
Characteristics and Influencing Factors of Ecological Stoichiometry of Shrub Fine Roots in the Alpine Region of Northwest China


 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
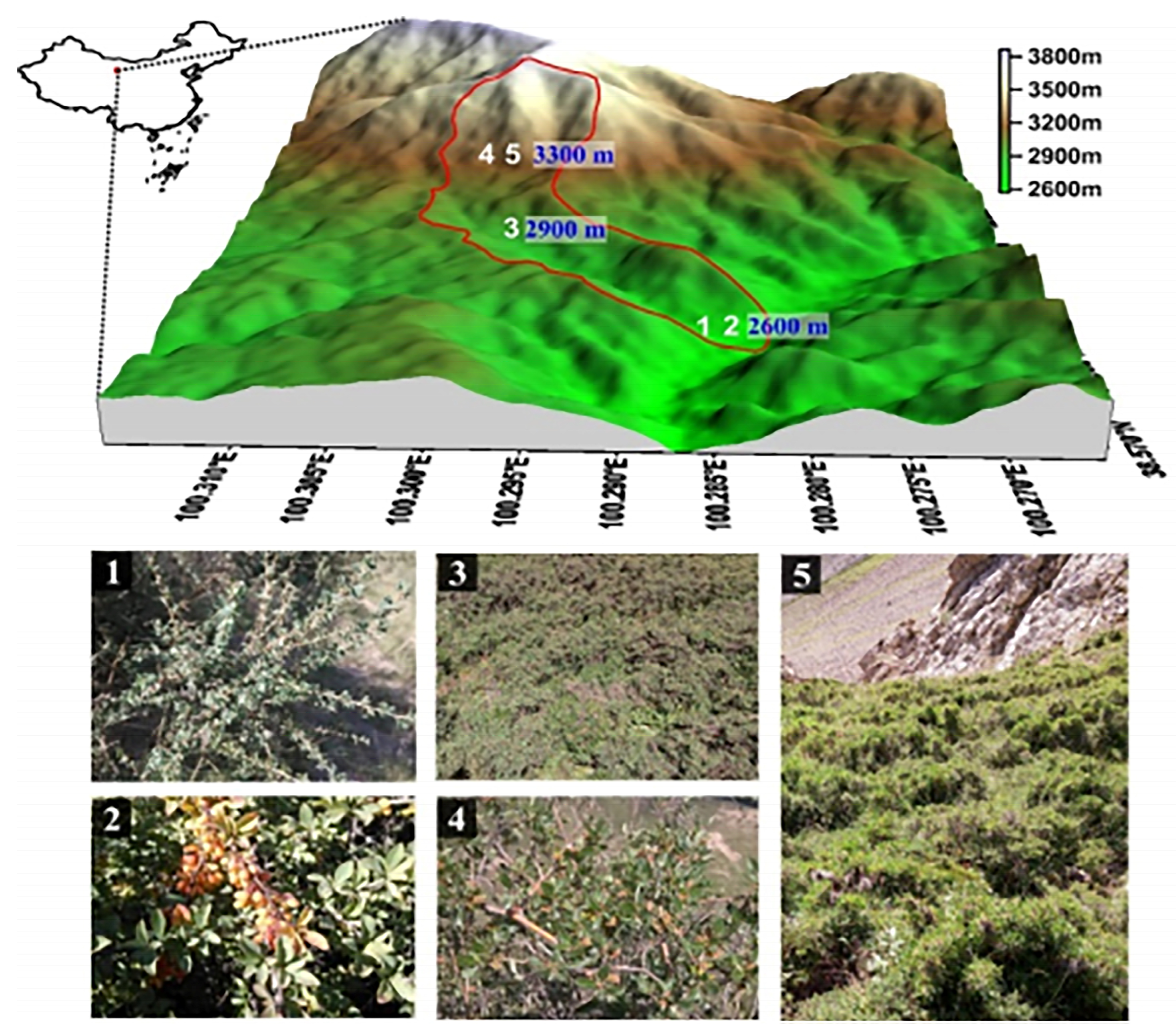
2.1. Sampling Areas
2.2. Sample Collection and Processing
2.3. Chemical Properties Analysis
2.4. Data Analysis
3. Results
3.1. Stoichiometric Characteristics of Shrub Fine Roots
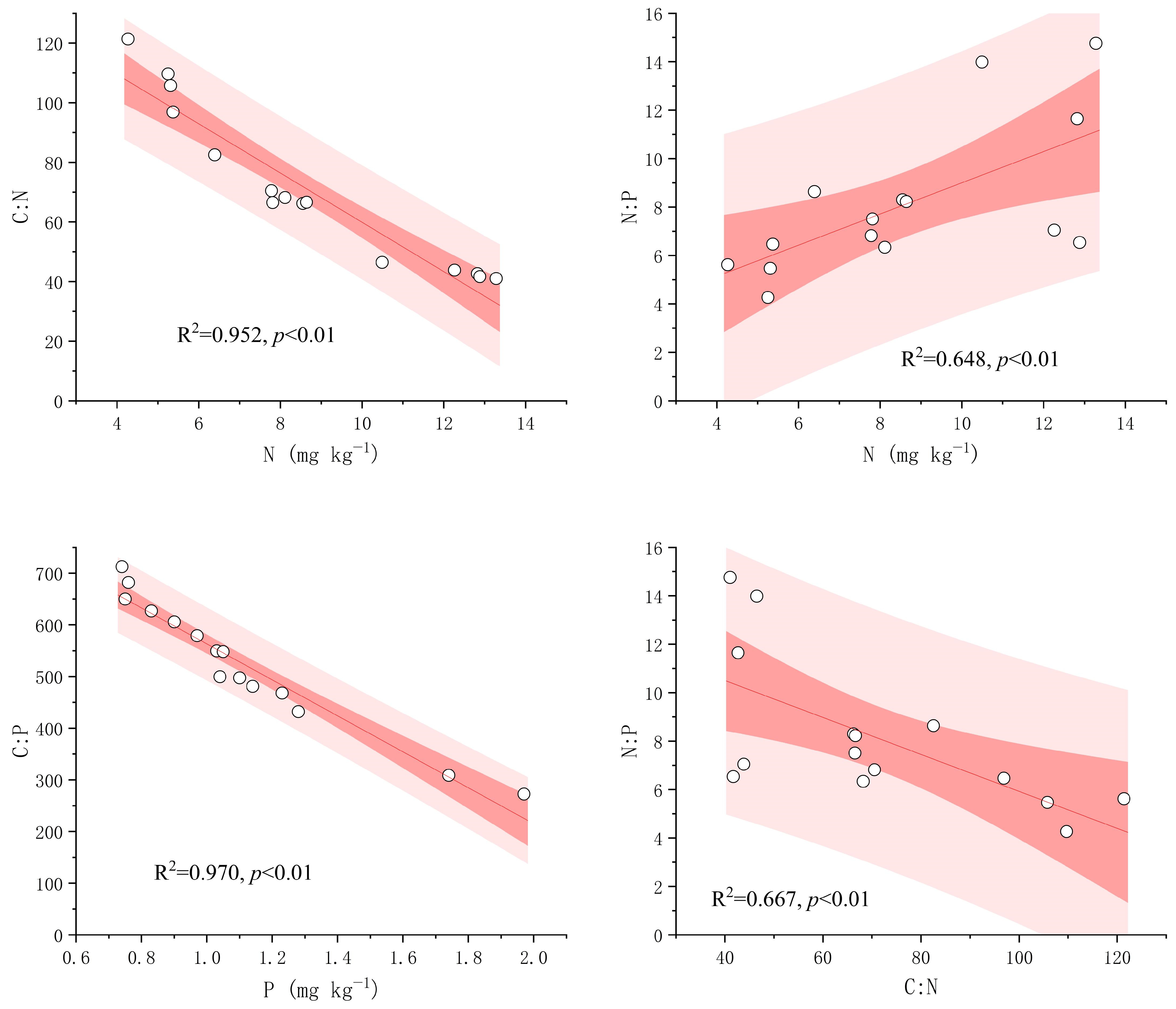
3.2. Correlations Between Fine-Root Stoichiometric Characteristics
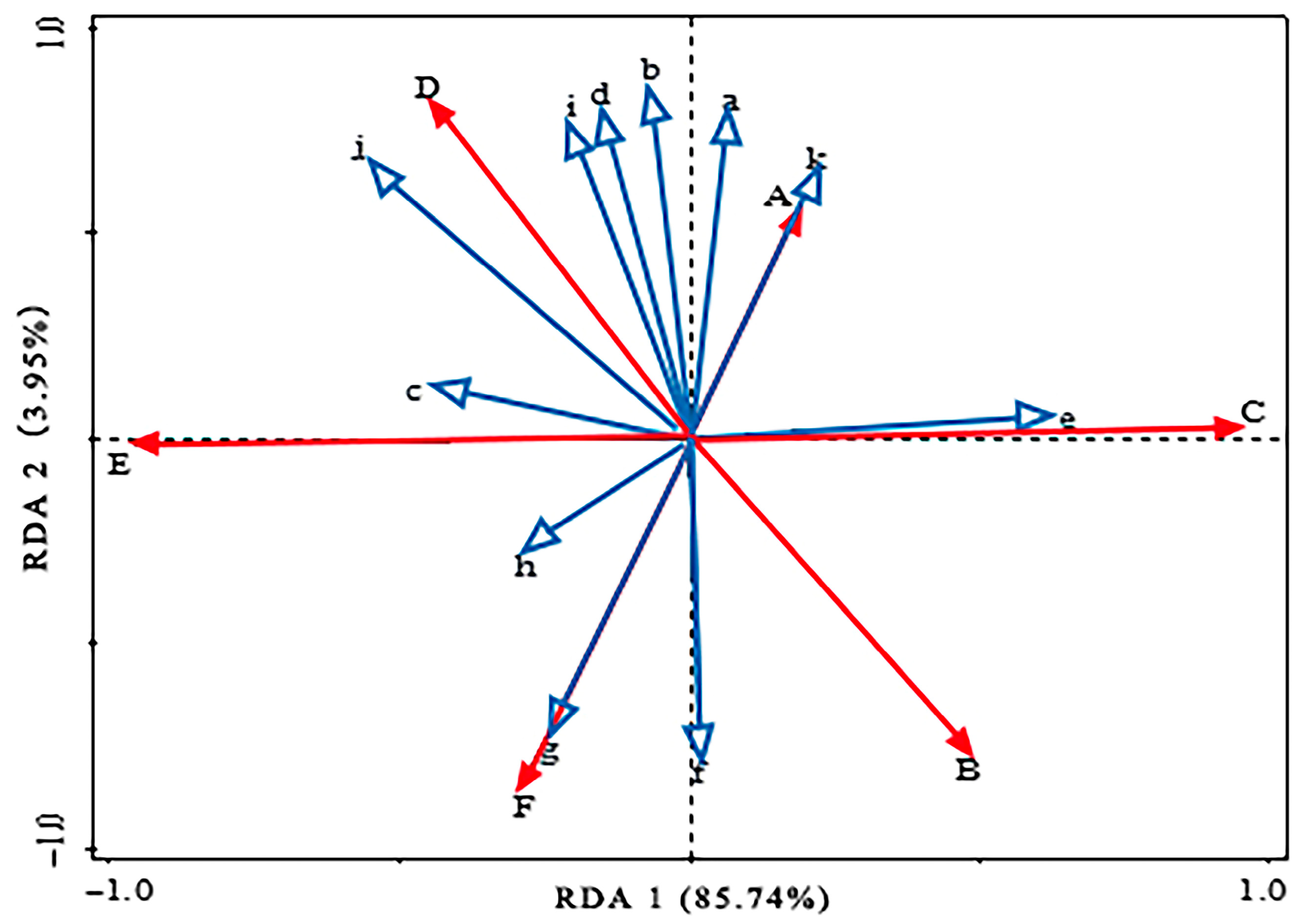
3.3. Relationships Between Fine-Root Stoichiometric Characteristics and Soil Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hessen, D.O. Stoichiometry in food webs: Lotka revisted. Oikos 1997, 79, 195–200. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, W.C.; Xu, M.P.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Response of forest growth to C:N: P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Platesu, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Qiu, X.C.; Wang, H.B.; Peng, D.L.; Liu, X.; Yang, F.; Li, Z.; Cheng, S. Thinning drives C:N: P stoichiometry and nutrient resorption in Larix principis-rupprechtii plantations in North China. For. Ecol. Manag. 2020, 462, 117984. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef]
- Vitouseck, P.M. Nutrient cycling and nutrient use efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Liu, R.; Wang, D. C: N: P stoichiometric characteristics and seasonal dynamics of leaf-root-litter-soil in plantations on the loess plateau. Ecol. Indic. 2021, 127, 107772. [Google Scholar] [CrossRef]
