
The Mitochondrial Genome of Littoraria melanostoma Reveals a Phylogenetic Relationship within Littorinimorpha


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Sequencing, Assembling and Analysis
2.3. Phylogenetic Analysis
3. Results and Discussion
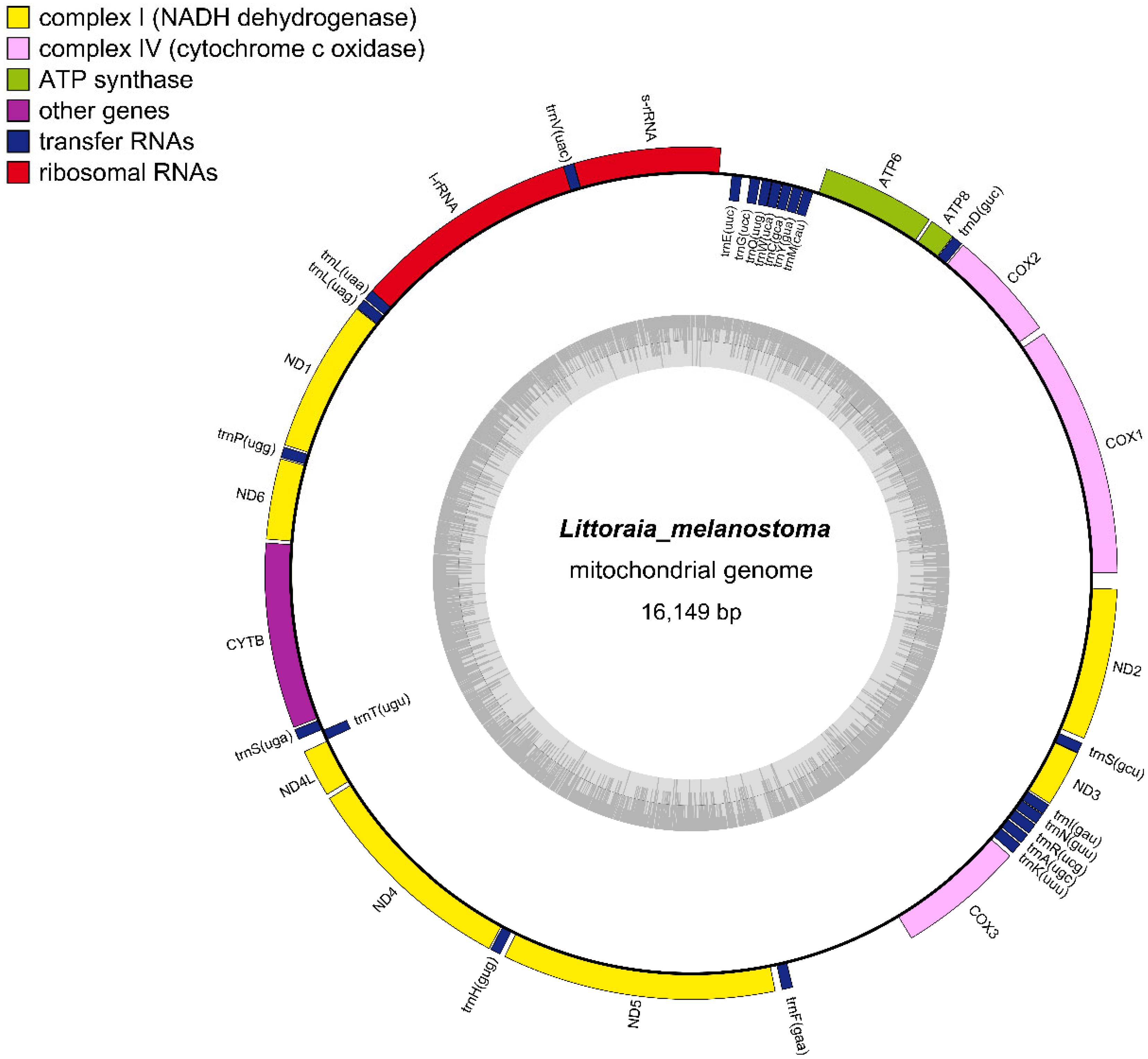
3.1. Genome Structure and Organization
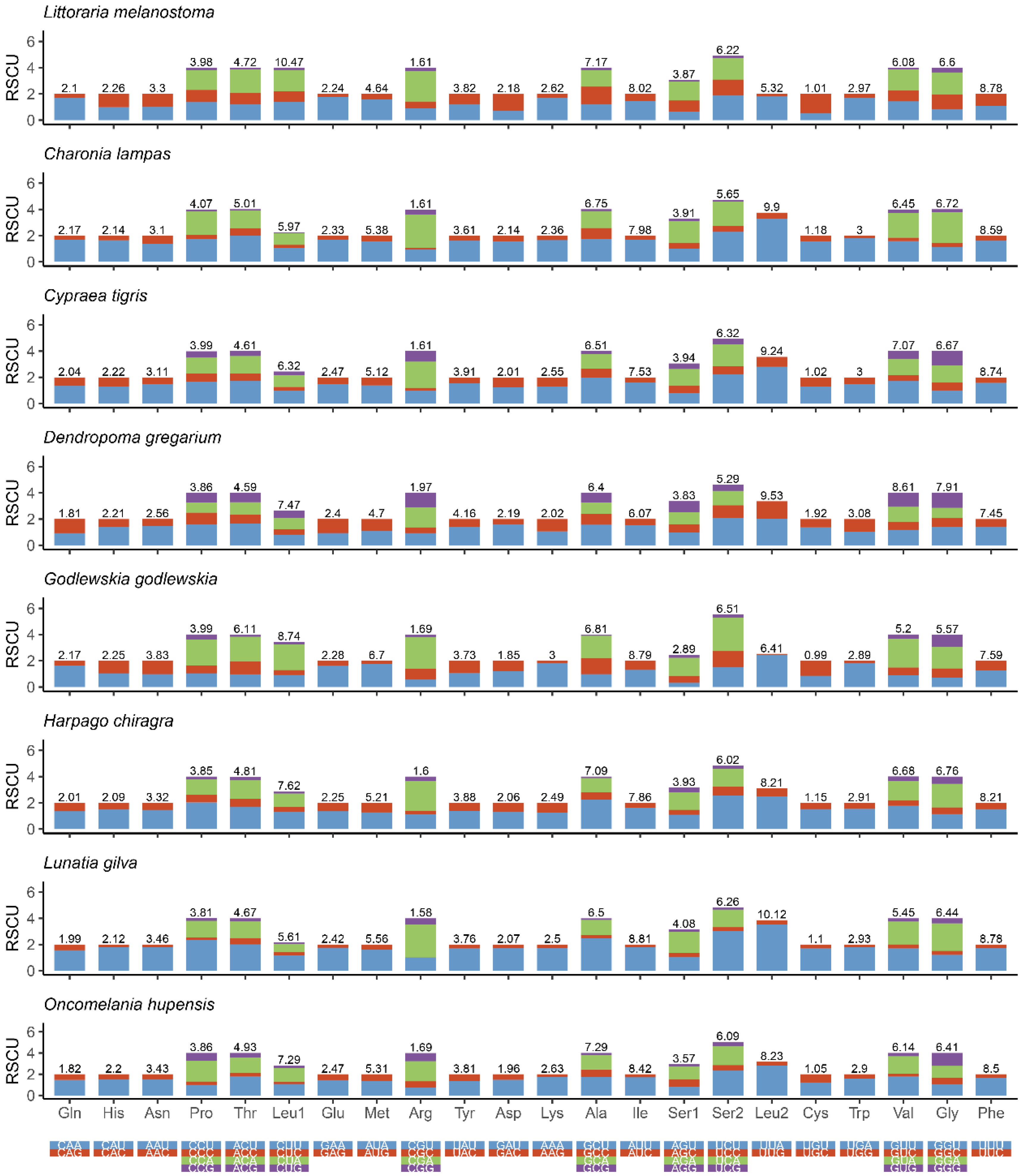
3.2. PCGs and Codon Usage
3.3. Ribosomal and Transfer RNA Genes
3.4. Intergenic Spaces and Overlapping Sequences
3.5. Control Regions
3.6. Phylogenetic Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torres, P.; Alfiado, A.; Glassom, D.; Jiddawi, N.; Macia, A.; Reid, D.G.; Paula, J. Species composition, comparative size and abundance of the genus Littoraria (Gastropoda: Littorinidae) from different mangrove strata along the East African coast. Hydrobiologia 2008, 614, 339–351. [Google Scholar] [CrossRef]
- Berry, A.; Chew, E. Reproductive systems and cyclic release of eggs in Littorina melanostoma from Malayan mangrove swamps (Mollusca: Gastropoda). J. Zool. 1973, 171, 333–344. [Google Scholar] [CrossRef]
- Lee, O.H.; Williams, G.A.; Hyde, K.D. The diets of Littoraria ardouiniana and L. melanostoma in Hong Kong mangroves. J. Mar. Biol. Assoc. UK 2001, 81, 967–973. [Google Scholar] [CrossRef]
- Hamilton, P. Intertidal distribution and long-term movements of Littorina irrorata (Mollusca: Gastropoda). Mar. Biol. 1978, 46, 49–58. [Google Scholar] [CrossRef]
- Alfaro, A.C. Diet of Littoraria scabra, while vertically migrating on mangrove trees: Gut content, fatty acid, and stable isotope analyses. Estuar. Coast. Shelf Sci. 2008, 79, 718–726. [Google Scholar] [CrossRef]
- Little, C.; Stirling, P. Activation of a mangrove snail, Littorina scabra scabra (L.) (Gastropoda: Prosobranchia). Mar. Freshwater Res. 1984, 35, 607–610. [Google Scholar] [CrossRef]
- Cook, L. Systematic effects on morph frequency in the polymorphic mangrove snail Littoraria pallescens. Heredity 1990, 65, 423–427. [Google Scholar] [CrossRef]
