

Changes in Composition of Mollusks within Corallina officinalis Turfs in South Istria, Adriatic Sea, as a Response to Anthropogenic Impact
 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
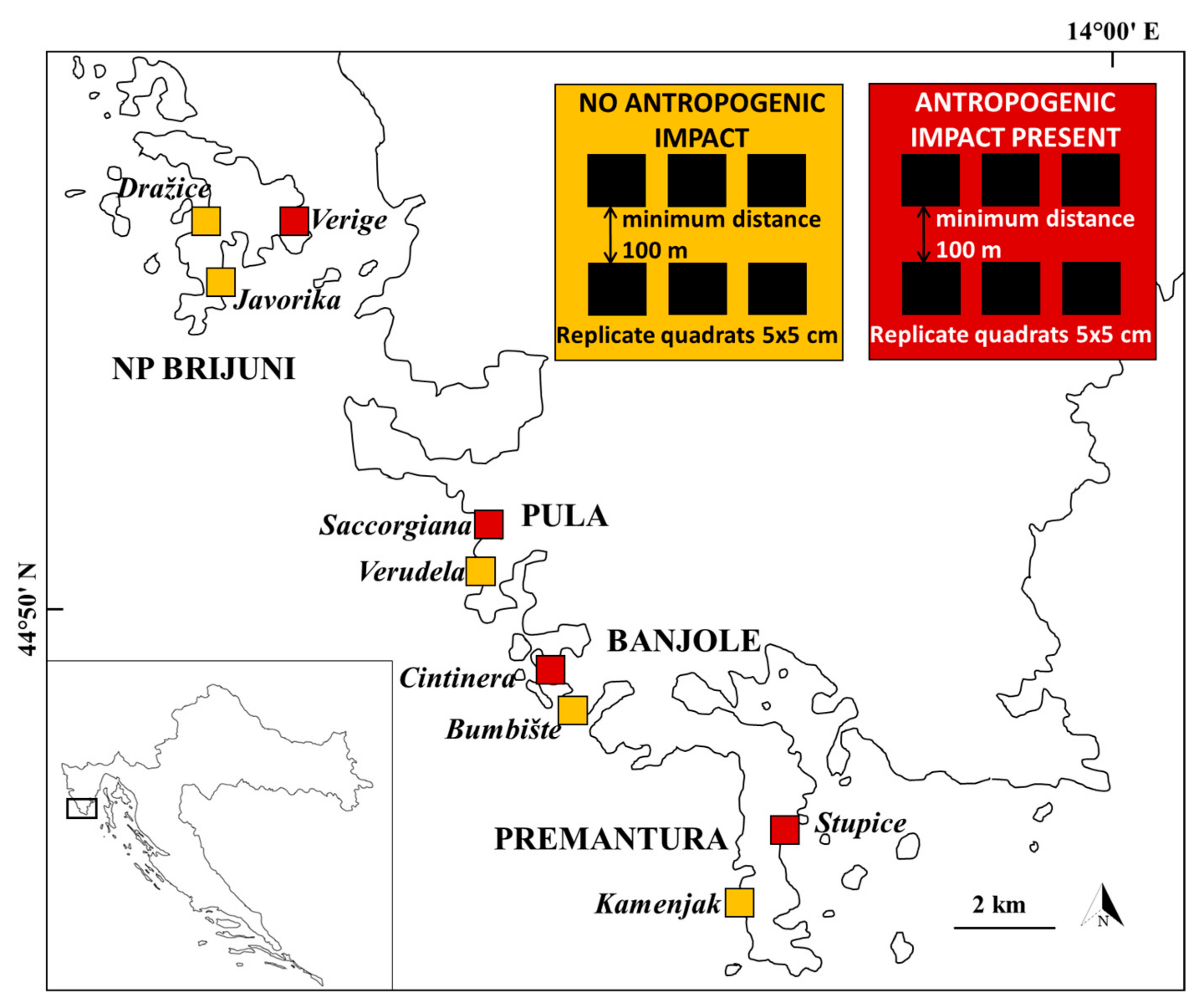
2.1. Sampling Design
2.2. Sampling Methods
2.3. Quantifying Anthropogenic Impact
2.4. Statistical Data Processing
3. Results
3.1. Categorization of Research Locations Depending on Anthropogenic Impact
3.2. Diversity of Mollusks within Corallina Officinalis Settlements
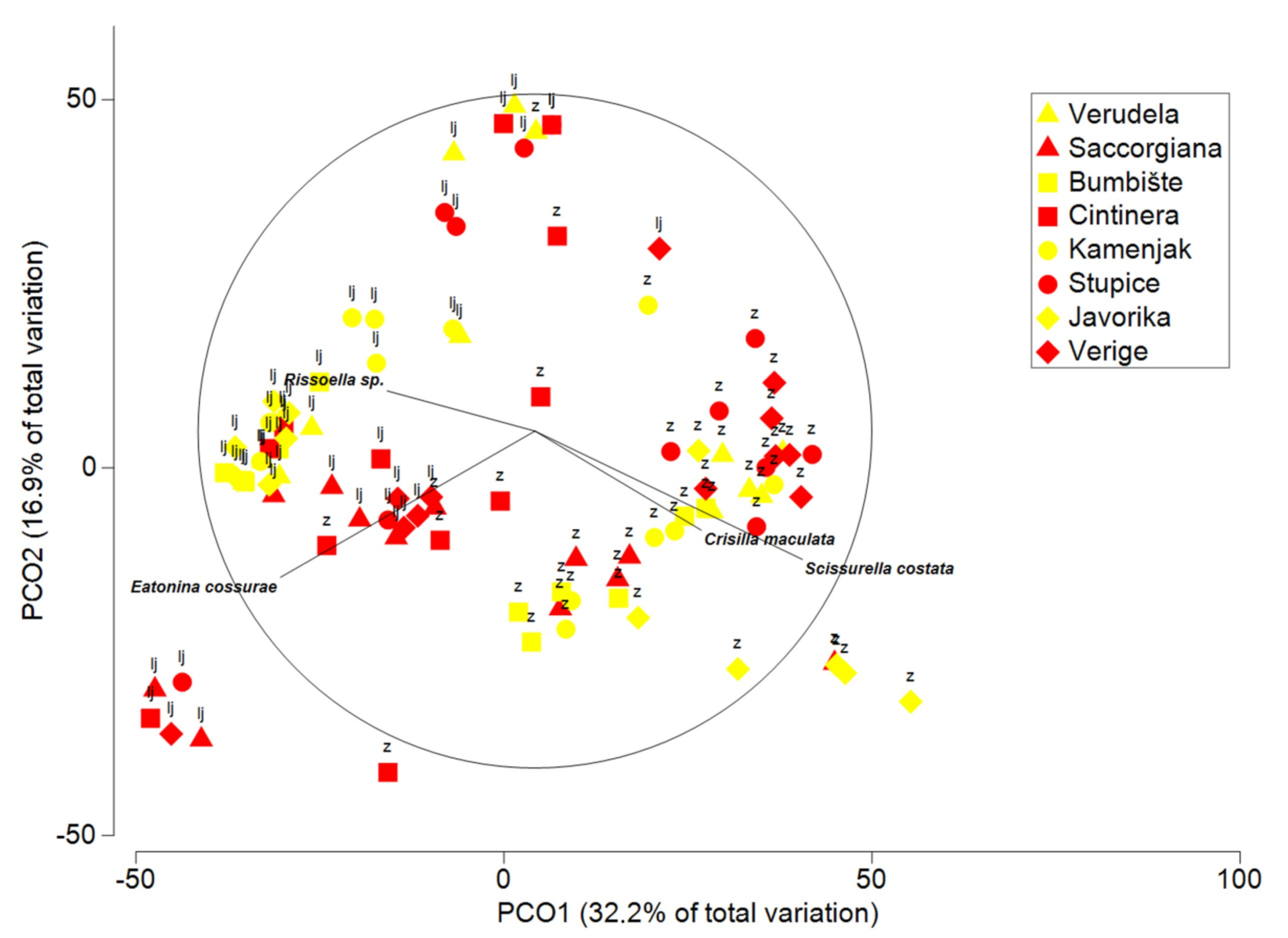
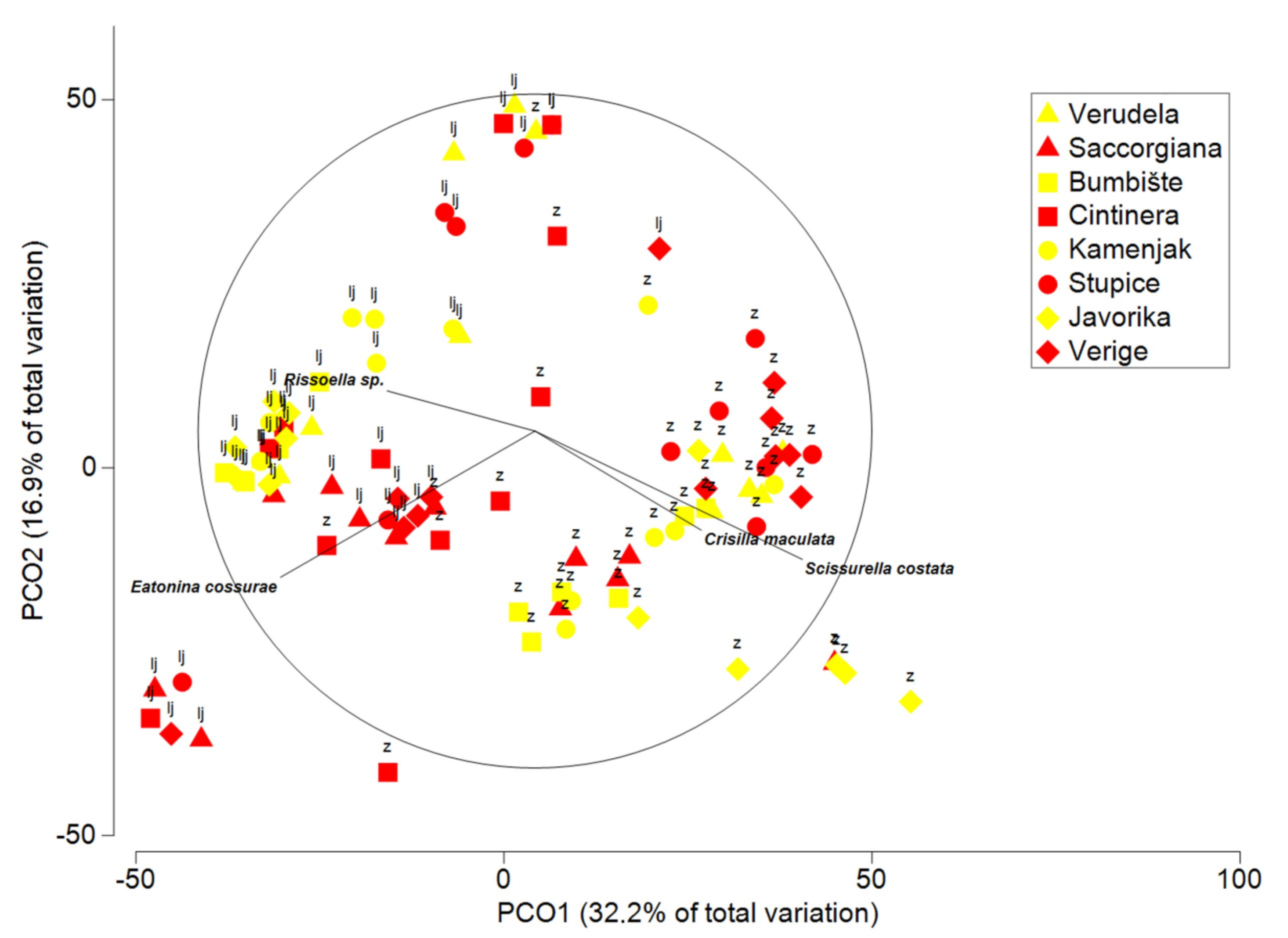
3.3. Multivariate Analysis of Gastropods
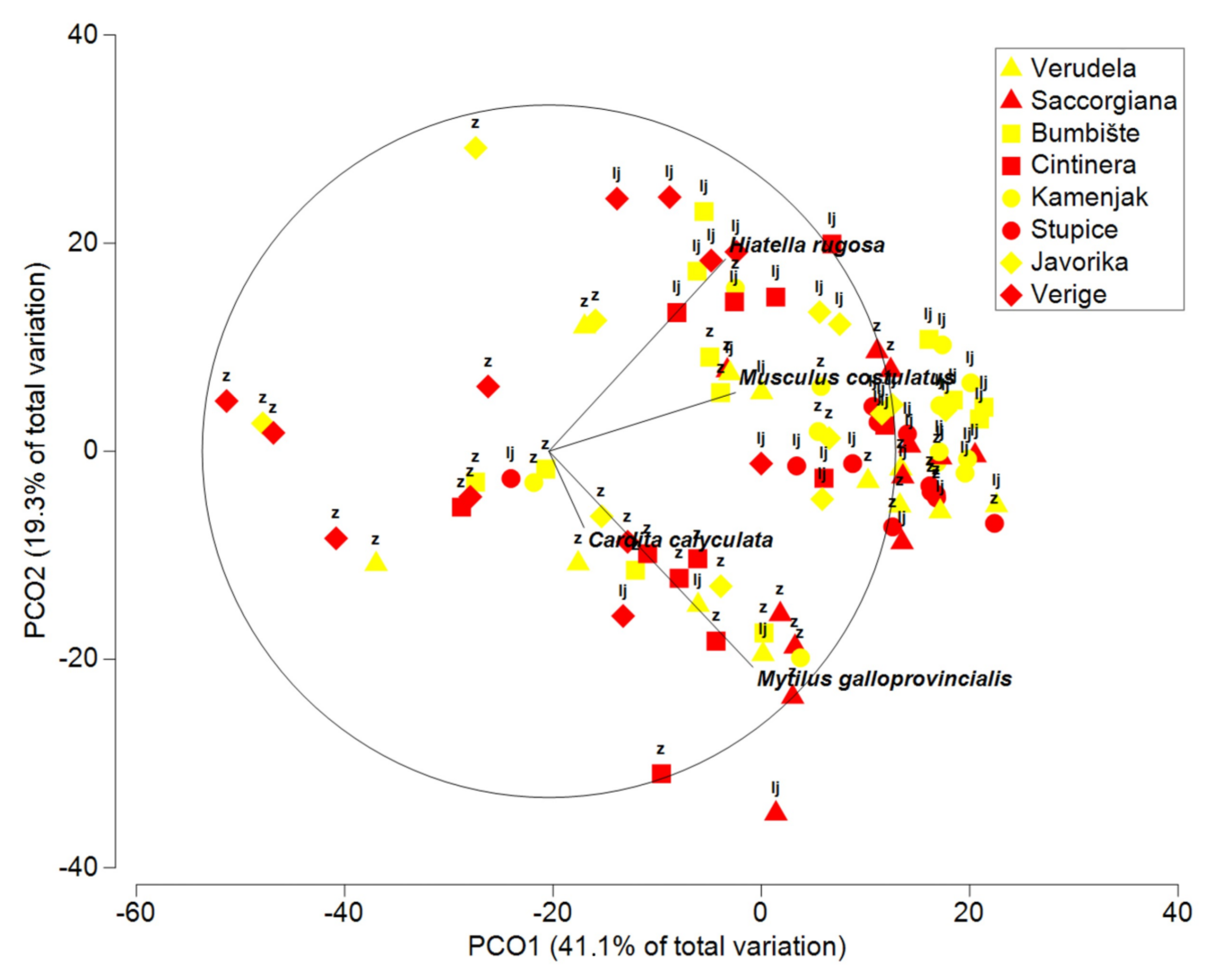
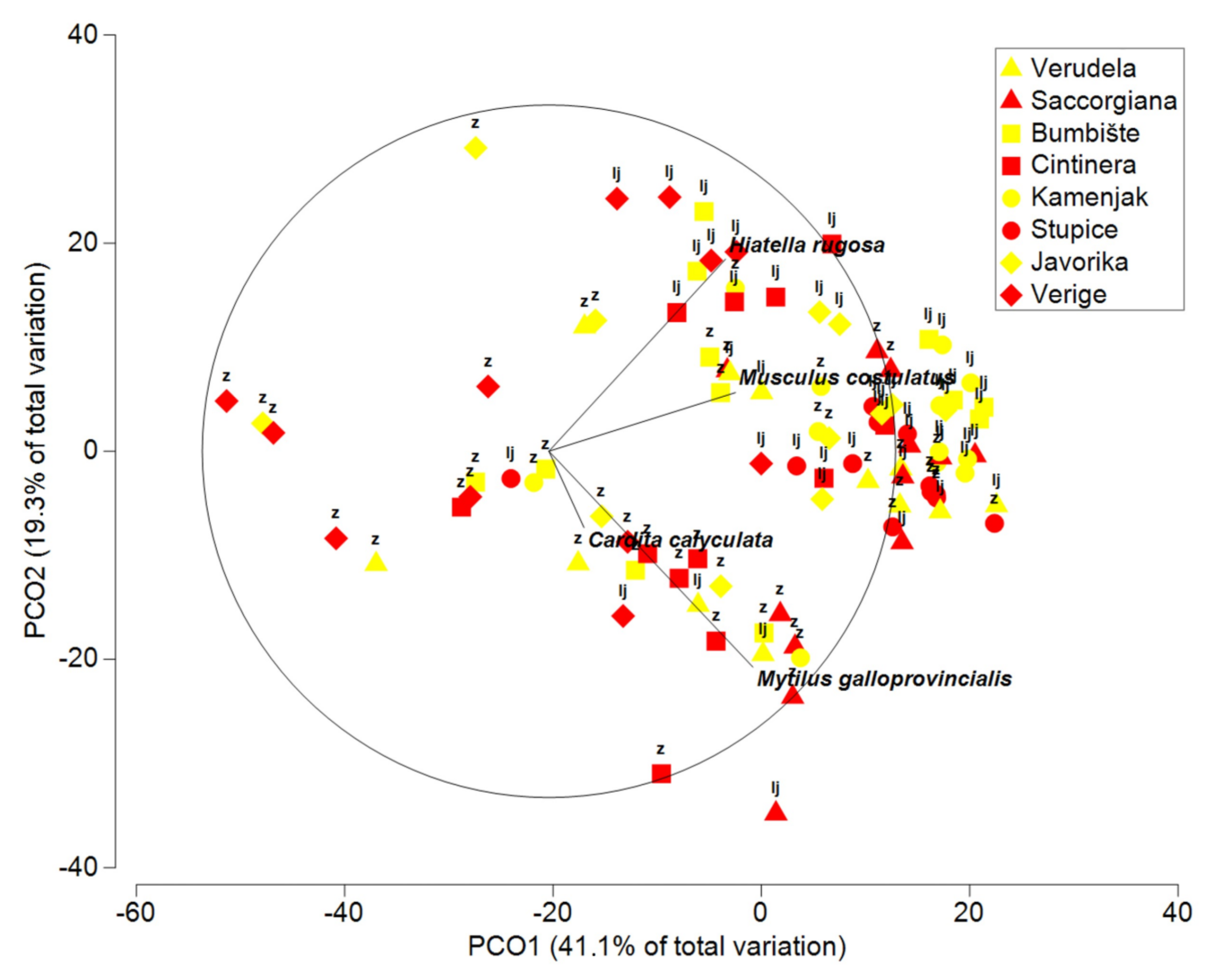
3.4. Multivariate Analysis of Bivalves
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative Human Impacts on Mediterranean and Black Sea Marine Ecosystems: Assessing Current Pressures and Opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [PubMed]
- Fraschetti, S.; Bianchi, C.; Terlizzi, A.; Fanelli, G.; Morri, C.; Boero, F. Spatial variability and human disturbance in shallow subtidal hard substrate assemblages: A regional approach. Mar. Ecol. Prog. Ser. 2001, 212, 1–12. [Google Scholar] [CrossRef]
- Bertolino, M.; Betti, F.; Bo, M.; Cattaneo-Vietti, R.; Pansini, M.; Romero, J.; Bavestrello, G. Changes and stability of a Mediterranean hard bottom benthic community over 25 years. J. Mar. Biol. Assoc. UK 2015, 96, 341–350. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Esposito, V.; Giacobbe, S.; Renzi, M.; Mangano, M.C.; Vivona, P.; Consoli, P.; Scotti, G.; Andaloro, F.; Romeo, T. Ecological assessment of a heavily human-stressed area in the Gulf of Milazzo, Central Mediterranean Sea: An integrated study of biological, physical and chemical indicators. Mar. Pollut. Bull. 2016, 106, 260–273. [Google Scholar] [CrossRef]
- Mosbahi, N.; Serbaji, M.M.; Pezy, J.-P.; Neifar, L.; Dauvin, J.-C. Response of benthic macrofauna to multiple anthropogenic pressures in the shallow coastal zone south of Sfax (Tunisia, central Mediterranean Sea). Environ. Pollut. 2019, 253, 474–487. [Google Scholar] [CrossRef]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr. Mar. Biol. Annu. Rev. 2006, 44, 123–195. [Google Scholar]
- Pasquali, D.; Marucci, A. The Effects of Urban and Economic Development on Coastal Zone Management. Sustainability 2021, 13, 6071. [Google Scholar] [CrossRef]
- Buono, F.; Soriani, S.; Camuffo, M.; Tonino, M.; Bordin, A. The difficult road to Integrated Coastal Zone Management implementation in Italy: Evidences from the Italian North Adriatic Regions. Ocean Coast. Manag. 2015, 114, 21–31. [Google Scholar] [CrossRef]
- Murray, S.N.; Littler, M.M. Patterns of Algal Succession in a Perturbated Marine Intertidal Community. J. Phycol. 1978, 14, 506–512. [Google Scholar] [CrossRef]
- Chakraborty, S.; Bhattacharya, T.; Singh, G.; Maity, J.P. Benthic macroalgae as biological indicators of heavy metal pollution in the marine environments: A biomonitoring approach for pollution assessment. Ecotoxicol. Environ. Saf. 2014, 100, 61–68. [Google Scholar] [CrossRef]
- Duffy, J.E.; Benedetti-Cecchi, L.; Trinanes, J.; Muller-Karger, F.E.; Ambo-Rappe, R.; Boström, C.; Buschmann, A.H.; Byrnes, J.; Coles, R.G.; Creed, J.; et al. Toward a Coordinated Global Observing System for Seagrasses and Marine Macroalgae. Front. Mar. Sci. 2019, 6, 317. [Google Scholar] [CrossRef]
- Rindi, F.; Gavio, B.; Díaz-Tapia, P.; Di Camillo, C.G.; Romagnoli, T. Long-term changes in the benthic macroalgal flora of a coastal area affected by urban impacts (Conero Riviera, Mediterranean Sea). Biodivers. Conserv. 2020, 29, 2275–2295. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Lipej, L. Loss of canopy-forming algal richness and coverage in the northern Adriatic Sea. Ecol. Indic. 2021, 125, 107501. [Google Scholar] [CrossRef]
- Matias, M.G.; Underwood, A.J.; Coleman, R.A. Interactions of components of habitats alter composition and variability of assemblages. J. Anim. Ecol. 2007, 76, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Ros, J.D.; Cardell, M.J. Effect on benthic communities of a major input of organic matter and other pollutants (coast off Barcelona, Western Mediterranean). Toxicol. Environ. Chem. 1991, 31, 441–450. [Google Scholar] [CrossRef]
- Dauer, D.M. Biological criteria, environmental health and estuarine macrobenthic community structure. Mar. Pollut. Bull. 1993, 26, 249–257. [Google Scholar] [CrossRef]
- Cabana, D.; Sigala, K.; Nicolaidou, A.; Reizopoulou, S. Towards the implementation of the Water Framework Directive in Mediterranean transitional waters: The use of macroinvertebrates as biological quality elements. Adv. Oceanogr. Limnol. 2013, 4, 212–240. [Google Scholar] [CrossRef]
- Dauvin, J.; Ruellet, T. Polychaete/amphipod ratio revisited. Mar. Pollut. Bull. 2007, 55, 215–224. [Google Scholar] [CrossRef]
- Borja, A.; