
A New Species of Cyprinid Genus Opsariichthys (Teleostei: Cyprinidae) from the Pearl River, Southern China †

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Storage
2.2. Morphological Description
2.3. Morphological Analysis
2.4. Molecular Phylogenetic Analysis
2.4.1. DNA Extraction, PCR Amplification, and Sequencing
2.4.2. Phylogenetic Analysis
3. Results
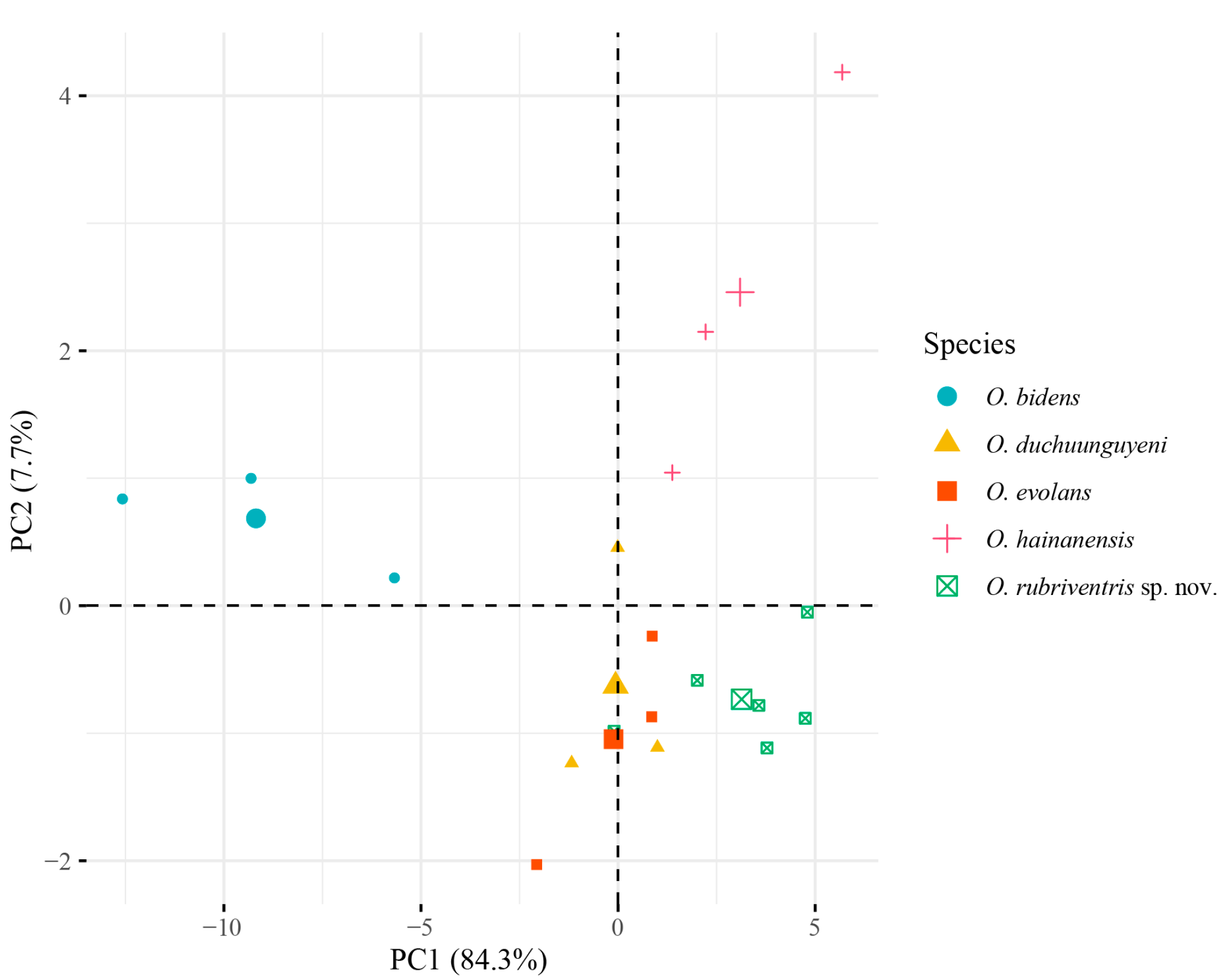
3.1. Morphological Analysis
3.2. Molecular Phylogenetic Analysis
3.3. Taxonomic Description
3.3.1. Diagnosis
3.3.2. Description
3.3.3. Fin
3.3.4. Scales
3.3.5. Coloration in Life
3.3.6. Head Structure
3.3.7. Etymology
3.3.8. Distribution and Habitat
4. Discussion
5. Comparative Materials
6. Nomenclatural Acts Registration
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Species | Locality | Source |
|---|---|---|---|
| PP975436 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975437 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975438 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975439 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975440 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975441 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975442 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975443 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975444 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975445 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975446 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975447 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975448 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975449 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975450 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975451 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975452 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975453 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975454 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975455 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975456 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975457 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975458 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975459 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975460 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975461 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975462 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975463 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975464 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PP975465 | Opsariichthys rubriventris sp. nov. | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| KJ940956 | Opsariichthys duchuunguyeni | Ky Cung—Bang Giang River basin, northern Vietnam | [9] |
| KJ940957 | Opsariichthys duchuunguyeni | Ky Cung—Bang Giang River basin, northern Vietnam | [9] |
| MZ507526 | Opsariichthys bidens | Balidian, Huzhou, Zhejiang, China | [36] |
| MZ507527 | Opsariichthys bidens | Balidian, Huzhou, Zhejiang, China | [36] |
| MZ507528 | Opsariichthys bidens | Balidian, Huzhou, Zhejiang, China | [36] |
| MZ507529 | Opsariichthys bidens | Balidian, Huzhou, Zhejiang, China | [36] |
| AY958197 | Opsariichthys uncirostris | L. Biwa, Shiga, Japan | [4] |