- Lee, O.H.; Williams, G.A. Spatial distribution patterns of Littoraria species in Hong Kong mangroves. Hydrobiologia 2002, 481, 137–145. [Google Scholar] [CrossRef]
- Blanco, J.F.; Cantera, J.R. The vertical distribution of mangrove gastropods and environmental factors relative to tide level at Buenaventura Bay, Pacific Coast of Colombia. Bull. Mar. Sci. 1999, 65, 617–630. [Google Scholar]
- Reid, D.G. Habitat and zonation patterns of Littoraria species (Gastropoda: Littorinidae) in Indo-Pacific mangrove forests. Biol. J. Linn. Soc. 1985, 26, 39–68. [Google Scholar] [CrossRef]
- Chen, S.Y.; Chen, B.; Liao, J.J.; Chen, G.L.; Huang, Y.; Chen, G.C. Composition and distribution pattern of Littorinid snails in young rehabilitated mangroves. Chin. J. Ecol. 2017, 36, 460. [Google Scholar]
- Reid, D.G.; Dyal, P.; Williams, S.T. A global molecular phylogeny of 147 periwinkle species (Gastropoda, Littorininae). Zool. Scr. 2012, 41, 125–136. [Google Scholar] [CrossRef]
- Lai, T.H.; He, B.Y. Studies on the macrobenthos species diversity for Guangxi mangrove areas. Guangxi Sci. 1998, 5, 166–172. [Google Scholar]
- Chen, G.; Ye, Y. Restoration of Aegiceras corniculatum mangroves in Jiulongjiang Estuary changed macro-benthic faunal community. Ecol. Eng. 2011, 37, 224–228. [Google Scholar] [CrossRef]
- Tang, C.Y. Vertical Distribution Patterns of Mangrove Arboreal Mollusks and Its Influencing Factors. Master’s Thesis, Xiamen University, Xiamen, China, 2019. [Google Scholar]
- Reid, D.G.; Dyal, P.; Williams, S. Global diversification of mangrove fauna: A molecular phylogeny of Littoraria (Gastropoda: Littorinidae). Mol. Phylogenet Evol. 2010, 55, 185–201. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Li, Y.L.; Xing, T.F.; Liu, J.X. First mitochondrial genome of a periwinkle from the genus Littoraria: Littoraria sinensis. Mitochondr. DNA 2019, 4, 4124–4125. [Google Scholar] [CrossRef] [PubMed]
- Irwin, A.R.; Strong, E.E.; Kano, Y.; Harper, E.M.; Williams, S.T. Eight new mitogenomes clarify the phylogenetic relationships of Stromboidea within the caenogastropod phylogenetic framework. Mol. Phylogenet Evol. 2021, 158, 107081. [Google Scholar] [CrossRef]
