Abstract
We describe efforts to reintroduce the extinct-in-the-wild Spix’s Macaw (Cyanopsitta spixii) within its historical range in eastern Brazil. Twenty captive-reared Spix’s Macaws were released, along with 15 Blue-Winged Macaws (Primolius maracana), as heterospecific flocks in two events during the dry and rainy seasons of 2022. We monitored the release groups through daily observation and telemetry tracking from early June 2022 to early June 2023. We documented an overall first-year survival of 58.3% (CI: 37.8–78.7%), with 65% of Spix’s Macaws establishing a stable area of activity within 5 km of the release site, excluding any temporary long-distance forays. Eighty-five percent of released Spix’s Macaws exhibited flock cohesion, including interactions with and integration into wild Blue-Winged Macaw groups at the release site. Several released Spix’s Macaws formed pair-bonds with conspecifics and engaged in nest cavity exploration, breeding behavior, and territorial defense of nest sites, with three females also laying and incubating eggs. One nesting pair successfully hatched and reared chicks in an artificial nest cavity. These releases employed a novel reintroduction strategy using a surrogate species model, the sympatric Blue-Winged Macaw, to “mentor” and facilitate post-release adaptation by the target species and increase the total number of individuals in the release cohort. Lastly, participatory monitoring by local citizens is considered a way to engage and involve local communities in species and habitat conservation and potentially create new employment opportunities in the region.
1. Introduction
Over the past few centuries, there has been an exceptionally rapid loss of biodiversity, often termed the sixth mass extinction [1,2], primarily attributed to anthropogenic activities and global climate change. This scenario can adversely affect forest-dependent species such as many Psittaciformes (hereafter called parrots) and represents a significant threat to their survival and population persistence [3]. Globally, this avian group is highly at risk, with nearly a third (29%) of these species threatened with extinction [4], mainly as a result of anthropogenic activities [4,5].
The extinct-in-the-wild Spix’s Macaw, Cyanopsitta spixii, is an example of a parrot species that became extinct before effective conservation measures could be implemented [6,7]. Although the species was discovered more than 200 years ago in the region of Juazeiro, northeastern Brazil [8,9], the accurate location of contemporary populations was only confirmed in 1986, when the last three wild individuals were found in the municipality of Curaçá, State of Bahia [10,11]. The species is endemic to a restricted semiarid area of a unique dry forest domain in Brazil called the “Caatinga”. Little is known about the ecology and behavior of Spix’s Macaw in the wild, as existing data come from observations of the last specimens in the wild and anecdotal information from people residing within its historical range [10,11,12,13,14,15]. Spix’s Macaw appears to have been dependent on riparian gallery forests dominated by the Caribbean trumpet tree (Tabebuia aurea) bordering seasonal streams in the states of Bahia and Pernambuco [13,15]. Anecdotal accounts also suggest that Spix’s Macaw lived in small flocks [14]. Their dietary preference includes “faveleira” (Cnidoscolus quercifolius) and fruits of Jatropha mollissima, as well as other seasonally available food items [15]. The macaws nested in natural cavities and those created by the Crimson-Crested Woodpecker (Campephilus melanoleucos), primarily in Tabebuia trees of the riparian forests, and occurred during the rainy season, which ranges from December to April [15]. Within the historical range of Spix’s Macaw, five other parrot species are sympatric [16]. Among these, the Blue-Winged Macaw (Primolius maracana) is the most similar in terms of ecology and biology to Spix’s Macaw as it shares the same habitat, nesting cavities and food items. In addition, the last male of Spix’s Macaw observed in the wild from 1990 to 2000 formed an apparent pair bond with a Blue-Winged Macaw [17,18]. These characteristics make the Blue-Winged Macaw an appropriate model species to assist in the reintroduction of Spix’s Macaw [15].
Due to its inherent beauty and rarity, Spix’s Macaw was a victim of both Brazilian and international pet collectors, even before its geographical range was accurately delimited. Captures of wild birds for the pet trade were first reported in the early 20th century and intensified during the 1960s through to the 1980s, leading to a precipitous population decline [14]. Furthermore, the expansion of agriculture and livestock over the past three centuries in the region [13] led to substantial habitat loss and degradation. These factors together led to the eventual extinction of the species in the wild, where it was last recorded in 2000 [7]. Accordingly, the reintroduction of captive-reared individuals has become the only viable option for Spix’s Macaw conservation [19,20,21] and re-establishing the species in the wild [22]. To achieve this goal, the Brazilian government, along with national and international non-governmental organizations (NGOs), developed strategic processes and actions for species conservation over the last two decades [15,20,21]. The resulting National Action Plan (NAP) was based on thorough review of the best available information, including well-defined and achievable goals, and incorporated stakeholder perspectives [21] and agreement among those involved, as indicated by IUCN Species Survival Commission (SSC). Measurable improvements in the ex situ population status were achieved with the substantial upscaling of a collaboratively conceived species conservation plan [20]. After stable ex situ reproduction and growing populations were achieved, the reintroduction plan—one of the most challenging conservation efforts of Brazil—was implemented.
The reintroduction process typically includes steps such as the selection of an appropriate release site, intensive pre-release training and preparation of release candidates, release of sufficient numbers of individuals, and allocation of adequate resources for monitoring results [22,23]. As part of the overall strategy for species recovery, two protected areas (i.e., Spix’s Macaw Refuge and Spix’s Macaw Environmental Protection Area) were established within the species’ historic occurrence area in northern Bahia state to enable the protection and recovery of the habitat and integration of local communities with the species’ conservation [21,24]. The refuge was formed from private lands, where a breeding and reintroduction facility was built in 2020 to begin the ongoing in situ conservation project [25,26]. Therefore, the long-term goal of the reintroduction of Spix’s Macaw in its original area of occurrence was to promote a continuous population increase in the wild, combined with community involvement in sustainable practices for conserving native habitats and associated species [21].
The reintroduction of Spix’s Macaw and establishment of a viable population required designing and testing species-specific reintroduction techniques. There was only one previous attempt to reintroduce a wild-hatched female in 1995 to mate with the last male, which was unsuccessful [15]. Therefore, we conducted initial pilot releases in 2022 to achieve the reintroduction and restoration of the species in an appropriate location, reduce the risk of global extinction and increase the species’ local and regional importance as a symbol of conservation. Due to ecological similarities [15], sympatric Blue-Winged Macaws were used as a surrogate species to refine Spix’s Macaw release methodologies. Importantly, because the Blue-Winged Macaws to be released were all “wild-sourced” (i.e., captured as wild adults or nestlings by project personnel), rather than captive-bred, they, therefore, had not been subject to the potential erosion of adaptive behaviors that can accrue from generations of captive-breeding [27,28,29] or the potential maintenance of maladaptive traits in captive populations [29,30,31]. Accordingly, our overarching objective was to use the innate “biological culture” of the wild Blue-Winged Macaws to increase the likelihood of the success of Spix’s Macaw’s reintroduction via close association of the target species with a “mentor” species as part of a heterospecific group. The survival advantages that may accrue to individual species when part of heterospecific groups are well documented across a variety of taxa [32,33,34,35,36,37,38,39]. Moreover, augmenting the sizes of the release groups via the inclusion of Blue-Winged Macaws also allows for a potential decrease in overall per capita predation risk, without a concomitant increase in the total numbers of Spix’s Macaws released. Thus, this methodology was also designed to reduce or mitigate potential post-release Allee effects [40,41,42,43]. Importantly, to prevent the potential formation of heterospecific pair bonds [18], each species was reared separately and only combined as heterospecific release groups during the final months of pre-release training.
For the pilot releases, we used 20 individuals of Spix’s Macaw along with 15 Blue-Winged Macaws. Our intermediate goals were as follows: (1) to integrate groups of Blue-Winged Macaws with Spix’s Macaws to prepare a cohesive heterospecific group; (2) conduct two release events of Spix’s Macaws with Blue-Winged Macaws; (3) carry out post-release monitoring of the reintroduced population; (4) quantify specific indicators of reintroduction success—cohesion of the release groups, fidelity to the reintroduction site, and survival over the periods of 3 months and 1 year post-release [43]; (5) use adaptive management to evaluate and refine techniques to establish the population in the region; and (6) engage and involve the local community and society in the conservation, study, and monitoring of the species and its habitat. Here, we describe the results of the first year of the reintroduction of Spix’s Macaw in its native habitat.
2. Materials and Methods
2.1. Reintroduction Site
We chose the most suitable area for the reintroduction of Spix’s Macaw following methodology of White et al. [19], near the area used by the last wild individuals in the 1990s [15] (Figure 1). This area, located within the Caatinga forest domain, was also the area chosen to establish an in situ captive breeding and release facility. Although White et al. [12] found no relationship between the success of psittacine reintroductions and whether parrots were released within or outside their historical range, all extant evidence suggests that Spix’s Macaw is a dry forest habitat specialist and closely associated with, and adapted to, the Caatinga domain. Numerous studies [44,45,46,47,48] have found that the successful establishment of avian species outside their native habitat is less likely for species that are habitat and dietary specialists or those with narrow or limited ecological niches.
Figure 1.
General location of the reintroduction of Spix’s Macaw in the Caatinga domain of eastern Brazil. Confirmed range based on Barros et al. [15]. Precise location of releases not depicted for security reasons.
2.2. Release Candidate Observation and Training
In 2020, 52 Spix’s Macaws were transferred to the captive breeding facility to (1) begin breeding within the species’ historical range, (2) enable a population to be re-established in the wild, and (3) promote fidelity to the release site, group cohesion, and reproduction after release [43]. Candidates selected for releases were later placed in the training and release facility, which included two different-sized central enclosures (Figure 2), with birds first placed in the smaller of these enclosures. Once the birds demonstrated controlled flying skills, they were transferred to the larger enclosure, where they could progressively and safely improve their flight skills. Observations were made daily to detect (1) flock social interactions, (2) aggressiveness, (3) pair formation (e.g., allopreening, allofeeding), (4) feeding proficiency, and (5) predator recognition. We based our observations on Marcuk et al. [25], which provides the first comprehensive description of the behavioral repertoire of Spix’s Macaw in captivity. No agonistic interactions were recorded during this period in the training and release enclosure.

Figure 2.
Training and release cage complex used to reintroduce captive-reared Spix’s Macaws in the Caatinga forest domain of eastern Brazil. The cage complex is located at the edge of an adjacent riparian gallery forest. Note a large U-shaped flight and release module. The person standing in the foreground of the complex provides a visual dimensional scale. Photo credit: ACTP.
After acclimatization in the smaller enclosures, the group was transferred to a large “U- shaped” flight and release module, with a height of 7 m, width of 4 m, cumulative length of 47 m, and volumetric flight space of approximately 1316 m3 (Figure 2). The objective of the release module was to safely allow for the complex flight movement of birds, provide training to exercise flight muscles and allow interaction between individuals to be released, as well as with other wild birds (competitors and predators). To develop and maximize the flight agility of the birds, native woody vegetation was planted and established within the enclosure. Natural perches were also strategically placed to facilitate flight training and provide natural fruits and seeds. The release module was also equipped with two suspended feeders next to the front exit doors. Outside of the release doors, two suspended feeders were also placed for supplementary post-release feeding. All feeders were raised and lowered using a system of pulleys and steel cables. Food in the release module was offered twice daily, in the same proportions as previously offered in the breeding center. Native fruits, flowers, and seeds were also provided as food sources [15] according to their availability, both in the feeding stations and on branches of the trees from which they were sourced for later feeding tree recognition. Visual contact between the macaws and their keepers was limited to the minimum necessary to provide food, count the numbers of individuals, and observe behavior.
2.3. Release Groups
Captive-reared Spix’s Macaws were selected for release based on criteria such as genetics, age, gender, absence of avian pathogens, social behavior, and general physical condition. To maximize the genetic diversity of the reintroduced population, we employed data from the species’ Studbook and genetic sequencing. Birds aged three to seven years old were selected for release, as they may more quickly learn new skills and show fewer maladaptive or stereotypical behaviors derived from prolonged captivity. This age range also encompasses the age (3–5 years) at which the species becomes reproductively mature. The Blue-Winged Macaws released comprised one locally captured wild adult male and 1–2-year-old juveniles collected as wild nestlings in the region. The minimum number of individuals for the release was based, in part, on the traditional releases of White et al. [43,49], as well as availability. To prepare for release, Spix’s Macaws and Blue-Winged Macaws were maintained separately in three flocking enclosures, located approximately 350 m from the actual release site, and acclimatized for at least 1.5 years. Subsequent release flock formation was carried out in the training and release enclosure beginning in February 2022. Flight training, which consisted of allowing birds to fly freely within the flocking enclosures, was continued in the release module (Figure 2) for a total of four months for the first release group. The first release group consisted of 8 Spix’s Macaws and 8 Blue-Winged Macaws (n = 16 birds), while the second group consisted of 12 Spix’s Macaws and 7 Blue-Winged Macaws (n = 19 birds). The total cohort released in 2022, thus, comprised 20 Spix’s Macaws and 15 Blue-Winged Macaws.
2.4. Health Protocol
All release candidates were submitted to clinical evaluation and tested for known or potential psittacine pathogens present in Brazil [50]. All the birds tested negative via PCR for aviadenovirus, avipoxvirus, bornavirus, circovirus, alpha-herpesvirus, polyomavirus, paramyxovirus, Chlamydia psittaci, Mycobacterium avium, M. intracelullare, Mycoplasma gallisepticum, M. sinavae, and Salmonella spp. All birds were also prophylactically deparasitized during the pre-release preparation.
2.5. Telemetry Instrumentation
Two days prior to the transfer to the training and release cage complex, all Spix’s Macaws and two Blue-Winged Macaws were equipped with a “dummy” radio collar to acclimate the birds to foraging and flying with an actual radio collar [51]. The dummies were replaced with functioning radio collars (Model SI-2C, Holohil Systems Ltd., Ontario, CA, USA) two days prior to release. The transmitters weighed 13 g, or ~4% of body mass [52,53]. Although some avian telemetry studies have successfully used solar-powered GPS or data logger transmitters, at the time of reintroduction, no suitable such units had been developed that would simultaneously meet the maximum weight requirement (<14 g) and be sufficiently robust for deployment on this species [54].
2.6. Releases
After acclimatization in the training and release enclosure and observation of behavior, the first release group was selected and placed in the U-shaped release module. Spix’s Macaws that remained in the release enclosure were kept in the central module, while the release module was kept open from early June to early October (four months), allowing previously released birds to return during the day or at dusk. After this period, the birds confined in the central modules were transferred to the release module, which was opened again in early December and remained open thereafter following the initial release.
A “Type B soft release” [43] was conducted with conspecifics maintained at the release site (within the breeding and release enclosures) that allow interaction between the free-living and captive populations. Post-release supplementary feeding [22,43] was also conducted. Food and water were offered twice daily in two feeders suspended in front of the enclosure. Two additional feeders were established in trees within the area between the release and breeding facilities.
The first release period occurred during June, at the onset of the dry season. This time was selected to promote flock cohesion and site fidelity, as the availability of natural food items tends to be lower, thus enhancing the tendency of released birds to return to the feeders near and within the enclosures. Additionally, this time is outside the breeding season and its associated territoriality, further increasing the probability of flock cohesion and release site fidelity. The second release event occurred in December, near the onset of the breeding season, to promote potential post-release reproduction and attendant enhanced long-term site fidelity [43].
2.7. Post-Release Monitoring
The radio-tagged macaws were radio tracked 7 days/week, using two 6-meter observation towers located 350 and 800 m from the release cage, as well as from various strategic ground points throughout the immediate area. The monitoring was carried out using R-1000 or R-5000 receivers (Communication Specialists Inc., Orange, CA, USA), handheld 2-element Yagi antennas (RA-23K VHF Antenna, Telonics Inc., Mesa, AZ, USA), GPS eTrex 30x (datum WGS84, decimal degree), Avenza® maps (Avenza Systems Inc., Toronto, ON, Canada), and a compass. Spix’s Macaws were sighted and counted, and signals were detected every day at morning feeding (approximately 6:30 am), afternoon feeding (approximately 12:30 pm), and late afternoon (approximately 4:30 pm). When a radio signal was not detected and the tagged bird was not sighted, we searched the area in expanding concentric circles until contact was made or the signal was detected. During visual sightings, macaw behaviors and activities (e.g., foraging, preening, resting, etc.) were recorded.
2.8. Habitat Management
To facilitate nesting and promote associated release site fidelity, we installed 12 nest boxes [43,55] in Caribbean trumpet trees near the release site. We also installed zinc bands on the trunks of nest trees and suppressed the surrounding vegetation to avoid potential nest predation by scansorial predators [55,56]. We also removed Africanized bees (Apis mellifera) and Trigona aff. fuscipennis from four Caribbean trumpet trees adjacent the release area to further increase the availability of potential nest cavities.
2.9. Data Analyses
We estimated post-release survival using the Kaplan–Meier product-limit estimator [57,58] and reported weekly and cumulative survival and 95% confidence intervals at 3 months and 1 year post-release. This analytical method is appropriate when censored observations (unknown fates) are present, cohorts are released at different times (staggered entry), in which proportional survival (e.g., by age, sex) is not differentiated [57,58]. We assumed no age-specific differences in survival among the released macaws. As the species—like most psittacines—exhibits strong conspecific attraction, we likely violated the model assumption of the independence of individual survival probabilities. However, this would affect only precision levels around survival point estimates and not the estimates themselves [59,60]. We also compared early (6-month) survival trajectories of the two release groups using a log-rank test [61] to assess any potential differences in early post-release survival between these groups. Differences in survival trajectories were considered significant at alpha < 0.05.
2.10. Release Success Indicators
Based on White et al. [22,43], we used the following parameters as metrics of reintroduction success:
- (1)
- Survival, assessed as the percentage of released individuals that survived at 3 months and 1 year following release.
- (2)
- Fidelity to release site, defined as the percentage of macaws that established a stable area of activity within 5 km of the release site, excluding any temporary long-distance forays.
- (3)
- Flock cohesion, defined as the percentage of surviving individuals that directly interacted (e.g., flying, foraging, perching, or roosting together). This includes any direct interactions or integration into wild heterospecific groups at or near the release site.