| AB218897 | Opsariichthys uncirostris | L. Biwa, Shiga, Japan | [37] |
| MH350625 | Opsariichthys evolans | Rong River, Fengshun, Guangdong, China | [38] |
| MH350626 | Opsariichthys evolans | Rong River, Fengshun, Guangdong, China | [38] |
| MH350627 | Opsariichthys evolans | Rong River, Fengshun, Guangdong, China | [38] |
| MH350628 | Opsariichthys evolans | Rong River, Fengshun, Guangdong, China | [38] |
| MH350629 | Opsariichthys evolans | Rong River, Fengshun, Guangdong, China | [38] |
| MH350630 | Opsariichthys evolans | Rong River, Fengshun, Guangdong, China | [38] |
| MH350779 | Opsariichthys macrolepis | Yangtze River, Zunyi, Guizhou, China | [38] |
| MH350780 | Opsariichthys macrolepis | Yangtze River, Zunyi, Guizhou, China | [38] |
| MH350781 | Opsariichthys macrolepis | Yangtze River, Zunyi, Guizhou, China | [38] |
| MH350782 | Opsariichthys macrolepis | Yangtze River, Zunyi, Guizhou, China | [38] |
| MH350784 | Zacco acanthogenys | Xin’an River, Huangshan, Anhui, China | [38] |
| MH350785 | Zacco acanthogenys | Xin’an River, Huangshan, Anhui, China | [38] |
| MH350786 | Zacco acanthogenys | Xin’an River, Huangshan, Anhui, China | [38] |
| MN325062 | Opsariichthys hainanensis | Changhua River, Qiongzhong County, Hainan, China | [38] |
| MN325063 | Opsariichthys hainanensis | Changhua River, Qiongzhong County, Hainan, China | [38] |
| MN325064 | Opsariichthys hainanensis | Changhua River, Qiongzhong County, Hainan, China | [38] |
| MN325065 | Opsariichthys hainanensis | Changhua River, Qiongzhong County, Hainan, China | [38] |
| MN325066 | Opsariichthys hainanensis | Changhua River, Qiongzhong County, Hainan, China | [38] |
| AB198972 | Zacco platypus | Yoshino River, Kochi, Japan | [39] |
| AB572372 | Zacco platypus | Naka River, Kanto Plain, Japan | [40] |
| AB572384 | Zacco platypus | Naka River, Kanto Plain, Japan | [40] |
| AB572406 | Zacco platypus | Naka River, Kanto Plain, Japan | [40] |
| AB572415 | Zacco platypus | Naka River, Kanto Plain, Japan | [40] |
| PQ283921 | Parazacco spilurus | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PQ283922 | Parazacco spilurus | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| PQ283923 | Parazacco spilurus | Xizhijiang River, Huizhou, Guangdong, China | This Study |
| NC025286 | Nipponocypris koreanus | Gwangcheon River, South Korea | [41] |
| NC015538 | Aphyocypris normalis | Unknown | [16] |
References
- Chen, Y.Y. A revision of opsariichthine cyprinid fishes. Oceanol. Limnol. Sin. 1982, 13, 293–299. [Google Scholar]
- Chen, I.S.; Chang, Y.C. The Photographic Guide of Inland Water Fishes of Taiwan; Sheichuan Press: Keelung, Taiwan, 2005; Volume I, Cypriniformes. [Google Scholar]
- Liao, N.L.; Huang, S.P.; Wang, T.Y. Interspecific Mating Behavior Between Introduced Zacco platypus and Native Opsariichthys evolans in Taiwan. Zool. Stud. 2020, 59, e6. [Google Scholar] [CrossRef]
- Huang, S.P.; Wang, F.Y.; Wang, T.Y. Molecular Phylogeny of the Opsariichthys Group (Teleostei: Cypriniformes) Based On Complete Mitochondrial Genomes. Zool. Stud. 2017, 56, e40. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, F.; Yu, D.; Liu, H. Mitochondrial divergence suggests unexpected high species diversity in the opsariichthine fishes (Teleostei: Cyprinidae) and the revalidation of Opsariichthys macrolepis. Ecol. Evol. 2019, 9, 2664–2677. [Google Scholar] [CrossRef] [PubMed]