- Liu, Y.; He, J.W.; Yu, H.; Lin, Y.M.; Wang, D.J. Nutrients (C, N, P) contents and stoichiometric ratios of fine root, coarse root and leaf in dominant shrubs in dry-hot valley. Mt. Res. 2020, 38, 29–39. [Google Scholar]
- Zhang, H.; Gong, L.; Ding, Z.; Wu, X. Fine-root soil stoichiometry of picea schrenkiana fisch. et Mey. and its correlation with soil environmental factors under different nitrogen input levels in the Tianshan Mountains, Xinjiang. Forests 2023, 14, 426. [Google Scholar] [CrossRef]
- Heljä-Sisko, H.; Kirsi, M.; Seppo, K.; Esko, V.; Eino, M. Below and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. For. Ecol. Manag. 2002, 165, 317–326. [Google Scholar]
- Wu, J.S.; Shen, Z.X.; Zhang, X.Z.; Shi, P.L. Biomass allocation patterns of alpine grassland species and functional groups along a precipitation gradient on the Northern Tibetan Plateau. J. Mt. Sci. 2013, 10, 1097–1108. [Google Scholar] [CrossRef]
- Li, N.; Wang, G.X.; Yang, Y.; Gao, Y.H.; Liu, G.S. Plant production, and carbon and nitrogen source pools, are strongly intensified by experimental warming in alpine ecosystems in the Qinghai-Tibet Plateau. Soil Biol. Biochem. 2011, 43, 942–953. [Google Scholar]
- Qian, D.W.; Cao, G.M.; Du, Y.G.; Li, Q.; Guo, X.W. Impacts of climate change and human factors on land cover change in inland mountain protected areas: A case study of the Qilian Mountain National Nature Reserve in China. Environ. Monit. Assess. 2019, 191, 486. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Li, S.W.; Ran, Y.H.; Wang, H.W.; Wu, J.C.; Lian, X.H.; Luo, D.L. Mapping frozen ground in the Qilian Mountains in 2004–2019 using Google earth engine cloud computing. Remote Sens. 2021, 13, 149. [Google Scholar] [CrossRef]