- (4)
- Prompt reproduction, defined here as reproduction (including egg laying) by released macaws during the initial 18 months post-release [43].
2.11. Social Engagement
Prior to and during the release and post-release monitoring period, we established projects aimed at disseminating information, engaging local communities, and providing opportunities for new environmentally sustainable income alternatives.
The entities involved in the project played an active role by engaging in visits to rural properties, where they elucidated the project’s objectives and distributed informative posters to facilitate recognition of Spix’s Macaw. Additionally, they took part in community association meetings and aired a program on local radio titled “Blue Hope,” where they explained the project and the significance of the protected areas.
Given that the most prevalent messaging application in the region is WhatsApp®, two dedicated chat groups were established. These groups served not only to update residents about project advancements but also as a platform for receiving feedback regarding bird sightings. Notably, on each occasion of bird dispersal, valuable insights and guidance from the community were transmitted through this channel.
The establishment of the protected area and the facility provided new employment opportunities for the locals in the entities involved in the project. Some projects with environment recovery and the payment of carbon or biodiversity credits are taking place in the protected areas, bringing the concept of sustainable income alternatives to local community members.
3. Results
3.1. Releases
3.1.1. First Release
The release module was opened at dawn on the day of release (11 June 2022). Blue-Winged Macaws were the first birds to leave the release module and utilize outside feeders. Approximately three hours afterwards, the first Spix’s Macaw exited the module and also visited an outside feeder (Figure 3). Throughout the day, five of eight Spix’s Macaws left the enclosure and engaged in exploratory flights around the release enclosure. During the afternoon, one male dispersed approximately 700 m from the release site after being disturbed by a low-flying Black Vulture (Coragyps atratus) and returned to the release site the following morning.

Figure 3.
The first captive-reared reintroduced Spix’s Macaw leaving the training and release enclosure on 11 June 2022. Photo credit: Tim Flach/ACTP.
Initially, released macaws occupied the area between the release and breeding enclosures and interacted with the breeding birds, as well as the birds in the center module of the release cage. They were frequently seen in small groups using this area, roosting either inside or on the roof of the release cages, and perching and feeding together with “mentor” Blue-Winged Macaws (Figure 4) in nearby trees, such as “quixabeira” (Sideroxylon obtusifolium), “catingueira” (Cenostigma pyramidale), “baraúna” (Schinopsis brasiliensis), “aroeira” (Myracrodruon urundeuva), mesquite (Neltuma juliflora), and “pereiro” (Aspidosperma pyrifolium), as well as engaging in flights with groups of up to 18 Blue-Winged Macaws. One week after release, some Spix’s Macaws were observed in catingueira trees, and 25 days after release, all eight Spix’s Macaws were observed foraging together in catingueira and pereiro trees (Figure 5). Thereafter, they were frequently seen foraging not only in catingueira (Figure 6) but also “brinco-de-soim” (Pithecellobium diversifolium).

Figure 4.
A released Spix’s Macaw foraging together with four Blue-Winged Macaws in a “quixabeira” tree (Sideroxylon obtusifolium) near the release site in the State of Bahia, Brazil, on 30 April 2023. Photo credit: Tyson Chapman/ACTP.

Figure 5.
All eight of the first group of released captive-reared Spix’s Macaws foraging together in “catinguera” (Cenostigma pyramidale) and “pereiro” (Aspidosperma pyrifolium) trees near the release site in the State of Bahia, Brazil, on 7 July 2022. Photo credit: Tyson Chapman/ACTP.

Figure 6.
A released captive-reared Spix’s Macaw foraging on seed pods of “catinguera” (Cenostigma pyramidale) near the release site in the State of Bahia, Brazil, on 25 July 2022. Photo credit: Tyson Chapman/ACTP.
After the first weeks of release, the released birds no longer used the release enclosure as a dormitory, roosting instead in trees near the enclosure or on the roof of the release enclosure together with the Blue-Winged macaws. This pattern continued until the breeding season, at which time some individuals formed pairs. At the onset of the breeding season, the Blue-Winged Macaws also ceased flying in large groups and visited the release enclosure less frequently. Progressively, some pairs or small groups of Spix’s Macaws also expanded their flight radius up to 3 km from the release area. The released birds also responded as a group to raptors by using mobbing and agonistic vocalizations, displayed especially towards the Black Vulture, Turkey Vulture (Cathartes aura), and Crested Caracara (Caracara plancus).
The first bonded pair began to disperse to the riparian gallery forests three months after the release. Copulation was observed in a Schinopsis brasiliensis tree near the breeding center, and they began to visit a nest box installed nearby. This bonded pair remained together until the predation of the male by a Crane Hawk (Geranospiza caerulescens) in October 2022. Subsequently, the female paired with another female and occupied the nest box near the breeding center, in which she laid a clutch of three eggs. A second pair was observed attempting to copulate and briefly established themselves as a pair, before separating for unknown reasons. One other Spix’s Macaw was lost to an unknown predator during September 2022. Thus, two (25%) Spix’s macaws released in Group 1 were predated.
3.1.2. Second Release
The second release (10 December 2022) was carried out in the same way and at the same time as in June. The birds took less time to leave the enclosures than during the first release: ten minutes for the Blue-Winged Macaws and one hour for the first Spix’s Macaws. Two days after release, one Spix’s Macaw was predated approximately 10 km from the release site, with puncture lesions in the head and neck consistent with raptor predation. One month later, another Spix’s Macaw was predated less than 3 km from the release site. Evidence found at the site (feathers, transmitter, mammalian scrape) suggested likely mammalian predation. A third predation event occurred in May 2023, when an Oncilla (Leopardus tigrinus) was seen leaping from a tree near the release site carrying a dead Spix’s Macaw. Three (25%) Spix’s Macaws released in Group 2 were predated during the first six months post-release, the same proportion as in Group 1.
The macaws of the second release group roosted less frequently in the release enclosure than birds of Group 1, generally using them only for feeding and diurnal resting. Roosting primarily occurred in trees near the release area, such as “leucena” (Leucaena leucocephala), Caribbean trumpet tree, “umbuzeiro” (Spondias tuberosa), “pau d’colher” (Maytenus rigida), coconut (Cocos nucifera), faveleira, “pau-ferro” (Libidibia ferrea), and “juazeiro” (Sarcomphalus joazeiro).
Compared to Group 1, birds in the second release subgroup interacted more with the captive breeding population in the breeding center, in addition to more actively exploring other areas within and outside the 5 km radius. Approximately one month after the second group release, local residents also began to report more frequent sightings of Spix’s Macaws flying and foraging in the local area. The birds released in June also changed their behavior and pairings after the second release. One previously bonded pair separated and began to interact with the macaws in the breeding enclosure, with each later forming apparent pair bonds with different individuals.
3.2. Success Indicators
3.2.1. Survival
Cumulative overall survival was 58.3% (CI: 37.8–78.7%) at one year following initial release (Figure 7). Survival at six months post-release was 71.4% (CI: 38–100%) for the first release group and 71.3% (CI: 43–99.6%) for the second group, and the survival trajectories did not differ (χ2 = 0.20, p = 0.65, df = 1) between groups (Figure 8). It is noteworthy that the post-release survival of the initial group was 100% up to three months post-release (Figure 7 and Figure 8), and it was 81.5% for the second group, indicating that the birds were released in good physical condition and able to quickly adapt to the release environment. In total, five Spix’s Macaws were predated and two could not be located (i.e., censored observations) by the end of the first year post-release (June 2023).
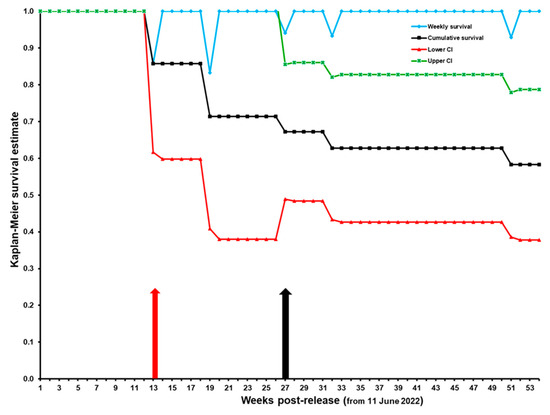
Figure 7.
Weekly and cumulative Kaplan–Meier survival trajectories for 20 captive-reared Spix’s Macaws released in two groups during 2022 in the State of Bahia, Brazil. The red arrow marks 3 months post-release (from 11 June 2022). The black arrow indicates the week of the second release.
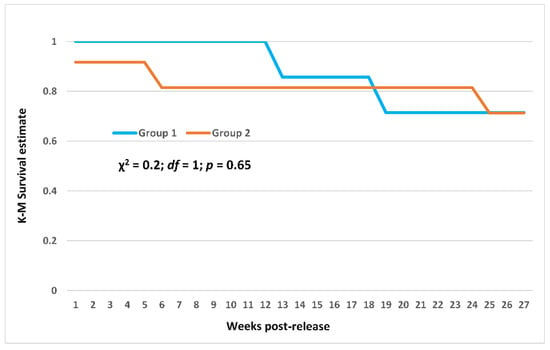
Figure 8.
Kaplan–Meier survival trajectories during the initial six months (27 weeks) post-release for two groups of captive-reared Spix’s Macaws released in 2022 in the State of Bahia, Brazil.
3.2.2. Site Fidelity and Flock Cohesion
After the first release, 62.5% (n = 5) of Spix’s Macaws exhibited site fidelity and settled within 5 km of the release site (excluding any temporary long-distance forays). However, after the second release, some pairs and groups expanded the area of use, and they also showed less group cohesion. Two Blue-Winged Macaws released with radio collars also dispersed from the reintroduced group at 5 and 46 days after release. Overall, 65% (n = 13) of all released Spix’s Macaws exhibited site fidelity and 85% (n = 17) also remained cohesive as a group, occupying the same areas together or moving out of the release area but returning daily or at most the next morning. Indeed, the group cohesion of the first release was 100% (e.g., Figure 5) during the first 2 months post-release.
By the end of the first year, 70% of the released Spix’s Macaws had also performed exploratory flights beyond a 5 km radius of the release site, with most returning to the area. Seven individuals dispersed and did not voluntarily return or otherwise disappeared from the release area. Three of them were subsequently rescued (see below) and reintegrated into the group, two were predated, and two disappeared (one of them had also been rescued previously). The dispersive movements and subsequent predations were associated with group flights in which one of the birds did not accompany the flock back to the release site.
The birds that dispersed following the first release event flew approximately 10 km/day [21]. After the second release, the Spix’s Macaws began to fly more than 30 km away from the release site in a single day, being detected by several local residents and then returning to the release site to roost at the end of the day. Following both the release events, Spix’s Macaws were frequently observed interacting with wild groups of Blue-Winged Macaws at or near the release site (e.g., Figure 4).
3.2.3. Reproduction Attempts
We observed at least six heterosexual pairs formed among the birds released. Two of the heterosexual pairs had taken a nest and defended it with similar results—both pairs were seen mating, but unfortunately they were split when the males, in both cases, were killed by predators. In both cases, the death of the paired male was followed by the females forming a same-sex pair (two different same-sex pairs). At the end of January, these same-sex pairings dissolved, resulting in two new same-sex pairs and a heterosexual pair. In February 2023, one female–female pair began to occupy a nest box installed near the release site, only leaving that location to forage. These female pair aggressively defended its territory and prevented other birds from entering the nesting tree. Two clutches of eggs were laid and incubated by this pair, before the pair separated, and one member of the pair subsequently formed a heterosexual pairing. The second same-sex pair also took possession of a nest and laid two clutches of eggs. The third same-sex pair, although aggressive around a nest box, never laid eggs. The heterosexual pair discovered a natural nest cavity in a Caribbean trumpet tree and was observed excavating and defending it. Of the total of six heterosexual pairs formed, four occupied nest boxes. Of the four occupied nests, two pairs split due to the death of a mate prior to egg laying. Subsequently, two heterosexual pairs occupied nests, with only one laying eggs. The first clutch from this heterosexual pair consisted of three infertile eggs, while the second clutch was also three eggs, two of which were fertile and hatched, and the chicks were successfully parent-reared to fledging age (Figure 9). However, during routine nest monitoring, we noticed that the parents were plucking feathers from the chicks as they approached fledging age. We hypothesized that daily temperatures up to 37 °C at the time had resulted in excessive heat in the nest box and a consequent behavioral response by the parents, as such behavior has previously been observed in captive populations of this species (Cromwell Purchase, pers. observ.). The monitoring team made a few minor alterations to the nest box to increase ventilation without major changes that could disrupt parental care. These changes were apparently successful, and the parents stopped plucking the chicks. However, an alternative hypothesis is that this aberrant behavior may also have been due to inexperienced captive-reared parents. Nevertheless, shortly before fledging, one nestling was found dead inside the nest with a femoral laceration and contusions of unknown cause(s). The second nestling fledged from the nest on 8 November 2023, but it was subsequently found dead with cranial trauma on 17 November 2023. Direct behavioral observations and circumstances indicated that the cause of death was due to an agonistic interaction with another unidentified macaw. This was the first record of captive-reared Spix’s macaws breeding in the wild post-release.

Figure 9.
Two Spix’s Macaw nestlings (left panel) inside an artificial nest near the reintroduction site in the Caatinga forest domain of Brazil. Photo taken 1 September 2023. These were the first chicks to hatch following the reintroduction of the species in 2022, and (right panel) one of the nestling Spix’s Macaws a few days prior to its fledging on 8 November 2023. This was the first record of a Spix’s Macaw fledging in the wild in over 30 years. Photo credit: Cromwell Purchase/ACTP (left panel), Candice Purchase/ACTP (right panel).
3.2.4. Local Community Engagement
Due to effective local community interaction and engagement, we were successful in finding and rescuing four birds who disappeared post-release. Following the June 2022 release, two birds left the release area for extended periods, where they traveled far enough that they were unlikely to return unassisted. The community was informed that they were missing and asked to keep vigilant and report any sightings as quickly as possible. The community response was overwhelming in all instances. The community’s quick response to sightings gave our monitoring teams an area and direction in which to search; this response, combined with radio telemetry, made for successful recapture and return operations. A similar event occurred after the December 2022 release, when two birds also left the release area and traveled beyond an area in which there could be a likelihood of returning unassisted, and they were subsequently located and returned with the help of information from members of the local community who informed our monitoring team of their last sighted locations. WhatsApp® groups and social networks were, thus, valuable communication channels, as local residents quickly reported sightings of the birds, which aided post-release tracking. The local radio station also proved effective for communicating the results obtained and allowing engagement with local communities.
4. Discussion
4.1. Success of the Release
The reintroduction of Spix’s Macaw represents one of the most challenging endangered species management and recovery plans ever implemented in Brazil. However, extensive planning for species recovery yielded effective outcomes [62], and the initial experience of Spix’s Macaw releases was considered successful, based on our stated criteria and those of White et al. [22], which categorized as successful those psittacine reintroductions with >50% first-year survival and subsequent reproduction by released birds. Our primary success indicator exceeded our initial projected goals of at least 30% first-year survival. We established the target goals based on the results obtained by Collazo et al. [60] for captive-reared Hispaniolan Parrots (Amazona ventralis) and White et al. [29,43,49] for captive-reared Puerto Rican Parrots (Amazona vittata). We achieved nearly 60% cumulative survival one year after the first release event and over 70% at 6 months after each release event. We hypothesize that the success indicators will increase over the coming years concomitantly with increasing numbers of Spix’s Macaws in the wild [43,63]. Indeed, according to a recent Population Viability Analysis (PVA) for the species [21], annual releases of around 20 Spix’s Macaws at the current release site for the next 20 years will be critical to the long-term persistence of the species on the landscape. The importance of regular population supplementation and continued support of this nascent wild population cannot be overemphasized.
We also witnessed the formation of at least six heterosexual pairs, one of which (male from first and female from second release) nested and laid eggs in an artificial nest box near the breeding facility. This pair produced two clutches of eggs. The first was of three eggs, and all were infertile. The second clutch was also three eggs, with two being fertile. Significantly, both fertile eggs hatched, and the naïve first-time parents successfully fed the chicks. Unfortunately, both nestlings eventually died; one died shortly before fledging, and the other died several days after fledging. Regardless, this breeding event—the first known successful nesting in the wild by Spix’s Macaws in over 30 years—within a short time post-release increases the probability of more successful reproduction (i.e., recruitment) during the next (2024) breeding season.
We expected release site fidelity and group cohesion of approximately 67% at one year after release, based on White et al. [43]. Overall, 85% of the released Spix’s Macaws exhibited group cohesion, either occupying the same areas together or dispersing and managing to return to the release site daily. Not surprisingly, we observed greater group cohesion following the first release event, as it occurred during the non-breeding season. The first group of Spix’s Macaws established a core use area near and adjacent to the release and breeding cages, and group excursions did not exceed 5 km, which facilitated monitoring and likely promoted greater survival. However, after the second release, some birds dispersed up to 50 km, which might have mainly been a consequence of the following factors: (1) formation of pairs and search for nests in the breeding season, (2) formation of groups and pairs with released individuals that already had experience in habitat use, and (3) abundance of fruits in the rainy season. Therefore, we believe that releases carried out early in the dry season, when birds are concentrated in larger groups and can more readily access supplementary feeding, may increase cohesion and fidelity to the release area. Thereafter, independence can occur gradually as the birds steadily improve their abilities to search for food and protect themselves from predators.
The first months of release are typically the ones with the highest risk, especially due to predation [23,43,49]. Birds of prey likely represent the main threat to the reintroduction of Spix’s Macaw, as also reported elsewhere [43,49]. It is noteworthy that predations primarily occurred with Individuals that dispersed from a group, either as singletons or as pairs, and did not return to the release site. Such individuals are more likely to be predated, as has been reported for captive-raised Puerto Rican Parrots [49].
4.2. Tracking
Post-release monitoring is a fundamental component of reintroductions [23], enabling the evaluation of their progress and facilitating necessary adjustments [64]. However, it is one of the most difficult components to implement [65] due to a variety of factors including the unpredictable behavior of the released animals and consequent long-distance dispersal away from the release site [66]. The topography and remoteness of the release site, the availability of skilled personnel, and the existence of suitable tracking equipment are also factors that increase the difficulty of monitoring [65]. Accurately determining post-reintroduction survival for macaws is another challenging and expensive issue related to the reintroductions [67]. Monitoring Spix’s Macaws individually marked with radio collars was our best available method for the species, given the characteristics of the species and the current lack of a suitable alternative (GPS or 3G) available for a parrot of this size. Transmitters attached in neck collars withstood the strong beaks and allowed macaws to easily manipulate food items and engage in allopreening behavior [68].
Blue-Winged Macaws are extremely mobile, and their movements tend to vary according to landscape composition [69,70]. The released Spix’s Macaws did not exhibit the same movement pattern as the Blue-Winged Macaws in the area; however, the monitoring of the released birds was a challenge with VHF technology. The inclusion of the local residents and monitoring teams formed by local citizens improved our post-release monitoring. Mobile technology and citizen science has the capability to increase data collection associated with less expensive methods [71,72,73]. The widespread use of “smart” devices, combined with applications that provide georeferenced photos, makes them both a rapid and a rich source for data reporting [74,75,76] that is spatially and temporally accurate. Conversely, direct interaction between Spix’s Macaws and people can potentially weaken the reintroduction process, causing habituation and dependence on inappropriate food or even poaching. We recorded the disappearance of two birds; one of them disappeared suddenly and may have been the victim of poaching. It is also possible that this bird could have flown beyond the range of monitoring and not been seen by members of the local communities. Systematic searches and investigative actions were carried out throughout the region but were not successful.
The transmitter signals resulted in at least six months of reliable detections of most released birds, with some transmitters functioning for up to a year. Due to the relatively flat terrain, the maximum range of signal reception was up to 7 km from elevated sites such as hills or steeper slopes. Therefore, for future Spix’s Macaw telemetry, we suggest the use of fixed receptors placed strategically throughout the release area, utilizing topographical features (e.g., hilltops) to improve signal detection, pending the future development of an effective GPS-style device for this species and area.
4.3. The Surrogate Model Species
To the best of our knowledge, Spix’s Macaw reintroduction is the first such effort for parrots that deliberately employed a “mixed species model” for maximizing the survival and post-release adaptation of the target species. Our use of heterospecific release groups allowed an increase in total cohort size without a concomitant increase in the actual number of Spix’s Macaws initially at risk. Wild-sourced Blue-Winged Macaws directly interacted with Spix’s Macaws during pre-release training and later served as an effective “mentor” for released Spix’s Macaws, who followed and mimicked the actions of the Blue-Winged Macaws with which they were released (e.g., Figure 4). During both releases, Blue-Winged Macaws were also the first to exit the release enclosure and begin foraging on native foods, followed soon thereafter by Spix’s Macaws. Blue-Winged Macaws also quickly recognized potential predators and gave appropriate alarm calls to which Spix’s Macaws also responded, as has also been reported with Superb Fairy-Wrens (Malurus cyaneus) and White-Browed Scrubwrens (Sericornis frontalis) in Australia [34]. Indeed, one of the primary advantages of mixed-species flocks is an increase in predator detection and avoidance by all member species [34,35,39], particularly when member species share common predators [39], as is the case with Spix’s and Blue-Winged Macaws. This mutualistic interspecific communication and increased efficiency of predator detection [36] further leads to increased foraging efficiency via a reduction in individual vigilance requirements [35,38]. For example, Sridhar et al. [37] reported increased foraging rates and decreased scanning rates in mixed-species foraging flocks compared to intraspecific flocks, and they believed that participation in mixed-species flocks could result in higher fitness. Accordingly, we believe that the presence of co-released Blue-Winged Macaws likely increased Spix’s Macaws’ probability of survival and adaptation post-release, thereby lessening some of the inherent Allee effects associated with early phases of reintroductions [42,43,77,78].
Although, initially, we did not record the frequent use of riparian gallery forests by released Spix’s Macaws, we predicted the progressive use of these habitats based on observed behavior of the Blue-Winged Macaw. Silva [79] estimated the density of P. maracana in the historical occurrence area of Spix’s Macaw and, when calibrated with the Soil-Adjusted Vegetation Index (SAVI), indicated a strong relationship between the species and arboreal vegetation in the riparian forest. Such a relationship may also be indicative of both historical and future habitat preference of the sympatric and ecologically similar Spix’s Macaw [14,80]. Indeed, Cavalcanti et al. [81] showed a spatial correlation of 85.5% between the historical occurrence of Spix’s Macaw and landscapes of seasonal rivers throughout the Caatinga. The floodplain landscape harbors a dense arboreal vegetation covering a width of up to 1 km (e.g., Figure 2). Caribbean trumpet trees, as well as baraúna, quixabeira, “marizeiro” (Geoffroea spinosa), and juazeiro, are common elements of the emergent flora [81]. Reintroduced Spix’s Macaws occupying such habitat patches could eventually function as a source population for the colonization of other areas and may be vital for maintaining the species’ regional occurrence [82].
4.4. Protected Areas
Preserved habitats are considered vital to the conservation of large-bodied parrots in the semiarid region [83]. The protected areas for Spix’s Macaw comprise 120,000 hectares [24], but habitat quality is not homogenous across most of the landscape [81]. Grazing and selective cutting has had a significant impact on riparian vegetation in the region [81]. Dias et al. [84] demonstrated that the presence of cattle and donkeys within the protected area in Caatinga may represent a severe threat to the local biotas, resulting in the extensive degradation of its vegetation and hampering the recuperation of native habitats, as well as jeopardizing the survival of its native species, reducing water quality, and eroding ecosystem services. Moreover, climate projections for the protected areas of Spix’s Macaw showed a reduction in the area of environmental suitability that overlaps the distribution of key resources for Spix’s Macaw [85]. Therefore, the long-term suitability of the protected areas for Spix’s Macaw is dependent on continued restoration of degraded areas, especially riparian forests. Accordingly, an integrated strategy to recover the riparian vegetation related to alluvial plain landforms must include the fragment of this vegetation in the protected area.
The survival of the reintroduced Spix’s Macaw also depends on the co-operation of local and regional landowners in the conservation of remnants of native vegetation on private lands, as the protected areas and surrounding landscape consist of privately owned lands. Larger forested fragments containing suitable nest sites and roosts, as well as small patches of trees within the landscape matrix, may be important for increasing the landscape connectivity and geographic expansion of both macaw species’ populations [54].
4.5. Human Interaction
Some human-related difficulties related to the reintroduction of parrots are poaching, as well as interactions with people who may feed birds and make them vulnerable to, or dependent on, humans [86,87]. The low socioeconomic indices among rural communities in semiarid areas, such as the Caatinga, can complicate efforts to restrain illegal captures and wildlife trade [88]. To maximize the probability of a successful reintroduction, the support of local communities is vital in our reintroduction approach [89,90]. The interactions between people and wildlife sharing the same space and resources are factors considered in our perspective. Therefore, we considered the engagement of local people to be a critical first step for the success and effectiveness of our conservation programs [91]. Based on this assumption, we first created volunteer opportunities directly linked to monitoring via telemetry, sightings, reports, and a network of sharing data. The training and information sharing provides long-term benefits to the project by forming a new generation of citizen scientists in the Spix’s Macaw reintroduction area, making future participatory monitoring more likely and effective.
The human dimensions of human–bird interactions and the reintroduction implications for the local communities are still poorly known or underexplored in reintroduction projects in Brazil [87]. However, the Spix’s Macaw reintroduction project is an exception in several aspects in terms of social engagement and communities’ participation in resource conservation. Furthermore, we created two protected areas to support the reintroduction project [21], an effort that included active participation by members of local communities. In this way, local residents became stakeholders in the species reintroduction project via active participatory dialogue between scientific and traditional knowledge [92]. These proactive actions can also help to reduce and control the illegal trade in birds [93,94].
Conservation reintroductions are truly multi-disciplinary endeavors [65]. The knowledge, values, and interactions of local residents with wildlife are seldom considered in most reintroduction projects [23,87]. The Spix’s Macaw project proactively engaged with local communities as part of pre-release reintroduction planning. These actions are positively connected and preventive and utilize a behavioral change approach [87,93,94]. Moreover, local community engagement and volunteer programs are the most effective ways to integrate local residents into project actions and potentially decrease difficulties involved in maintaining long-term financial support for the project [65].
5. Conclusions
Our results suggest that the Spix’s Macaw reintroduction program began with successful and encouraging results. After over 20 years of extinction in the wild, the first free-flying individuals of the species once again occupy and nest in their historical native habitat. Importantly, the first breeding of Spix’s macaws in the wild in over 30 years occurred within nine months of the second release, suggesting that the captive-reared birds were not only quickly acclimatizing to their habitat but also healthy enough to reproduce. Most encouragingly, the breeding event demonstrates that Spix’s Macaws that have been bred for multiple generations in captivity are still capable of successfully hatching and rearing chicks in the wild. However, to ensure long-term population persistence, it will be necessary to continue regular annual releases of approximately 20 captive-reared birds at the release site over the next 20 years [21].