- Lee, W.K.; Hou, B.K.; Ju, S.J.; Kim, S.J. Complete mitochondrial genome of the hydrothermal vent provannid snail Alviniconcha boucheti (Gastropoda: Abyssochrysoidea) from the North Fiji Basin. Mitochondr. DNA 2020, 5, 1848–1849. [Google Scholar] [CrossRef]
- Xu, T.; Sun, J.; Chen, C.; Qian, P.Y.; Qiu, J.W. The mitochondrial genome of the deep-sea snail Provanna sp. (Gastropoda: Provannidae). Mitochondr. DNA 2016, 27, 4026–4027. [Google Scholar] [CrossRef]
- MolluscaBase. 2023. Available online: https://www.molluscabase.org (accessed on 5 September 2023). [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—A suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575–W581. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.P.; Sotelo, G.; Larsson, T.; Johannesson, K.; Panova, M.; Faria, R. Comparative mitogenomic analysis of three species of periwinkles: Littorina fabalis, L. obtusata and L. saxatilis. Mar. Genom. 2017, 32, 41–47. [Google Scholar] [CrossRef]
- Song, W.T.; Gao, X.G.; Li, Y.F.; Liu, W.D.; Liu, Y.; He, C.B. Comparison of mitochondrial genomes of bivalves. Hereditas 2009, 31, 1127–1134. [Google Scholar] [CrossRef]
- Hoffmann, R.J.; Boore, J.; Brown, W. A novel mitochondrial genome organization for the blue mussel, Mytilus Edulis. Genetics 1992, 131, 397–412. [Google Scholar] [CrossRef]
- Jiang, D.; Zheng, X.; Zeng, X.; Kong, L.; Li, Q. The complete mitochondrial genome of Harpago chiragra and Lambis lambis (Gastropoda: Stromboidea): Implications on the Littorinimorpha phylogeny. Sci. Rep. 2019, 9, 17683. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Hwang, U.W. The complete mitochondrial genome of an endangered triton snail Charonia lampas (Littorinimorpha: Charoniidae) from South Korea. Mitochondr. DNA 2021, 6, 956–958. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomy Superfamily | Species | bp | Accession Number |
|---|---|---|---|
| Littorinoidea | Littoraria melanostoma (Gray, 1839) | 16,149 | NC064398 |
| Littoraria ardouiniana (Heude, 1885) | 16,261 | ON920192 | |
| Littoraria sinensis (R. A. Philippi, 1847) | 16,420 | MN496138 | |
| Littorina brevicula (R. A. Philippi, 1844) | 16,356 | MT362562 | |
| Littorina saxatilis (Olivi, 1792) | 16,887 | KU952094 | |
| Littorina fabalis (W. Turton, 1825) | 16,418 | KU952092 | |
| Melarhaphe neritoides (Linnaeus, 1758) | 15,676 | MH119311 | |
| Stromboidea | Harpago chiragra (Linnaeus, 1758) | 16,404 | MN885884 |
| Lambis lambis (Linnaeus, 1758) | 15,481 | MH115428 | |
| Conomurex luhuanus (Linnaeus, 1758) | 15,799 | KY853669 | |
| Tibia fusus (Linnaeus, 1758) | 16,083 | MZ359282 | |
| Xenophora japonica Kuroda & Habe, 1971 | 15,684 | MW244823 | |
| Laevistrombus canarium (Linnaeus, 1758) | 15,626 | MT937083 | |
| Varicospira cancellate (Lamarck, 1816) | 15,864 | MW244822 | |
| Tridentarius dentatus (Linnaeus, 1758) | 15,500 | MW244820 | |
| Terebellum terebellum (Linnaeus, 1758) | 15,478 | MW244821 | |
| Struthiolaria papulose (Martyn, 1784) | 15,475 | MW244818 | |
| Aporrhais serresiana (Michaud, 1828) | 15,455 | MW244817 | |
| Ministrombus variabilis (Swainson, 1820) | 15,292 | MW244824 | |
| Onustus exutus (Reeve, 1842) | 16,043 | MK327366 | |
| Cypraeoidea | Cypraea tigris Linnaeus, 1758 | 16,177 | MK783263 |
| Purpuradusta gracilis (Gaskoin, 1849) | 16,240 | OP723877 | |
| Mauritia arabica (Linnaeus, 1758) | 16,926 | MZ667219 | |
| Lyncina vitellus (Linnaeus, 1758) | 16,269 | OP714183 | |
| Naria erosa (Linnaeus, 1758) | 16,020 | OP738004 | |
| Erronea caurica (Linnaeus, 1758) | 15,857 | OP714186 | |
| Monetaria annulus (Linnaeus, 1758) | 16,087 | LC469295 | |
| Naticoidea | Paratectonatica tigrine (Röding, 1798) | 16,148 | MN419027 |
| Naticarius hebraeus (Martyn, 1786) | 15,384 | KP716634 | |
| Polinices sagamiensis Pilsbry, 1904 | 15,383 | MK478018 | |
| Mammilla mammata (Röding, 1798) | 15,319 | MK433194 | |
| Cryptonatica andoi (Nomura, 1935) | 15,302 | MK433195 | |
| Euspira gilva (R. A. Philippi, 1851) | 16,119 | MN419026 | |
| Neverita didyma (Röding, 1798) | 15,252 | MK548644 | |
| Notocochlis gualteriana (Récluz, 1844) | 15,176 | MK500872 | |
| Glossaulax reiniana (Dunker, 1877) | 15,254 | MH543334 | |
| Tanea lineata (Röding, 1798) | 15,156 | MK507894 | |
| Vermetoidea | Dendropoma gregarium M. G. Hadfield & Kay, 1972 | 15,641 | HM174252 |
| Ceraesignum maximum (G. B. Sowerby I, 1825) | 15,578 | HM174253 | |
| Thylacodes squamigerus (P. P. Carpenter, 1857) | 15,544 | HM174255 | |
| Eualetes tulipa (Rousseau, 1843) | 15,078 | NC_014585 | |
| Tonnoidea | Charonia lampas (Linnaeus, 1758) | 15,405 | MG181942 |
| Tonna galea (Linnaeus, 1758) | 17,504 | OR282483 | |
| Ficus variegate Röding, 1798 | 15,736 | MW376482 | |
| Bursa rhodostoma (G. B. Sowerby II, 1835) # | 15,393 | MW316791 | |
| Tutufa rubeta (Linnaeus, 1758) | 15,397 | MW316790 | |
| Galeodea echinophora (Linnaeus, 1758) | 15,388 | KP716635 | |
| Monoplex parthenopeus (Salis Marschlins, 1793) | 15,270 | EU827200 | |
| Truncatelloidea | Oncomelania hupensis Gredler, 1881 | 15,191 | EU079378 |
| Stenothyra glabra A. Adams, 1861 | 15,704 | MZ196218 | |
| Tricula hortensis Attwood & D. S. Brown, 2003 | 15,179 | EU440735 | |
| Potamopyrgus antipodarum (J. E. Gray, 1843) | 16,846 | OQ161206 | |
| Bithynia leachii (Sheppard, 1823) | 15,682 | MT410857 | |
| Godlewskia godlewskii (W. Dybowski, 1875) | 15,224 | KY697387 | |
| Baicalia turriformis (W. Dybowski, 1875) | 15,127 | KY697386 | |
| Korotnewia korotnewi (Lindholm, 1909) ## | 15,171 | KY697389 | |
| Maackia herderiana (Lindholm, 1909) | 15,154 | KY697388 | |
| Calyptraeoidea | Desmaulus extinctorium (Lamarck, 1822) | 16,608 | OQ511529 |
| Abyssochrysoidea | Alviniconcha adamantis S. B. Johnson, Warén, Tunnicliffe, Van Dover, Wheat, T. F. Schultz & Vrijenhoek, 2014 | 16,342 | OL351262 |
| Desbruyeresia armata C. Chen, H. K. Watanabe & Sasaki, 2019 | 15,825 | OM063153 | |
| Provanna clathrate Sasaki, Ogura, H. K. Watanabe & Fujikura, 2016 | 15,344 | OK632464 | |
| Ifremeria nautilei Bouchet & Warén, 1991 | 15,664 | KC757644 |
| Littoraria melanostoma | ||||||||
|---|---|---|---|---|---|---|---|---|
| A% | T% | G% | C% | AT% | GC% | AT Skew | GC Skew | |
| Mitogenome | 29.79 | 34.37 | 14.66 | 21.18 | 64.16 | 35.84 | −0.071 | −0.182 |
| All PCGs | 27.77 | 34.62 | 15.15 | 22.46 | 62.39 | 37.61 | −0.110 | −0.194 |
| COX1 | 26.43 | 34.18 | 17.71 | 21.68 | 60.61 | 39.39 | −0.128 | −0.101 |
| COX2 | 28.97 | 30.57 | 17.76 | 22.71 | 59.53 | 40.47 | −0.027 | −0.122 |
| ATP8 | 32.70 | 36.48 | 10.69 | 20.13 | 69.18 | 30.82 | −0.055 | −0.306 |
| ATP6 | 26.44 | 36.06 | 13.22 | 24.28 | 62.50 | 37.50 | −0.154 | −0.295 |
| ND1 | 26.09 | 34.50 | 15.23 | 24.17 | 60.60 | 39.40 | −0.139 | −0.227 |
| ND6 | 27.31 | 34.94 | 13.05 | 24.70 | 62.25 | 37.75 | −0.123 | −0.309 |
| CYTB | 25.70 | 33.07 | 15.09 | 26.14 | 58.77 | 41.23 | −0.125 | −0.268 |
| ND4L | 25.70 | 33.07 | 15.09 | 26.14 | 68.01 | 31.99 | −0.125 | −0.268 |
| ND4 | 28.56 | 37.21 | 13.57 | 20.66 | 65.77 | 34.23 | −0.132 | −0.207 |
| ND5 | 30.15 | 33.13 | 13.35 | 23.36 | 63.29 | 36.71 | −0.047 | −0.273 |
| COX3 | 25.90 | 32.18 | 19.36 | 22.56 | 58.08 | 41.92 | −0.108 | −0.076 |
| ND3 | 27.35 | 39.32 | 15.10 | 18.23 | 66.67 | 33.33 | −0.180 | −0.094 |
| ND2 | 28.89 | 37.38 | 14.39 | 19.33 | 66.27 | 33.73 | −0.128 | −0.147 |
| Position | Size (bp) | Intergenic Nucleotides | Codon | Strand | ||||
|---|---|---|---|---|---|---|---|---|
| Gene | From | To | Start | Stop | ||||
| 1 | COX1 | 1 | 1536 | 1536 | ATG | TAA | H | |
| 2 | COX2 | 1575 | 2261 | 687 | 38 | ATG | TAA | H |
| 3 | trnD(guc) | 2268 | 2336 | 69 | 6 | H | ||
| 4 | ATP8 | 2338 | 2496 | 159 | 1 | ATG | TAA | H |
| 5 | ATP6 | 2512 | 3207 | 696 | 15 | ATG | TAA | H |
| 6 | trnM(cau) | 3240 | 3306 | 67 | 32 | L | ||
| 7 | trnY(gua) | 3310 | 3377 | 68 | 3 | L | ||
| 8 | trnC(gca) | 3382 | 3446 | 65 | 4 | L | ||
| 9 | trnW(uca) | 3448 | 3514 | 67 | 1 | L | ||
| 10 | trnQ(uug) | 3514 | 3578 | 65 | −1 | L | ||
| 11 | trnG(ucc) | 3590 | 3656 | 67 | 11 | L | ||
| 12 | trnE(uuc) | 3710 | 3777 | 68 | 53 | L | ||
| 13 | s-rRNA | 3856 | 4756 | 901 | 78 | H | ||
| 14 | trnV(uac) | 4754 | 4822 | 69 | −3 | H | ||
| 15 | l-rRNA | 4801 | 6219 | 1419 | −22 | H | ||
| 16 | trnL(uaa) | 6210 | 6277 | 68 | −10 | H | ||
| 17 | trnL(uag) | 6284 | 6352 | 69 | 6 | H | ||
| 18 | ND1 | 6353 | 7291 | 939 | 0 | ATG | TAA | H |
| 19 | trnP(ugg) | 7301 | 7369 | 69 | 9 | H | ||
| 20 | ND6 | 7374 | 7871 | 498 | 4 | ATG | TAA | H |
| 21 | CYTB | 7890 | 9029 | 1140 | 18 | ATG | TAA | H |
| 22 | trnS(uga) | 9040 | 9107 | 68 | 10 | H | ||
| 23 | trnT(ugu) | 9111 | 9178 | 68 | 3 | L | ||
| 24 | ND4L | 9185 | 9481 | 297 | 6 | ATG | TAG | H |
| 25 | ND4 | 9505 | 10845 | 1341 | 23 | ATT | TAA | H |
| 26 | trnH(gug) | 10852 | 10918 | 67 | 6 | H | ||
| 27 | ND5 | 10947 | 12624 | 1678 | 28 | ATT | CTT | H |
| 28 | trnF(gaa) | 12663 | 12732 | 70 | 38 | H | ||
| CR | 12733 | 13505 | 773 | 0 | ||||
| 29 | COX3 | 13506 | 14285 | 780 | 773 | ATG | TAA | H |
| 30 | trnK(uuu) | 14307 | 14378 | 72 | 21 | H | ||
| 31 | trnA(ugc) | 14385 | 14451 | 67 | 6 | H | ||
| 32 | trnR(ucg) | 14459 | 14527 | 69 | 7 | H | ||
| 33 | trnN(guu) | 14533 | 14602 | 70 | 5 | H | ||
| 34 | trnI(gau) | 14604 | 14671 | 68 | 1 | H | ||
| 35 | ND3 | 14679 | 15029 | 351 | 7 | ATA | TAA | H |
| 36 | trnS(gcu) | 15029 | 15095 | 67 | −1 | H | ||
| 37 | ND2 | 15123 | 16053 | 931 | 27 | ATG | AAT | H |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Yang, M.; Duan, H.; Liao, X. The Mitochondrial Genome of Littoraria melanostoma Reveals a Phylogenetic Relationship within Littorinimorpha. Diversity 2023, 15, 1005. https://doi.org/10.3390/d15091005
Chen K, Yang M, Duan H, Liao X. The Mitochondrial Genome of Littoraria melanostoma Reveals a Phylogenetic Relationship within Littorinimorpha. Diversity. 2023; 15(9):1005. https://doi.org/10.3390/d15091005
Chicago/Turabian StyleChen, Kun, Mingliu Yang, Haisheng Duan, and Xin Liao. 2023. "The Mitochondrial Genome of Littoraria melanostoma Reveals a Phylogenetic Relationship within Littorinimorpha" Diversity 15, no. 9: 1005. https://doi.org/10.3390/d15091005
APA StyleChen, K., Yang, M., Duan, H., & Liao, X. (2023). The Mitochondrial Genome of Littoraria melanostoma Reveals a Phylogenetic Relationship within Littorinimorpha. Diversity, 15(9), 1005. https://doi.org/10.3390/d15091005