- IUSS Working Group. World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014. [Google Scholar]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Commun. Soil Sci. Plant Anal. 2008, 19, 1467–1476. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 2001, 55, 11–33. [Google Scholar] [CrossRef]
- Ma, X.H. Research Methods of Forest Ecosystem Positioning; China Science and Technology Press: Beijing, China, 1994. (In Chinese) [Google Scholar]
- Braak, C.J.F.T.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’S Guide: Software for Canonical Community Ordination (Version 4.5); Section on Permutation Methods; Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Cao, Y.B.; Wang, B.T.; Wei, T.T.; Ma, H.H. Ecological stoiehiometric characteristics and element reserves of three stands in a closed forest on the Chinese loess plateau. Environ. Monit. Assess. 2016, 188, 80. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, E103–E122. [Google Scholar] [CrossRef]
- Yu, Q.; Elser, J.J.; He, N.P.; Wu, H.H.; Chen, Q.S.; Zhang, G.M.; Han, X.G. Stoichiometric homeostasis of vascular plants in the Inner Mongoliagrassland. Oecologia 2011, 66, 1–10. [Google Scholar] [CrossRef]
- Ågren, G.I. The C: N: P stoichiometry of autotrophs-theory and observations. Ecol. Lett. 2004, 7, 185–191. [Google Scholar] [CrossRef]
- Lasota, J.; Ewa, B. C: N: P stoichiometry as an indicator of Histosol drainage in lowland and mountain forest ecosystems. For. Ecosyst. 2021, 8, 39. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Sehulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Mayor, J.R.; Sanders, N.J.; Classen, A.T.; Bardgett, R.D.; Clément, J.C.; Fajardo, A.; Lavorel, S.; Sundqvist, M.K.; Bahn, M.; Chisholm, C. Elevation alters ecosystem properties across temperate treelines globally. Nature 2017, 542, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Schreeg, L.A.; Santiago, L.S.; Wright, S.J.; Turner, B.L. Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 2016, 95, 2062–2068. [Google Scholar] [CrossRef]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- He, X.L.; Ma, J.; Jin, M.; Li, Z. Characteristics and controls of ecological stoichiometry of shrub leaf in the alpine region of northwest China. Catena 2023, 224, 107005. [Google Scholar] [CrossRef]
- Franklin, O.; Ågren, G.I. Leaf senescence and resorption as mechanisms of maximizing photosynthetic production during canopy development at N limitation. Funct. Ecol. 2002, 16, 727–733. [Google Scholar] [CrossRef]
- Huang, D.; Wang, D.; Ren, Y. Using leaf nutrient stoichiometry as an indicator of flood tolerance and eutrophication in the riparian zone of the Lijang River. Ecol. Ind. 2019, 98, 821–829. [Google Scholar] [CrossRef]
- Dong, C.; Qiao, Y.; Cao, Y.; Chen, Y.; Wu, X.; Xue, W. Seasonal variations in carbon, nitrogen, and phosphorus stoichiometry of a Robinia pseudoacacia plantation on the Loess Hilly region, China. Forests 2021, 12, 214. [Google Scholar] [CrossRef]
- Yang, X.; Chi, X.; Ji, C.; Liu, H.; Ma, W.; Mohhammat, A. Variations of leaf N, P concentrations in shrubland biomes across northern China: Phylogeny, climate and soil. Biogeosciences 2016, 12, 18973–18998. [Google Scholar] [CrossRef]
- Hu, Y.F.; Shu, X.Y.; He, J.; Zhang, Y.L.; Xiao, H.H.; Tang, X.Y. Storage of C, N and P affected by afforestation with salix cupularis in an alpine semiarid desert ecosystem. Land Degrad. Dev. 2018, 2, 188–198. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shrubs | Soil Depth (cm) | Soil Type | Soil pH | Altitude (m) | Slope Gradient (°) | Slope Aspect (°) | Basal Diameter (mm) | Coverage (%) | Average Height (m) |
|---|---|---|---|---|---|---|---|---|---|
| CT | 60 | Chestnut soil | 8.5 | 2600 | 22 | Southwest | 25 | 50 | 1.4 |
| BD | 60 | Chestnut soil | 8.5 | 2600 | 30 | West | 20 | 70 | 1.8 |
| PF | 60 | Alpine meadow soil | 8.4 | 2900 | 33 | East | 16 | 90 | 0.9 |
| CJ | 60 | Alpine meadow soil | 7.7 | 3300 | 40 | Northwest | 20 | 60 | 0.6 |
| SG | 60 | Alpine meadow soil | 6.6 | 3300 | 32 | Northwest | 26 | 55 | 1.4 |
| Parameter | CT | BD | PF | CJ | SG | Mean | SD | CV | F Value | p Value |
|---|---|---|---|---|---|---|---|---|---|---|
| C (g kg−1) | 526.66 ± 33.85 a | 556.38 ± 25.55 a | 540.61 ± 17.72 a | 531.30 ± 10.20 a | 551.93 ± 29.95 a | 541.38 | 12.80 | 0.02 | 0.79 | 0.56 |
| N (g kg−1) | 12.20 ± 1.50 a | 7.86 ± 1.27 b | 7.09 ± 1.50 b | 10.98 ± 2.77 a | 4.94 ± 0.58 b | 8.61 | 2.64 | 0.31 | 9.27 | <0.01 |
| P (g kg−1) | 0.92 ± 0.18 b | 0.94 ± 0.17 b | 1.08 ± 0.23 ab | 1.58 ± 0.48 a | 0.99 ± 0.24 b | 1.10 | 0.25 | 0.22 | 2.84 | 0.04 |
| C:N | 43 ± 2.79 c | 71 ± 9.31 b | 78 ± 15.94 b | 50 ± 13.76 c | 112 ± 8.13 a | 71.34 | 24.23 | 0.34 | 18.22 | <0.001 |
| C:P | 584 ± 78.52 a | 603 ± 94.55 a | 513 ± 101.33 ab | 360 ± 121.94 b | 576 ± 106.96 a | 527.61 | 88.95 | 0.17 | 2.87 | 0.04 |
| N:P | 13 ± 1.62 a | 8 ± 0.22 b | 6 ± 0.25 cd | 7 ± 0.49 bc | 5 ± 0.74 d | 8.11 | 2.88 | 0.35 | 44.08 | <0.001 |
| Variables | MCR % | F-Ratio | p-Value |
