. and Y.C.; resources, Z.W. and H.Z.; data curation, Y.C.; writing—original draft preparation, K.G. and Y.C.; writing—review and editing, F.C. and R.L.; visualization, F.C.; funding acquisition, R.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study is funded by the Zhejiang Provincial Natural Science Foundation of China (LD21C030001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The 16S rRNA gene sequences are available in the GenBank database.
Acknowledgments
The first author thanks Huimin Fang and Yuping Fan for physical assistance in the process of collecting samples, and thanks Zhenfei Xing for technical assistance with the TEM images.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Whitton, B.A. The Ecology of Cyanobacteria II: Their Diversity in Time and Space; Springer: Dordrecht, The Netherlands, 2012; 669p. [Google Scholar]
- Paerl, H.W.; Huisman, J. Climate—Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef]
- Anagnostidis, K.; Komárek, J. Modern approach to the classification system of the Cyanophytes 3: Oscillatoriales. Algol. Stud. 1988, 50, 327–472. [Google Scholar]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphyletic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Kaštovský, J.; Gomez, E.B.; Hladil, J.; Johansen, J.R. Cyanocohniella calida gen. et sp. nov. (Cyanobacteria: Aphanizomenonaceae) a new cyanobacterium from the thermal springs from Karlovy Vary, Czech Republic. Phytotaxa 2014, 181, 279–292. [Google Scholar] [CrossRef]
- Casamatta, D.A.; Johansen, J.R.; Vis, M.L.; Broadwater, S.T. Molecular and morphological characterization of ten polar and near-polar strains within the Oscillatoriales (Cyanobacteria). J. Phycol. 2005, 41, 421–438. [Google Scholar] [CrossRef]
- Hašler, P.; Dvořák, P.; Poulíčková, A.; Casamatta, D.A. A novel genus Ammassolinea gen. nov. (Cyanobacteria) isolated from sub–tropical epipelic habitats. Fottea 2014, 14, 241–248. [Google Scholar] [CrossRef]
- Osorio-Santos, K.; Pietrasiak, N.; Bohunicka, M.; Miscoe, L.; Kovacik, L.; Martin, M.P.; Johansen, J.R. Seven new species of Oculatella (Pseudanabaenales, Cyanobacteria): Taxonomically recognizing cryptic diversification. Eur. J. Phycol. 2014, 49, 450–470. [Google Scholar] [CrossRef]
- Dvořák, P.; Poulíčková, A.; Hašler, P.; Belli, M.; Casamatta, D.A.; Papini, A. Species concepts and speciation factors in cyanobacteria, with connection to the problems of diversity and classification. Biodivers. Conserv. 2015, 24, 739–757. [Google Scholar] [CrossRef]
- Genuário, D.B.; Vaz, M.G.M.V.; Hentschke, G.S.; Sant’anna, C.L.; Fiore, M.F. Halotia gen. nov., a phylogenetically and physiologically coherent cyanobacterial genus isolated from marine coastal environments. Int. J. Syst. Evol. Microbiol. 2015, 15, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Hauerová, R.; Hauer, T.; Kaštovský, J.; Komárek, J.; Lepšová-Skácelová, O.; Mareš, J. Tenebriella gen. nov.—The dark twin of Oscillatoria. Mol. Phylogenet. Evol. 2021, 165, 107293. [Google Scholar] [CrossRef] [PubMed]
- Johansen, J.R.; Olsen, C.E.; Lowe, R.L.; Fučíková, K.; Casamatta, D.A. Leptolyngbya species from selected seep walls in the Great Smoky Mountains National Park. Algol. Stud. 2008, 126, 21–36. [Google Scholar] [CrossRef]
- Turicchia, S.; Ventura, S.; Komárková, J.; Komárek, J. Taxonomic evaluation of the cyanobacterial microflora from alkaline marshes of northern Belize. 2. Diversity of oscillatorialean genera. Nova Hedwig. 2009, 89, 165–200. [Google Scholar] [CrossRef]
- Komárek, J. Several problems of the polyphasic approach in the modern cyanobacterial system. Hydrobiologia 2018, 811, 7–17. [Google Scholar] [CrossRef]
- Strunecký, O.; Raabova, L.; Bernardova, A.; Ivanova, A.P.; Semanova, A.; Crossley, J.; Kaftan, D. Diversity of cyanobacteria at the Alaska North Slope with description of two new genera: Gibliniella and Shackletoniella. FEMS Microbiol. Ecol. 2019, 96, fiz189. [Google Scholar] [CrossRef]
- Strunecký, O.; Ivanova, A.P.; Mareš, J. An updated classification of cyanobacterial orders and families based on phylogenomic and polyphasic analysis. J. Phycol. 2022, 59, 12–51. [Google Scholar] [CrossRef]
- Mai, T.; Johansen, J.R.; Pietrasiak, N.; Bohunická, M.; Martin, M.P. Revision of the Synechococcales (Cyanobacteria) through recognition of four families including Oculatellaceae fam. nov. and Trichocoleaceae fam. nov. and six new genera containing 14 species. Phytotaxa 2018, 365, 1–59. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, Z.; Huang, L.; Zhang, L.; Yu, G.; Chen, M.; Li, R.; Qiu, B. Chlorophyll f production in two new subaerial cyanobacteria of the family Oculatellaceae. J. Phycol. 2023, 59, 370–382. [Google Scholar] [CrossRef]
- Luz, R.; Cordeiro, R.; Kaštovský, J.; Johansen, J.R.; Dias, E.; Fonseca, A.; Urbatzka, R.; Vasconcelos, V.; Gonçalves, V. New terrestrial cyanobacteria from the Azores Islands: Description of Venetifunis gen. nov. and new species of Albertania, Kovacikia and Pegethrix. Phycologia 2023, 62, 483–498. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen–Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Goodman, A.E. Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl. Environ. Microbiol. 1995, 61, 3875–3883. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef]
- Gkelis, S.; Rajaniemi, P.; Vardaka, E.; Moustaka–Gouni, M.; Lanaras, T.; Sivonen, K. Limnothrix redekei (Van Goor) Meffert (Cyanobacteria) strains from Lake Kastoria, Greece from a separate phylogenetic group. Microb. Ecol. 2005, 49, 176–182. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Page, R.D.M. TreeView: An application to display phylogenetic trees on personal computers. Bioinformatics 1996, 12, 357–358. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Mathews, D.H.; Disney, M.D.; Childs, J.L.; Schroeder, S.J.; Zuker, M.; Turner, D.H. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Nucleic Acids Res. 2004, 101, 7287–7292. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Geng, R.; Shan, L.; Liu, Y.; Zhang, H.; Xiao, P.; Li, R. Taxonomic Discussion on Cyanobacterial Systematics at Family Level, with Special Regards to Phormidiaceae by Using the Strains of Chinese Newly Recorded Genera Ancylothrix and Potamolinea. Diversity 2022, 14, 301. [Google Scholar] [CrossRef]
- Cai, F.; Li, S.; Zhang, H.; Yu, G.; Li, R. Nodosilinea hunanesis sp. nov. (Prochlorotrichaceae, Synechococcales) from a Freshwater Pond in China Based on a Polyphasic Approach. Diversity 2022, 14, 364. [Google Scholar] [CrossRef]
- Perkerson, R.B.; Johansen, J.R.; Kovacik, L.; Brand, J.; Kastovsky, J.; Casamatta, D.A. A unique pseudanabaenalean (Cyanobacteria) genus Nodosilinea gen. nov. based on morphological and molecular data. J. Phycol. 2011, 47, 1397–1412. [Google Scholar] [CrossRef]
- Sciuto, K.; Moro, I. Detection of the new cosmopolitan genus Thermoleptolyngbya (Cyanobacteria, Leptolyngbyaceae) using the 16S rRNA gene and 16S–23S ITS region. Mol. Phylogenet. Evol. 2016, 105, 15–35. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 4, 6–9. [Google Scholar]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Erwin, P.M.; Thacker, R.W. Cryptic diversity of the symbiotic cyanobacterium Synechococcus spongiarum among sponge hosts. Mol. Ecol. 2008, 17, 2937–2947. [Google Scholar] [CrossRef]
- Strunecký, O.; Bohunická, M.; Johansen, J.R.; Čapková, K.; Raabová, L.; Dvořák, P.; Komárek, J. A revision of the genus Geitlerinema and a description of the genus Anagnostidinema gen. nov. (Oscillatoriophycidae, Cyanobacteria). Fottea 2017, 17, 114–126. [Google Scholar] [CrossRef]
- Rasouli–Dogaheh, S.; Noroozi, M.; Khansha, J.; Amoozegar, M.A.; Rahimi, R.; Nikou, M.M.; Hauer, T. Khargia gen. nov., a new genus of simple trichal Cyanobacteria from the Persian Gulf. Fottea 2023, 23, 49–61. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).