- Perdices, A.; Sayanda, D.; Coelho, M.M. Mitochondrial diversity of Opsariichthys bidens (Teleostei, Cyprinidae) in three Chinese drainages. Mol. Phylogenet Evol. 2005, 37, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Jeon, S.R.; Kitagawa, T. Genetic differentiation of piscivorous chub (genus Opsariichthys) in Japan, Korea and Russia. Zool. Sci. 2002, 19, 601–610. [Google Scholar] [CrossRef]
- Chen, I.; Wu, J.; Huang, S. The taxonomy and phylogeny of the cyprinid genus Opsariichthys Bleeker (Teleostei: Cyprinidae) from Taiwan, with description of a new species. Environ. Biol. Fish. 2009, 86, 165–183. [Google Scholar] [CrossRef]
- Huynh, T.; Chen, I. A new species of cyprinid fish of genus Opsariichthys from Ky Cung—Bang Giang river basin, northern Vietnam with notes on the taxonomic status of the genus from northern Vietnam and southern China. J. Mar. Sci. Tech.-Jpn. 2013, 21, 135–145. [Google Scholar] [CrossRef]
- de Queiroz, K.; Gauthier, J. Phylogeny as a Central Principle in Taxonomy: Phylogenetic Definitions of Taxon Names. Syst. Biol. Biol. 1990, 39, 307–322. [Google Scholar] [CrossRef]
- Dunn, C.W.; Giribet, G.; Edgecombe, G.D.; Hejnol, A. Animal phylogeny and its evolutionary implications. Annu. Rev. Ecol. Evol. Syst. Rev. Ecol. Evol. Syst. 2014, 45, 371–395. [Google Scholar] [CrossRef]
- Briolay, J.; Galtier, N.; Brito, R.M.; Bouvet, Y. Molecular Phylogeny of Cyprinidae Inferred from cytochrome b DNA Sequences. Mol. Phylogenet Evol. 1998, 9, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Mesquita, N.; Dowling, T.E.; Gilles, A.; Coelho, M.M. Phylogenetic relationships of Eurasian and American cyprinids using cytochrome b sequences. J. Fish. Biol. 2002, 61, 929–944. [Google Scholar] [CrossRef]
- He, S.; Chen, Y.; Tsuneo, N. Sequences of cytochrome b gene for primitive cyprinid fishes in East Asia and their phylogenetic concerning. Chin. Sci. Bull. 2001, 46, 661–665. [Google Scholar] [CrossRef]
- Tang, K.L.; Agnew, M.K.; Hirt, M.V.; Lumbantobing, D.N.; Raley, M.E.; Sado, T.; Teoh, V.; Yang, L.; Bart, H.L.; Harris, P.M.; et al. Limits and phylogenetic relationships of East Asian fishes in the subfamily Oxygastrinae (Teleostei: Cypriniformes: Cyprinidae). Zootaxa 2013, 3681, 101–135. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.L.; Agnew, M.K.; Hirt, M.V.; Sado, T.; Schneider, L.M.; Freyhof, J.; Sulaiman, Z.; Swartz, E.; Vidthayanon, C.; Miya, M.; et al. Systematics of the subfamily Danioninae (Teleostei: Cypriniformes: Cyprinidae). Mol. Phylogenet Evol. 2010, 57, 189–214. [Google Scholar] [CrossRef]
- Chen, W.; Mayden, R.L. Molecular systematics of the Cyprinoidea (Teleostei: Cypriniformes), the world’s largest clade of freshwater fishes: Further evidence from six nuclear genes. Mol. Phylogenet Evol. 2009, 52, 544–549. [Google Scholar] [CrossRef]
- Zhu, L.; Yu, D.; Liu, H. Zacco sinensis sp. nov. (Cypriniformes: Cyprinidae), a New Fish Species from Northern China. Sichuan J. Zool. 2020, 39, 168–176. [Google Scholar] [CrossRef]
- Yan, Z.; Jiajun, Z.; Jinquan, Y. A new species of genus Zacco from Southern China (Cypriniformes: Cyprinidae). J. Shanghai Ocean Univ. 2023, 32, 544–552. [Google Scholar] [CrossRef]