The reintroduction planning process focused on developing a detailed conservation action plan for a single species, leading to a practical blueprint of actions designed to mitigate biological and anthropogenic threats to Spix’s Macaws’ population persistence and restore an extinct-in-the-wild species in its native habitat. The novel use of a “mixed species model” for the initial reintroduction cohorts proved an effective strategy for maximizing the post-release survival and adaptation of a species that had been reared for multiple generations in captivity and released in the absence of wild conspecifics. The reintroduction program also included actions designed to address diverse social and institutional challenges involved in achieving recovery goals. The protected area of Spix’s Macaw appears capable of supporting the new population, considering current habitat quantity and quality across the surrounding landscape. The high mobility of parrot species in general allows them to effectively exploit small patches of suitable habitat within fragmented landscapes [54,95,96], further highlighting the importance of more broad-scale conservation of similar habitats beyond the current protected areas.
Considering the substantial gap that typically exists between the funding needed for biodiversity conservation and the amount actually devoted to it, citizen science, in conjunction with less expensive methods such as bird marking and passive monitoring, has the capability to increase data acquisition. Accordingly, the hiring of adequately trained local residents can provide a means of direct communication and interaction with local communities and increase their involvement in, and support for, conservation efforts. People are more prone to engage positively with conservation if they are included in a participatory process [90,92,93].
Despite difficulties in its use in remote areas, where internet coverage is often insufficient, we also recommend using social media as a valuable adjunct to volunteer reporting of sightings of reintroduced birds. Local residents are considered valuable human resources and essential for protecting the focal species. For Spix’s Macaw reintroduction, they are also key participants in decisions that would affect them directly and indirectly. Spix’s Macaw serves as an effective flagship species in the conservation of the anthropogenically altered environment of the Caatinga and other native and endemic species dependent thereon, and its conservation and restoration in the wild will promote inclusive and sustainable socio-environmental development to support the vulnerable human populations that also share this unique habitat.
Author Contributions
C.P. (Cromwell Purchase), C.L., M.L.S. and T.H.W.J. designed the project. C.P. (Cromwell Purchase), C.P. (Candice Purchase), A.F., T.H.W.J., U.E.V. and C.L. observed and monitored the released Spix’s Macaws. C.L., A.F. and T.H.W.J. organized the dataset and conducted analyses. All authors have read and agreed to the published version of the manuscript.
Funding
Funding and logistical support for this project was provided by Vale Company, who sponsored work by ICMBio and Projeto Ararinha na Natureza. Additional support for all aspects of the project was provided by ACTP (Association for the Conservation of Threatened Parrots), GZRRC (Greens Zoological, Rescue and Rehabilitation Centre), BlueSky, Pairi Daiza Foundation, Wachtelland Park, ZGAP (Zoologische Gesellschaft für Arten-und Populationsschutz), Wisbroek, and Karl Matthes.
Institutional Review Board Statement
All activities were authorized and carried out according to SISBIO/ICMBio scientific permit numbers 53258, 79010, and 81900 and the Authorization of Fauna Management Nº 638484, INEMA-BA (expiration date 10 March 2025), as well as Wildlife Release Certificate Nº 16/2022.
Data Availability Statement
Due to the continuing high risk posed to this critically endangered species from illegal poaching and commercialization, specific locational data regarding the reintroduction are not being made public, and they may be requested upon consultation directly with, and at the discretion of, the lead author.
Acknowledgments
We thank all of the institutions and numerous local residents who supported these efforts, including ProCom/Grumbach, NuTropica®, Birdfarm.de, Kauri CAB, AVES publishing, Bowman Books, Al Wabra, Parrots International, and Museum Für. We are also grateful for the valuable support of ICMBio firefighters and local residents who assisted with the monitoring of the species, as well as all the local community residents who promptly reported sightings of missing macaws. Personnel with CEMAFAUNA/UNIVASF also assisted with searches for missing macaws and the control of Africanized bees. Four anonymous reviewers provided constructive comments for improving an earlier version of the manuscript. The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service. The use of trade names in this article does not imply endorsement by the United States government.
Conflicts of Interest
The authors declare that there are no conflicts of interest. The funding organizations had no role in the design, execution, interpretation, or writing of this study.
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R. The misunderstood sixth mass extinction. Science 2018, 360, 1080–1081. [Google Scholar] [CrossRef]
- Vergara-Tabares, D.L.; Cordier, J.M.; Landi, M.A.; Olah, G.; Nori, J. Global trends of habitat destruction and consequences for parrot conservation. Global. Chang. Biol. 2020, 26, 4251–4262. [Google Scholar] [CrossRef] [PubMed]
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.J.; Abbud, M.C.; Aguilar, J.; Alemán-Zelaya, U.; Aramburú, R.M.; Arce Arias, A.; Balas McNab, R.; Balsby, T.J.S.; et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef]
- Olah, G.; Butchart, S.H.M.; Symes, A.; Guzmán, I.M.; Cunningham, R.; Brightsmith, D.J.; Heinsohn, R. Ecological and socio-economic factors affecting extinction risk in parrots. Biod. Conserv. 2016, 25, 205–223. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Lowe, S.; Martin, R.W.; Symes, A.; Westrip, J.R.S.; Wheatley, H. Which bird species have gone extinct? A novel quantitative classification approach. Biol. Conserv. 2018, 227, 9–18. [Google Scholar] [CrossRef]
- BirdLife International. Species Factsheet: Cyanopsitta spixii. 2019. Available online: https://datazone.birdlife.org/species/factsheet/spixs-macaw-cyanopsitta-spixii (accessed on 4 December 2023).
- Sick, H. Ornitologia Brasileira; Nova Fronteira: Rio de Janeiro, Brazil, 1997. [Google Scholar]
- Kupferschmidt, K. A wild hope. Science 2022, 376, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Juniper, A.T. Spix´s Macaw: The Race to Save the World´s Rarest Bird; Washington Square Press: New York, NY, USA, 2002; p. 293. [Google Scholar]
- Roth, P. Bericht fiber ein 1985–1988 durchgefuhrtes Projekt-1. Papageien 1990, 90, 86–88. [Google Scholar]
- Roth, P. Bericht fiber ein 1985–1988 durchgefuhrtes Projekt-2. Papageien 1990, 90, 121–125. [Google Scholar]
- Juniper, A.T.; Yamashita, C. The conservation of Spix´s macaw. Oryx 1990, 24, 224–228. [Google Scholar] [CrossRef]
- Juniper, A.T.; Yamashita, C. The habitat and status of Spix’s Macaw Cyanopsitta spixii. Bird Conserv. Int. 1991, 1, 1–9. [Google Scholar] [CrossRef]
- Barros, Y.M.; Soye, Y.; Miyaki, C.Y.; Watson, R.; Crosta, R.L.; Lugarini, C. Plano de Ação Nacional Para a Conservação da Ararinha-azul: Cyanopsitta spixii; Instituto Chico Mendes de Conservação da Biodiversidade: Brasilia, Brazil, 2012; p. 145. [Google Scholar]
- Barnett, J.M.; Silva, C.L.G.; Araujo, H.F.P.; Roos, A.L.; Machado, C.G.; Uejima, A.M.K.; Naka, L.N. The avifauna of Curaçá (Bahia): The last stronghold of Spix’s Macaw. Rev. Bras. Ornit. 2014, 22, 121–137. [Google Scholar] [CrossRef]
- Barros, Y.M. Biologia Comportamental de Propyrrhura maracana (Aves, Psittacidae): Fundamentos Para Conservação In Situ de Cyanopsitta spixii (Aves, Psittacidae) na Caatinga; Universidade Estadual de São Paulo: São Carlos, Brazil, 2001; p. 97. [Google Scholar]
- Miyaki, C.Y.; Faria, P.J.; Griffiths, R.; Araujo, J.C.C.; Barros, Y.M. The last Spix’s Macaw and an Illiger’s Macaw produced a hybrid. Conserv. Gen. 2001, 2, 53–55. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Barros, Y.M.; Develey, P.F.; Llerandi-Román, I.C.; Monsegur-Rivera, A.O.; Trujillo-Pinto, A.M. Improving reintroduction planning and implementation through quantitative SWOT analysis. J. Nat. Conserv. 2015, 28, 149–159. [Google Scholar] [CrossRef]
- Lugarini, C.; Vercillo, E.U.; Purchase, C.; Watson, R.; Schischakin, N.A. Conservação da Ararinha-azul, Cyanopsitta spixii (Wagler, 1832): Desafios e Conquistas. Biodiversidade Bras. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Vercillo, U.; Oliveira-Santos, L.G.; Novaes, M.; Purchase, C.; Purchase, C.; Lugarini, C.; Ferreira, A.; Marco, P.; Marcuk, V.; Franco, J.L.A. Spix’s Macaw (Cyanopsitta spixii (Wagler, 1832)) population viability analysis. Bird Conserv. Int. 2023, 33, e67. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collar, N.J.; Moorhouse, R.J.; Sans, V.; Stolen, E.D.; Brightsmith, D.J. Psittacine reintroductions: Common denominators of success. Biol. Conserv. 2012, 148, 106–115. [Google Scholar] [CrossRef]
- IUCN/SSC. Guidelines for Reintroductions and Other Conservation Translocations, Version 1.0; IUCN Species Survival Commission: Gland, Suisse, 2013; p. 57. ISBN 978-2-8317-1609. [Google Scholar]
- Gobierno do Brasil. Decreto n° 9.402, de 5 de junho de 2018. Cria o Refúgio de Vida Silvestre da Ararinha Azul e a Área de Proteção Ambiental da Ararinha Azul. Available online: https://www.planalto.gov.br/ccivil_03_ato2015-2018/2018/decreto/d9402.htm (accessed on 14 February 2023).