|---|---|---|---|
| Soil AP | 33.2 | 6.4 | 0.024 * |
| Soil pH | 20.5 | 7.9 | 0.020 * |
| Soil total porosity | 17.6 | 4.3 | 0.080 |
| Soil bulk density | 6.4 | 2.8 | 0.130 |
| Soil C | 4.1 | 2.3 | 0.152 |
| Soil N | 4.0 | 2.0 | 0.164 |
| Soil noncapillary porosity | 2.0 | 0.7 | 0.438 |
| Soil P | 1.5 | 0.7 | 0.440 |
| Soil AN | 1.2 | 0.6 | 0.512 |
| Soil capillary porosity | 0.7 | 0.4 | 0.570 |
| Soil moisture content | 0.2 | <0.1 | 0.840 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Feng, Q.; Liu, W.; Chen, B.; Zhu, M.; Zhang, C.; Ta, F.; Tian, X.; Zhan, Y.; Li, X. Characteristics and Influencing Factors of Ecological Stoichiometry of Shrub Fine Roots in the Alpine Region of Northwest China. Diversity 2024, 16, 748. https://doi.org/10.3390/d16120748
Ma J, Feng Q, Liu W, Chen B, Zhu M, Zhang C, Ta F, Tian X, Zhan Y, Li X. Characteristics and Influencing Factors of Ecological Stoichiometry of Shrub Fine Roots in the Alpine Region of Northwest China. Diversity. 2024; 16(12):748. https://doi.org/10.3390/d16120748
Chicago/Turabian StyleMa, Jian, Qi Feng, Wei Liu, Bin Chen, Meng Zhu, Chengqi Zhang, Feng Ta, Xiaoping Tian, Yufang Zhan, and Xiaopeng Li. 2024. "Characteristics and Influencing Factors of Ecological Stoichiometry of Shrub Fine Roots in the Alpine Region of Northwest China" Diversity 16, no. 12: 748. https://doi.org/10.3390/d16120748
APA StyleMa, J., Feng, Q., Liu, W., Chen, B., Zhu, M., Zhang, C., Ta, F., Tian, X., Zhan, Y., & Li, X. (2024). Characteristics and Influencing Factors of Ecological Stoichiometry of Shrub Fine Roots in the Alpine Region of Northwest China. Diversity, 16(12), 748. https://doi.org/10.3390/d16120748