- Lin, H.; Kuo, P.; Wang, W.; Chiu, Y.; Ju, Y.; Lin, F.; Hsu, K. Speciation and differentiation of the genus Opsariichthys (Teleostei: Cyprinidae) in East Asia. Biochem. Syst. Ecol. 2016, 68, 92–100. [Google Scholar] [CrossRef]
- George, D.; Mallery, P. IBM SPSS Statistics 26 Step by Step: A Simple Guide and Reference, 16th ed.; Routledge: London, UK, 2019. [Google Scholar] [CrossRef]
- Hosoya, K.; Ashiwa, H.; Watanabe, M.; Mizuguchi, K.; Okazaki, T. Zacco sieboldii, a species distinct from Zacco temminckii (Cyprinidae). Ichthyol. Res. 2003, 50, 1–8. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Principal component analysis. Wires Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. Msor Connect. 2014, 1. [Google Scholar]
- Sevilla, R.; Diez, A.; Norén, M.; Mouchel, O.; Jérôme, M.; Verrez-Bagnis, V.; Pelt, H.; Favre-Krey, L.; Krey, G.; Con-Sortium, T.; et al. Primers and polymerase chain reaction conditions for DNA barcoding teleost fish based on the mitochondrial cytochrome b and nuclear rhodopsin genes. Mol. Ecol. Notes 2007, 7, 730–734. [Google Scholar] [CrossRef]
- Alzohairy, A. BioEdit: An important software for molecular biology. Gerf Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Chen, W.; Li, C.; Chen, F.; Li, Y.; Yang, J.; Li, J.; Li, X. Phylogeographic analyses of a migratory freshwater fish (Megalobrama terminalis) reveal a shallow genetic structure and pronounced effects of sea-level changes. Gene 2020, 737, 144478. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Kottelat, M. Freshwater Fishes of Northern Vietnam. A Preliminary Check-list of the Fishes Known or Expected to Occur in Northern Vietnam with Comments on Systematics and Nomenclature. In Environment and Social Development Unit, East Asia and Pacific Region; The World Bank: Washington, DC, USA, 2001; pp. 1–73. [Google Scholar]
- Kottelat, M. Fishes of Laos; WHT Publications Ltd.: Colombo, Sri Lanka, 2001; p. 198. [Google Scholar]
- Amaoka, H.K.; Araga, C.; Uyeno, T.; Yoshino, T. The Fishes of the Japanese Archipelago; Tokai University Press: Tokyo, Japan, 1984; Volume 1. [Google Scholar]
- Seehausen, O.; Wagner, C.E. Speciation in Freshwater Fishes. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 621–651. [Google Scholar] [CrossRef]
- Seehausen, O.; Takimoto, G.; Roy, D.; Jokela, J. Speciation reversal and biodiversity dynamics with hybridization in changing environments. Mol. Ecol. 2008, 17, 30–44. [Google Scholar] [CrossRef]
- Barluenga, M.; Stölting, K.N.; Salzburger, W.; Muschick, M.; Meyer, A. Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature 2006, 439, 719–723. [Google Scholar] [CrossRef]
- Liu, S.; Lian, Q.; Jia, Y.; Chi, M.; Li, F.; Jiang, J.; Liu, Y.; Zheng, J.; Cheng, S.; Gu, Z. Genetic diversity analysis of three Opsariichthys bidens populations in Zhejiang Province based on mitochondrial Cyt b gene sequences. Acta Agric. Zhejiangensis 2023, 35, 293–300. [Google Scholar] [CrossRef]
- Saitoh, K.; Sado, T.; Mayden, R.L.; Hanzawa, N.; Nakamura, K.; Nishida, M.; Miya, M. Mitogenomic Evolution and Interrelationships of the Cypriniformes (Actinopterygii: Ostariophysi): The First Evidence Toward Resolution of Higher-Level Relationships of the World’s Largest Freshwater Fish Clade Based on 59 Whole Mitogenome Sequences. J. Mol. Evol. 2006, 63, 826–841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, J.; Li, C.; Chen, J.; Li, W.; Jiang, S.; Hsu, K.; Zhao, M.; Lin, H.; Zhao, J. Spatial genetic structure of Opsariichthys hainanensis in South China. Mitochondrial Dna A 2020, 31, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kartavtsev, Y.P.; Chiba, S.N.; Uematsu, T.; Sviridov, V.V.; Hanzawa, N. Genetic divergence and phylogenetic independence of Far Eastern species in subfamily Leuciscinae (Pisces: Cyprinidae) inferred from mitochondrial DNA analyses. Genes Genet. Syst. 2007, 82, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Takamura, K.; Nakahara, M. Intraspecific invasion occurring in geographically isolated populations of the Japanese cyprinid fish Zacco platypus. Limnology 2015, 16, 161–170. [Google Scholar] [CrossRef]
- Chen, I.; Liu, Y.; Huang, S.; Shen, C. The complete mitochondrial genome of the Korean minnow Nipponocypris koreanus (Cypriniformes, Cyprinidae). Mitochondrial Dna A 2016, 27, 708–710. [Google Scholar] [CrossRef]





| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 O. rubriventris sp. nov. | - | |||||||||||
| 2 O. duchuunguyeni | 0.16 | - | ||||||||||
| 3 O. evolans | 0.14 | 0.11 | - | |||||||||
| 4 O. bidens | 0.14 | 0.14 | 0.12 | - | ||||||||
| 5 O. uncirostris | 0.15 | 0.15 | 0.13 | 0.05 | - | |||||||
| 6 O. macrolepis | 0.14 | 0.11 | 0.06 | 0.12 | 0.13 | - | ||||||
| 7 O. hainanensis | 0.14 | 0.12 | 0.08 | 0.12 | 0.13 | 0.09 | - | |||||
| 8 N. koreanus | 0.15 | 0.17 | 0.15 | 0.15 | 0.16 | 0.15 | 0.15 | - | ||||
| 9 Z. platypus | 0.12 | 0.15 | 0.14 | 0.13 | 0.14 | 0.13 | 0.15 | 0.14 | - | |||
| 10 Z. acanthogenys | 0.13 | 0.16 | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 | 0.07 | - | ||
| 11 P. spilurus | 0.16 | 0.20 | 0.17 | 0.17 | 0.18 | 0.17 | 0.16 | 0.14 | 0.17 | 0.16 | - | |
| 12 A. normalis | 0.16 | 0.20 | 0.18 | 0.17 | 0.18 | 0.18 | 0.17 | 0.15 | 0.15 | 0.16 | 0.16 | - |
| Species | D iii | A iii | P1 i | P2 i | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 7 | M | 9 | 10 | 11 | M | 12 | 13 | 14 | 15 | M | 8 | 9 | M | ||||
| O. rubriventris sp. nov. | 9 | 7 | 4 | 3 | 2 | 10 | 1 | 7 | 1 | - | 13 | 8 | 1 | 8 | |||
| O. evolans | 5 | 7 | 5 | - | - | 9 | - | - | 4 | 1 | 14 | 5 | - | 8 | |||
| O. duchuunguyeni | 5 | 7 | 5 | - | - | 9 | - | - | 5 | - | 14 | 4 | 1 | 8 | |||
| O. bidens | 5 | 7 | 5 | - | - | 9 | - | 2 | 2 | 1 | 14 | 5 | - | 8 | |||
| O. hainanensis | 5 | 7 | 5 | - | - | 9 | - | 2 | 3 | - | 14 | 4 | 1 | 8 | |||
| Species | LL A | LL B | PreD | ||||||||||||||
| 7 | 8 | 10 | M | 3 | 4 | 5 | M | 13 | 14 | 15 | 16 | 19 | 20 | M | |||
| O. rubriventris sp. nov. | 3 | 6 | - | 8 | 9 | - | - | 3 | 5 | 4 | - | - | - | - | 13 | ||
| O. evolans | 1 | 4 | - | 8 | - | 5 | - | 4 | - | - | 2 | 3 | - | - | 16 | ||
| O. duchuunguyeni | 2 | 3 | - | 8 | 5 | - | - | 3 | 3 | 2 | - | - | - | - | 13 | ||
| O. bidens | - | 2 | 3 | 10 | 2 | 3 | - | 4 | - | - | - | 2 | 1 | 2 | 19 | ||
| O. hainanensis | - | 5 | - | 8 | 5 | - | - | 3 | - | - | 1 | 4 | - | - | 16 | ||
| Species | SCP | LL | |||||||||||||||
| 14 | 15 | 16 | 17 | 18 | 20 | M | 39 | 40 | 41 | 42 | 43 | 47 | 49 | 50 | M | ||
| O. rubriventris sp. nov. | 7 | 2 | - | - | - | - | 14 | 2 | 4 | - | 1 | 2 | - | - | - | 40 | |
| O. evolans | - | - | 5 | - | - | - | 16 | - | - | - | 2 | 3 | - | - | - | 43 | |
| O. duchuunguyeni | - | - | 3 | 2 | - | - | 16 | - | - | - | 2 | 3 | - | - | - | 43 | |
| O. bidens | - | - | 1 | 1 | 1 | 2 | 18 | - | - | - | 1 | 1 | 1 | 1 | 1 | 47 | |
| O. hainanensis | - | - | 5 | - | - | - | 16 | - | - | 3 | 1 | 1 | - | - | - | 41 | |
| O. rubriventris sp. nov. | O. evolans | O. duchuunguyeni | O. bidens | O. hainanensis | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Holotype | Male | Female | Male | Female | Male | Female | Male | Female | Male | Female | |||||||||||||||||||||
| N | 1 | 5 | 3 | 3 | 2 | 3 | 2 | 3 | 2 | 3 | 2 | ||||||||||||||||||||
| Standard Length (mm) | 67.9 | 64.4–81.7 | 54.3–60.6 | 76.4–92.9 | 69.3–69.8 | 78.9–84.9 | 62.2–66.8 | 108.2–156.9 | 96.6–98.6 | 76.4–92.9 | 69.3–69.8 | ||||||||||||||||||||
| % in SL | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | Max | Min | Mean ± SD | |
| Head length | 24.4 | 27.2 | 24.2 | 25.4 ± 1.4 | 28.6 | 26.2 | 27.0 ± 1.4 | 27.2 | 23.5 | 25.5 ± 1.8 | 26.8 | 25.6 | 26.2 ± 0.8 | 27.0 | 25.6 | 26.1 ± 0.8 | 26.3 | 25.8 | 26.0 ± 1.2 | 29.1 | 27.6 | 28.1 ± 0.8 | 29.7 | 28.1 | 28.9 ± 1.1 | 30.7 | 28.1 | 29.7 ± 1.8 | 33.5 | 27.7 | 30.6 ± 4.1 |
| Body depth | 32.9 | 32.9 | 26.5 | 28.8 ± 2.3 | 34.3 | 30.4 | 32.3 ± 1.9 | 30.1 | 27.8 | 29.2 ± 1.2 | 25.0 | 23.1 | 24 ± 1.4 | 29.1 | 24.4 | 26.5 ± 2.4 | 23.2 | 21.7 | 22.5 ± 1.1 | 28.4 | 20.6 | 24 ± 4.0 | 27.4 | 22.8 | 25.1 ± 3.2 | 24.0 | 21.1 | 22.8 ± 0.4 | 24.7 | 21.0 | 22.9 ± 2.6 |
| Predorsal length | 50.9 | 50.9 | 45.3 | 47.8 ± 1.9 | 50.7 | 46.1 | 48.5 ± 2.3 | 50.7 | 44.0 | 48.1 ± 3.6 | 50.7 | 49.2 | 50 ± 1.0 | 49.8 | 49.0 | 49.3 ± 0.4 | 50.5 | 49.0 | 49.8 ± 1 | 55.1 | 49.5 | 51.7 ± 3.0 | 52.6 | 51.3 | 52 ± 1.0 | 51.5 | 51.0 | 51.4 ± 0.3 | 52.5 | 50.9 | 51.7 ± 1.1 |
| Predorsal truncus length | 35.9 | 35.9 | 30.7 | 32.0 ± 2.0 | 32.5 | 29.3 | 31.0 ± 1.6 | 28.7 | 27.5 | 28.2 ± 0.6 | 29.1 | 28.7 | 28.9 ± 0.3 | 30.4 | 24.4 | 27 ± 3.1 | 23.2 | 21.7 | 22.5 ± 1.1 | 34.1 | 29.1 | 31.6 ± 2.5 | 32.6 | 32.3 | 32.5 ± 0.2 | 31.0 | 29.8 | 30.3 ± 0.9 | 30.2 | 28.6 | 29.4 ± 1.2 |
| Dorsal fin base length | 13.6 | 16.0 | 12.8 | 14.0 ± 1.1 | 11.8 | 10.8 | 11.2 ± 0.6 | 13.7 | 11.9 | 12.7 ± 0.9 | 12.9 | 10.8 | 11.9 ± 1.5 | 14.0 | 12.4 | 13 ± 0.8 | 11.2 | 10.3 | 10.8 ± 0.6 | 13.2 | 11.7 | 12.5 ± 0.8 | 11.0 | 10.9 | 11 ± 0.1 | 12.9 | 11.4 | 12.2 ± 0.5 | 11.4 | 11.0 | 11.2 ± 0.3 |
| Dorsal fin length | 22.3 | 24.5 | 20.5 | 22.6 ± 1.6 | 20.4 | 18.9 | 19.7 ± 0.8 | 24.3 | 23.5 | 24 ± 0.4 | 19.9 | 18.9 | 19.4 ± 0.7 | 23.1 | 19.4 | 21.7 ± 2.1 | 25.0 | 19.8 | 22.4 ± 3.7 | 23.6 | 22.6 | 23.1 ± 0.5 | 22.1 | 19.4 | 20.8 ± 1.9 | 24.5 | 18.5 | 22.1 ± 1.0 | 18.1 | 17.6 | 17.8 ± 0.4 |
| Preanal length | 61.4 | 67.6 | 61.4 | 65.3 ± 2.4 | 68.4 | 66.6 | 67.5 ± 0.9 | 70.2 | 62.7 | 67.1 ± 3.9 | 69.2 | 68.5 | 68.9 ± 0.5 | 70.8 | 63.2 | 66.6 ± 3.9 | 68.2 | 67.4 | 67.8 ± 0.6 | 72.7 | 68.4 | 71.2 ± 2.4 | 72.2 | 71.7 | 71.9 ± 0.4 | 74.3 | 68.9 | 71.2 ± 1.0 | 73.2 | 71.4 | 72.3 ± 1.3 |
| Anal fin base length | 16.9 | 18.5 | 16.0 | 17.4 ± 0.9 | 16.2 | 14.8 | 15.6 ± 0.7 | 18.4 | 13.9 | 15.9 ± 2.3 | 14.2 | 14.2 | 14.2 ± 0.0 | 19.4 | 14.5 | 17.5 ± 2.7 | 14.5 | 14.3 | 14.4 ± 0.1 | 15.3 | 13.5 | 14.3 ± 0.9 | 13.0 | 12.6 | 12.8 ± 0.3 | 15.3 | 11.8 | 14.1 ± 0.1 | 12.9 | 11.8 | 12.3 ± 0.8 |
| Anal fin length | 34.5 | 37.6 | 26.2 | 33.0 ± 4.5 | 21.8 | 17.6 | 19.2 ± 2.3 | 35.4 | 34.7 | 35.2 ± 0.4 | 30.5 | 29.0 | 29.7 ± 1.1 | 38.1 | 29.2 | 34.3 ± 4.6 | 22.6 | 19.9 | 21.3 ± 1.9 | 35.4 | 30.8 | 33.7 ± 2.5 | 25.2 | 23.5 | 24.4 ± 1.2 | 33.6 | 17.2 | 28.0 ± 0.3 | 20.6 | 19.3 | 19.9 ± 0.9 |
| Prepectoral length | 24.0 | 25.8 | 23.1 | 24.4 ± 1.2 | 25.1 | 24.7 | 25.0 ± 0.2 | 24.2 | 21.6 | 23.2 ± 1.3 | 26.3 | 25.4 | 25.8 ± 0.6 | 25.9 | 24.4 | 25.2 ± 0.8 | 25.3 | 25.1 | 25.2 ± 0.1 | 27.1 | 25.6 | 26.2 ± 0.8 | 26.3 | 26.2 | 26.2 ± 0 | 27.1 | 24.9 | 25.7 ± 0.2 | 28.8 | 28.3 | 28.5 ± 0.4 |
| Pectoral fin base length | 4.8 | 5.8 | 4.6 | 5.3 ± 0.5 | 6.1 | 5.1 | 5.6 ± 0.5 | 5.6 | 5.5 | 5.6 ± 0.1 | 5.5 | 3.8 | 4.6 ± 1.2 | 6.2 | 5.4 | 5.7 ± 0.5 | 4.4 | 4.2 | 4.3 ± 0.1 | 5.7 | 5.0 | 5.2 ± 0.4 | 4.6 | 4.5 | 4.6 ± 0.1 | 5.2 | 4.3 | 4.7 ± 0.3 | 4.5 | 4.2 | 4.3 ± 0.2 |
| Pectoral fin length | 22.3 | 25.3 | 19.8 | 22.6 ± 2.0 | 19.3 | 16.9 | 17.8 ± 1.3 | 25.1 | 24.1 | 24.8 ± 0.6 | 22.7 | 21.4 | 22 ± 0.9 | 25.8 | 23.7 | 24.7 ± 1.0 | 22.9 | 19.1 | 21 ± 2.7 | 22.7 | 14.5 | 18.7 ± 4.1 | 19.1 | 18.8 | 19 ± 0.2 | 25.5 | 16.8 | 20.1 ± 5.5 | 18.3 | 15.8 | 17.1 ± 1.7 |
| Prepelvic length | 44.4 | 49.1 | 44.4 | 46.9 ± 1.9 | 51.1 | 49.5 | 50.2 ± 0.8 | 71.5 | 45.3 | 55.3 ± 14.2 | 51.7 | 50.1 | 50.9 ± 1.1 | 51.2 | 47.5 | 48.7 ± 2.1 | 49.5 | 46.4 | 48 ± 2.2 | 52.7 | 51.0 | 51.7 ± 0.9 | 55.5 | 53.6 | 54.5 ± 1.4 | 53.8 | 50.7 | 51.9 ± 0.3 | 54.8 | 54.8 | 54.8 ± 0.0 |
| Pelvic fin base length | 5.1 | 5.7 | 4.4 | 5.1 ± 0.4 | 5.3 | 3.6 | 4.3 ± 0.9 | 4.8 | 4.2 | 4.4 ± 0.3 | 5.0 | 4.9 | 4.9 ± 0 | 4.6 | 3.8 | 4.4 ± 0.5 | 4.0 | 3.3 | 3.7 ± 0.5 | 4.3 | 3.8 | 4.1 ± 0.3 | 3.8 | 3.4 | 3.6 ± 0.3 | 3.8 | 2.8 | 3.4 ± 0.1 | 4.1 | 4.1 | 4.1 ± 0.0 |
| Pelvic fin length | 15.9 | 17.2 | 14.0 | 15.9 ± 1.2 | 13.4 | 12.7 | 13.0 ± 0.4 | 18.6 | 16.2 | 17.3 ± 1.2 | 16.0 | 13.3 | 14.7 ± 1.8 | 17.7 | 17.3 | 17.5 ± 0.2 | 16.4 | 13.1 | 14.8 ± 2.4 | 16.3 | 15.7 | 15.9 ± 0.4 | 14.5 | 13.7 | 14.1 ± 0.6 | 16.1 | 14.3 | 15 ± 1.2 | 13.6 | 13.1 | 13.4 ± 0.4 |
| Caudal peduncle length | 19.3 | 19.3 | 16.5 | 17.9 ± 1.1 | 22.1 | 17.8 | 20.1 ± 2.2 | 19.6 | 17.7 | 18.4 ± 1.1 | 18.7 | 18.6 | 18.7 ± 0.1 | 19.7 | 17.7 | 18.6 ± 1 | 22.8 | 18.8 | 20.8 ± 2.9 | 23.1 | 17.9 | 19.9 ± 2.8 | 20.5 | 19.6 | 20 ± 0.7 | 17.6 | 17.0 | 17.2 ± 0.0 | 18.4 | 17.9 | 18.2 ± 0.4 |
| Caudal peduncle depth | 9.1 | 10.8 | 9.1 | 10.2 ± 0.6 | 11.1 | 10.8 | 11.0 ± 0.1 | 11.1 | 10.2 | 10.6 ± 0.4 | 10.2 | 9.6 | 9.9 ± 0.4 | 10.3 | 9.3 | 9.6 ± 0.6 | 9.8 | 9.0 | 9.4 ± 0.5 | 9.4 | 8.5 | 9.0 ± 0.5 | 10.2 | 9.5 | 9.9 ± 0.5 | 10.4 | 8.6 | 9.5 ± 0.8 | 9.9 | 9.5 | 9.7 ± 0.3 |
| Body width in anal origin | 11.2 | 12.9 | 10.4 | 11.9 ± 0.9 | 12.1 | 10.7 | 12.4 ± 0.4 | 12.6 | 12.0 | 12.4 ± 0.4 | 10.9 | 10.0 | 10.4 ± 0.6 | 11.9 | 9.3 | 10.7 ± 1.3 | 9.0 | 7.6 | 8.3 ± 1.0 | 10.5 | 8.2 | 9.6 ± 1.2 | 10.0 | 9.7 | 9.8 ± 0.2 | 9.3 | 6.9 | 8.1 ± 0.9 | 9.3 | 7.8 | 8.5 ± 1.0 |
| % in HL | |||||||||||||||||||||||||||||||
| Head depth | 79.9 | 85.8 | 77.7 | 81.4 ± 2.7 | 86.3 | 72.7 | 78.9 ± 6.9 | 82.5 | 77.1 | 78.9 ± 3.1 | 77.1 | 72.4 | 74.7 ± 3.3 | 80.1 | 74.7 | 77.6 ± 2.7 | 72.0 | 66.3 | 69.2 ± 4.0 | 68.9 | 65.3 | 67.6 ± 2.0 | 67.9 | 61.4 | 64.6 ± 4.6 | 68.5 | 62.3 | 65.5 ± 4.4 | 68.7 | 60.4 | 64.5 ± 5.9 |