- Marcuk, V.; Purchase, C.; de Boer, D.; Bürkle, M.; Scholtyssek, K. Qualitative description of the submission and agonistic behavior of the Spix’s Macaw (Cyanopsitta spixii, Spix 1824), with special reference to the displacement displays. J. Ethol. 2020, 38, 253–270. [Google Scholar] [CrossRef]
- Lugarini, C.; Vercillo, U. Como realizar a gestão de um projeto de alto risco? O relato da repatriação das Ararinhas-azuis ao Brasil. Biodiversidade Bras. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Ford, M.J. Selection in captivity during supportive breeding may reduce fitness in the wild. Conserv. Biol. 2002, 16, 815–825. [Google Scholar] [CrossRef]
- Frankham, R. Genetic adaptation to captivity in species conservation programs. Mol. Ecol. 2008, 17, 344–350. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collazo, J.A.; Dinsmore, S.J.; Llerandi-Román, I. Niche restriction and conservatism in a neotropical psittacine: The case of the Puerto Rican parrot. In Habitat Loss: Causes, Effects on Biodiversity and Reduction Strategies; Devore, B., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 1–83. [Google Scholar]
- Lenormand, T. Gene flow and the limits to natural selection. Trends Ecol. Evol. 2002, 17, 183–189. [Google Scholar] [CrossRef]
- Holt, R.D. Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives. Proc. Nat. Acad. Sci. USA 2009, 106, 19659–19665. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.H. Feeding behavior and predator avoidance in heterospecific groups. Bioscience 1977, 27, 332–339. [Google Scholar] [CrossRef]
- Herrera, C.M. Ecological aspects of heterospecific flocks formation in a Mediterranean passerine bird community. Oikos 1979, 33, 85–96. [Google Scholar] [CrossRef]
- Magrath, R.D.; Pitcher, B.J.; Gardner, J.L. A mutual understanding? Interspecific responses by birds to each other’s aerial alarm calls. Behav. Ecol. 2007, 18, 944–951. [Google Scholar] [CrossRef]
- Thiollay, J.M. Frequency of mixed species flocking in tropical forest birds and correlates of predation risk: An intertropical comparison. J. Avian Biol. 1999, 30, 282–294. [Google Scholar] [CrossRef]
- Goodale, E.; Kotagama, S.W. Response to conspecific and heterospecific alarm calls in mixed-species bird flocks of a Sri Lankan rainforest. Behav. Ecol. 2008, 19, 887–894. [Google Scholar] [CrossRef]
- Sridhar, H.; Beauchamp, G.; Shanker, K. Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Anim. Behav. 2009, 78, 337–347. [Google Scholar] [CrossRef]
- Martinez, A.E.; Zenil, R.T. Foraging guild influences dependence on heterospecific alarm calls in Amazonian bird flocks. Behav. Ecol. 2012, 23, 544–550. [Google Scholar] [CrossRef]
- Martinez, A.E.; Parra, E.; Gomez, J.P.; Vredenburg, V.T. Shared predators between primate groups and mixed species bird flocks: The potential for forest-wide eavesdropping networks. Oikos 2022, 2022, e0974. [Google Scholar] [CrossRef]
- Stephens, P.A.; Sutherland, W.J. Consequences of the Allee effect for behaviour, ecology and conservation. Trends Ecol. Evol. 1999, 14, 401–405. [Google Scholar] [CrossRef]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Inverse density dependence and the Allee effect. Trends Ecol. Evol. 1999, 14, 405–410. [Google Scholar] [CrossRef]
- Armstrong, D.P.; Wittmer, H.U. Incorporating Allee effects into reintroduction strategies. Ecol. Res. 2011, 26, 687–695. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Abreu, W.; Benitez, G.; Jhonson, A.; Lopez, M.; Ramirez, L.; Rodriguez, I.; Toledo, M.; Torres, P.; Velez, J. Minimizing potential Allee effects in psittacine reintroductions: An example from Puerto Rico. Diversity 2021, 13, 13. [Google Scholar] [CrossRef]
- Brooks, T. Are unsuccessful avian invaders rarer in the native range than successful invaders? In Biotic Homogenization; Lockwood, J.L., McKinney, M.L., Eds.; Kluwer Academic: New York, NY, USA, 2001. [Google Scholar]
- Cassey, P. Life history and ecology influences establishment success of introduced land birds. Biol. J. Linnean Soc. 2002, 76, 465–480. [Google Scholar] [CrossRef]
- Prinzing, A.; Durka, W.; Klotz, S.; Brandl, R. Which species become aliens? Evol. Ecol. Res. 2002, 4, 385–405. [Google Scholar]
- Cassey, P.; Blackburn, T.M.; Russell, G.J.; Jones, K.E.; Lockwood, J.L. Influences on the transport and establishment of exotic bird species: An analysis of the parrots (Psittaciformes) of the world. Global Chang. Biol. 2004, 10, 417–426. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Cassey, P.; Lockwood, J.L. The role of species traits in the establishment success of exotic birds. Global Chang. Biol. 2009, 15, 2852–2860. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collazo, J.A.; Vilella, F.J. Survival of captive-reared Puerto Rican Parrots released in the Caribbean National Forest. Condor 2005, 107, 426–434. [Google Scholar] [CrossRef]
- Vaz, F.F.; Serafini, P.P.; Locatelli-Dittrich, R.; Meurer, R.; Durigon, E.L.; Araújo, J.; Thomazelli, L.M.; Ometto, T.; Sipinski, E.A.B.; Sezerban, R.M.; et al. Survey of pathogens in threatened wild Red-tailed Amazon Parrot (Amazona brasiliensis) nestlings in Rasa Island, Brazil. Braz. J. Microbiol. 2017, 48, 747–753. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Abreu-González, W. Dummy transmitters for pre-release acclimation of captive-reared birds. Re-introduction News 2007, 26, 28–30. [Google Scholar]
- Kenward, R.E. A Manual for Wildlife Tagging; Academic Press: London, UK, 2001; p. 350. [Google Scholar]
- Godfrey, J.D.; Bryant, D.M.; Williams, M.J. Radio-telemetry increases free-living energy costs in the endangered Takahe Porphyrio mantelli. Biol. Conserv. 2003, 114, 35–38. [Google Scholar] [CrossRef]
- Davenport, L.C.; Boorsman, T.; Carrara, L.; Antas, P.T.Z.; Faria, L.; Brightsmith, D.J.; Herzog, S.K.; Soria-Auza, R.W.; Hennessey, A.B. Satellite telemetry of Blue-throated Macaws in Barba Azul Nature Reserve (Beni, Bolivia) reveals likely breeding areas. Diversity 2021, 13, 564. [Google Scholar] [CrossRef]
- Vilarta, M.R.; Wittkoff, W.; Lobato, C.; Oliveira, R.d.A.; Pereira, N.G.P.; Silveira, L.F. Reintroduction of the Golden Conure (Guaruba guarouba) in northern Brazil: Establishing a population in a protected area. Diversity 2021, 13, 198. [Google Scholar] [CrossRef]
- Koenig, S.E.; Wunderle, J.M., Jr.; Enkerlin-Hoeflich, E.C. Vines and canopy contact: A route for snake predation on parrot nests. Bird Conserv. Inter. 2007, 17, 1–15. [Google Scholar] [CrossRef]
- Pollock, K.H.; Moore, C.T.; Davidson, W.R.; Kellogg, F.E.; Doster, G.L. Survival rates of Bobwhite quail based on band recovery analyses. J. Wildl. Manage. 1989, 3, 1–6. [Google Scholar] [CrossRef]
- Pollock, K.H.; Winterstein, S.R.; Conroy, M.J. Estimation and analysis of survival distributions for radio-tagged animals. Biometrics 1989, 45, 99–109. [Google Scholar] [CrossRef]
- Tsai, K.; Pollock, K.H.; Brownie, C. Effects of violation of assumptions for survival analysis methods in radiotelemetry studies. J. Wildl. Manag. 1999, 63, 1369–1375. [Google Scholar] [CrossRef]
- Collazo, J.A.; White, T.H., Jr.; Vilella, F.J.; Guerrero, S.A. Survival of captive-reared Hispaniolan Parrots released in Parque Nacional del Este, Dominican Republic. Condor 2003, 105, 198–207. [Google Scholar] [CrossRef]
- Harrington, D.P.; Fleming, T.R. A class of rank test procedures for censored survival data. Biometrika 1982, 69, 553–566. [Google Scholar] [CrossRef]
- Byers, O.; Copsey, J.; Lees, C.; Miller, P.; Traylor-Holzer, K. Reversing the decline in threatened species through effective conservation planning. Diversity 2022, 14, 754. [Google Scholar] [CrossRef]
- Brightsmith, D.J.; Hilburn, J.; del Campo, A.; Boyd, J.; Frisius, M.; Frisius, R.; Janik, D.; Guillén, F. The use of hand-raised psittacines for reintroduction: A case study of scarlet macaws (Ara macao) in Peru and Costa Rica. Biol. Conserv. 2005, 121, 465–472. [Google Scholar] [CrossRef]
- Seddon, P.J.; Armstrong, D.P.; Maloney, R.F. Developing the science of reintroduction biology. Conserv. Biol. 2007, 21, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Berger-Tal, O.; Blumstein, D.T.; Swaisgood, R.R. Conservation translocations: A review of common difficulties and promising directions. Animal Conserv. 2019, 23, 121–131. [Google Scholar] [CrossRef]
- Harrington, L.A.; Moehrenschlager, A.; Gelling, M.; Atkinson, R.P.; Hughes, J.; Macdonald, D.W. Conflicting and complementary ethics of animal welfare considerations in reintroductions. Conserv. Biol. 2013, 27, 486–500. [Google Scholar] [CrossRef]
- Williams, S.; Haines, J. Scarlet macaw reintroduction on the Nicoya Peninsula of Costa Rica. In Global Conservation Translocation Perspectives: 2021; Soorae, P.S., Ed.; Case studies from around the globe; IUCN SSC Conservation Translocation Specialist Group, Environment Agency—Abu Dhabi and Calgary Zoo: Calgary, AB, Canada, 2021; pp. 133–136. [Google Scholar]
- Meyers, J.M. Evaluation of three radio transmitters and collar designs for Amazona. Wildl. Soc. Bull. 1996, 24, 15–20. [Google Scholar]
- Evans, B.E.I.; Ashley, J.; Marsden, S.J. Abundance, habitat use, and movements of Blue-winged Macaws (Primolius maracana) and other parrots in and around an Atlantic forest reserve. Wilson Bull. 2005, 117, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.F.C.; Galetti, M. Use of forest fragments by Blue-winged Macaws (Primolius maracana) within a fragmented landscape. Biodiv. Conserv. 2007, 16, 953–967. [Google Scholar] [CrossRef]
- Senar, J.C.; Carrillo-Ortiz, J.; Arroyo, L. Numbered neck collars for long-distance identification of parakeets. J. Field Ornithol. 2012, 83, 180–185. [Google Scholar] [CrossRef]
- Barve, V. Discovering and developing primary biodiversity data from social networking sites: A novel approach. Ecol. Inf. 2014, 24, 194–199. [Google Scholar] [CrossRef]
- MacPhail, V.J.; Colla, S.R. Power of the people: A review of citizen science programs for conservation. Biol. Conserv. 2020, 249, 1–15. [Google Scholar] [CrossRef]
- Burr, D.; Kline, J.; Perryman, A. A smartphone application for monitoring gopher tortoises in Florida. Fla. Sci. 2014, 77, 198–203. [Google Scholar]
- Pennington, C.; Freeborough, K.; Dashwood, C.; Dijkstra, T.; Lawrie, K. The national landslide database of Great Britain: Acquisition, communication and the role of social media. Geomorphology 2015, 249, 44–51. [Google Scholar] [CrossRef]
- Liebenberg, L.; Steventon, J.; Brahman, N.; Benadie, K.; Minye, J.; Langwane, H.; Xhukwe, Q. Smartphone Icon User Interface design for non-literate trackers and its implications for an inclusive citizen science. Biol. Conserv. 2016, 208, 155–162. [Google Scholar] [CrossRef]
- Deredec, A.; Courchamp, F. Importance of the Allee effect for reintroductions. Écoscience 2007, 14, 440–451. [Google Scholar] [CrossRef]
- Somers, M.J.; Graf, J.A.; Szkman, M.; Slotow, R.; Gusset, M. Dynamics of a small re-introduced population of wild dogs over 25 years: Allee effects and the implications of sociality for endangered species’ recovery. Oecologia 2008, 158, 239–247. [Google Scholar] [CrossRef]
- Silva, C.L.G. Densidade e Uso de Habitat de Psitacídeos na Área de Ocorrência Histórica da Extinta Ararinha-Azul; Centro de Ciências Agrárias (CCA)—Programa de Pós-Graduação em Biodiversidade: Areia, Brazil, 2016; p. 60. [Google Scholar]
- Nunes, M.F.C.; Galetti, M.; Marsden, S.; Pereira, R.S.; Peterson, A.T. Are large-scale distributional shifts of the Blue-winged Macaw (Primolius maracana) related to climate change? J. Biogeo. 2007, 34, 816–827. [Google Scholar] [CrossRef]
- Cavalcanti, L.C.S.; Rafael, L.M.; Barbosa, L.C.S.; Braz, A.M.; Ribeiro, J.R. Can landscape unit maps help the conservation of Spix’s Macaw (Cyanopsitta spixii)? Revista Ra’e Ga Espaço Geográfico Análise 2020, 8, 181. [Google Scholar] [CrossRef]
- Wilson, R.J.; Ellis, S.; Baker, J.S.; Lineham, M.E.; Whitehead, R.W.; Thomas, C.D. Large-scale patterns of distribution and persistence at the range margins of a butterfly. Ecology 2002, 83, 3357–3368. [Google Scholar] [CrossRef]
- Pichorim, M.; França Câmara, T.P.; Marques de Oliveira-Júnior, T.; Valdenor de Oliveira, D.; Galvão do Nascimento, E.P.; Mobley, J.A. A population of Blue-winged Macaw Primolius maracana in northeastern Brazil: Recommendations for a local Conservation Action Plan. Trop. Conserv. Sci. 2014, 7, 488–507. [Google Scholar] [CrossRef]
- Dias, D.M.; Massara, R.L.; Bocchiglieri, A. Use of habitats by donkeys and cattle within a protected area of the Caatinga dry forest biome in northeastern Brazil. Persp. in Ecol. Conserv. 2019, 17, 64–70. [Google Scholar] [CrossRef]
- Gomides, S.C.; Machado, T.M.; Evangelista-Vale, J.C.; Martins-Oliveira, A.T.; Pires-Oliveira, J.C.; Muller, A.; da Rosa, L.B.; Santos-Silva, D.L.; Eisenlohr, P.V. Assessing species reintroduction sites based on future climate suitability for food resources. Conserv. Biol. 2021, 35, 1821–1832. [Google Scholar] [CrossRef]
- Kanaan, V. Re-introduction of the Vinaceous-breasted Amazon at the Araucárias National Park, Santa Catarina, Brazil. In Global Re-Introduction Perspectives: 2016; Soorae, P.S., Ed.; Case-studies from around the globe; IUCN.SSC Reintroduction Specialist Group: Abu Dhabi, UAE, 2016; pp. 106–110. [Google Scholar]
- Martins, F.C.; Engel, M.T.; Schulz, F.; Martins, C.S.G. Human dimensions of the reintroduction of Brazilian birds. Front. Conserv. Sci. 2022, 3, 791103. [Google Scholar] [CrossRef]
- Barbosa, A.; Aécio, J.; Nóbrega, A.; Alves, N.; Romeu, R. Aspectos da caça e comércio ilegal da avifauna silvestre por populações tradicionais do semiárido paraibano. Rev. Biol. Ciên. Terra 2010, 10, 39–49. [Google Scholar]
- Ndenecho, E.N. Ecological planning and ecotourism development in Kimbi Game Reserve, Cameroon. J. Human Ecol. 2009, 27, 105–113. [Google Scholar]
- Estrada, A. Reintroduction of the Scarlet Macaw (Ara macao cyanoptera) in the Tropical Rainforests of Palenque, Mexico: Project Design and First Year Progress. Trop. Conserv. Sci. 2014, 7, 342–364. [Google Scholar] [CrossRef]
- Dayer, A.A.; Silva-Rodríguez, E.A.; Albert, S.; Chapman, M.; Zukowski, B.; Ibarra, J.T.; Gifford, G.; Echeverri, A.; Martínez-Salinas, A.; Sepúlveda-Luque, C. Applying conservation social science to study the human dimensions of Neotropical bird conservation. Condor 2020, 122, 1–15. [Google Scholar] [CrossRef]
- McKinley, D.C.; Miller-Rushing, A.J.; Ballard, H.L.; Bonney, R.; Brown, P.; Cook-Patton, S.C.; Evans, D.M.; French, R.A.; Parrish, J.K.; Phillips, T.B.; et al. Citizen science can improve conservation science, natural resource management, and environmental protection. Biol. Conserv. 2017, 208, 15–28. [Google Scholar] [CrossRef]
- Monroe, M.C. Two avenues for encouraging conservation behaviors. Human Ecol. 2003, 10, 113–125. [Google Scholar]
- Toomey, A.H.; Domrose, M.C. Can citizen science lead to positive conservation attitudes and behaviors? Res. Human Ecol. 2013, 20, 50–62. [Google Scholar]
- Carneiro, A.P.B.; Jimenez, J.E.; White, T.H., Jr.; Soto-Gamboa, M.R. Distribution of Slender-billed parakeets (Enicognathus leptorthynchus) in a fragmented agricultural landscape of southern Chile. Ornitol. Neotrop. 2012, 23, 201–213. [Google Scholar]
- White, T.H., Jr.; Jimenez, J.E. Lophozonia tree cavities used for nesting by Slender-billed Parakeets (Enicognathus leptorhynchus) in the central valley of southern Chile: A potentially vanishing keystone resource. Avian Res. 2017, 8, 3. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).