| Head width | 55.2 | 59.0 | 51.1 | 55.4 ± 2.6 | 57.1 | 55.6 | 56.2 ± 0.8 | 57.4 | 54.5 | 55.8 ± 1.5 | 49.7 | 49.1 | 49.4 ± 0.5 | 52.8 | 48.8 | 51.3 ± 2.2 | 50.5 | 46.0 | 48.2 ± 3.2 | 51.6 | 47.3 | 49.8 ± 2.2 | 53.3 | 46.4 | 49.9 ± 4.9 | 48.3 | 42.5 | 45.0 ± 4.1 | 47.1 | 40.5 | 43.8 ± 4.7 |
| Snout length | 32.6 | 34.5 | 28.4 | 31.1 ± 2.2 | 29.7 | 26.5 | 28.0 ± 1.6 | 30.6 | 30.5 | 30.5 ± 0 | 31.5 | 30.2 | 30.9 ± 0.9 | 35.3 | 30.2 | 32 ± 2.9 | 31.0 | 28.3 | 29.7 ± 2.0 | 33.8 | 31.3 | 32.2 ± 1.4 | 35.7 | 32.0 | 33.9 ± 2.6 | 32.6 | 28.6 | 30.7 ± 2.8 | 36.8 | 33.8 | 35.3 ± 2.1 |
| Eye diameter | 27.6 | 32.1 | 27.3 | 29.4 ± 1.9 | 33.9 | 30.3 | 31.6 ± 2.0 | 29.3 | 28.6 | 28.9 ± 0.4 | 27.4 | 26.0 | 26.7 ± 1.0 | 29.5 | 26.3 | 27.6 ± 1.7 | 31.3 | 30.3 | 30.8 ± 0.7 | 22.5 | 15.2 | 18 ± 3.9 | 20.7 | 18.7 | 19.7 ± 1.4 | 25.8 | 24.5 | 24.9 ± 0.9 | 22.5 | 22.1 | 22.3 ± 0.3 |
| Interorbital width | 37.6 | 39.1 | 35.6 | 37.2 ± 1.2 | 37.5 | 33.9 | 35.2 ± 2.0 | 36.5 | 33.0 | 35.3 ± 2 | 38.1 | 34.6 | 36.3 ± 2.5 | 34.1 | 33.2 | 33.5 ± 0.5 | 33.3 | 30.8 | 32.1 ± 1.8 | 35.1 | 29.6 | 32.2 ± 2.7 | 35.2 | 30.0 | 32.6 ± 3.7 | 30.2 | 25.9 | 28.5 ± 2.5 | 32.8 | 28.0 | 30.4 ± 3.4 |
| Upper jaw length | 38.1 | 41.7 | 38.1 | 39.5 ± 1.3 | 41.3 | 36.2 | 38.7 ± 2.5 | 39.0 | 38.5 | 38.7 ± 0.2 | 32.7 | 23.4 | 28 ± 6.6 | 40.2 | 36.2 | 38.8 ± 2.3 | 41.0 | 38.0 | 39.5 ± 2.2 | 51.5 | 41.5 | 46.6 ± 5 | 47.0 | 42.8 | 44.9 ± 3.0 | 49.8 | 47.0 | 48.2 ± 1.6 | 52.2 | 47.3 | 49.8 ± 3.4 |
| Lower jaw length | 37.6 | 41.3 | 37.3 | 38.7 ± 1.6 | 39.1 | 36.1 | 37.5 ± 1.5 | 38.3 | 36.9 | 37.8 ± 0.8 | 37.1 | 32.3 | 34.7 ± 3.4 | 39.3 | 35.3 | 37.1 ± 2.0 | 39.0 | 37.8 | 38.4 ± 0.8 | 50.6 | 42.8 | 46.8 ± 3.9 | 48.8 | 46.1 | 47.4 ± 1.9 | 50.7 | 48.9 | 49.6 ± 1.3 | 51.3 | 50.9 | 51.1 ± 0.3 |
| Mouth width | 26.2 | 35.7 | 26.2 | 30.6 ± 3.5 | 29.9 | 26.9 | 28.0 ± 1.6 | 29.0 | 25.7 | 26.9 ± 1.9 | 23.3 | 20.8 | 22.1 ± 1.8 | 31.8 | 26.3 | 28.9 ± 2.7 | 28.3 | 24.6 | 26.5 ± 2.6 | 36.0 | 27.0 | 32.2 ± 4.7 | 30.4 | 26.5 | 28.5 ± 2.7 | 27.3 | 23.4 | 25.6 ± 2.0 | 30.7 | 22.6 | 26.6 ± 5.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.-B.; Li, Y.-T.; Zhou, J.-J.; Li, C.; Weng, G.-X.; Lin, H.-D.; Wang, J.-J. A New Species of Cyprinid Genus Opsariichthys (Teleostei: Cyprinidae) from the Pearl River, Southern China †. Diversity 2024, 16, 596. https://doi.org/10.3390/d16100596
Chen J-B, Li Y-T, Zhou J-J, Li C, Weng G-X, Lin H-D, Wang J-J. A New Species of Cyprinid Genus Opsariichthys (Teleostei: Cyprinidae) from the Pearl River, Southern China †. Diversity. 2024; 16(10):596. https://doi.org/10.3390/d16100596
Chicago/Turabian StyleChen, Jia-Bo, Ying-Tao Li, Jia-Jun Zhou, Cheng Li, Guo-Xi Weng, Hung-Du Lin, and Jun-Jie Wang. 2024. "A New Species of Cyprinid Genus Opsariichthys (Teleostei: Cyprinidae) from the Pearl River, Southern China †" Diversity 16, no. 10: 596. https://doi.org/10.3390/d16100596
APA StyleChen, J.-B., Li, Y.-T., Zhou, J.-J., Li, C., Weng, G.-X., Lin, H.-D., & Wang, J.-J. (2024). A New Species of Cyprinid Genus Opsariichthys (Teleostei: Cyprinidae) from the Pearl River, Southern China †. Diversity, 16(10), 596. https://doi.org/10.3390/d16100596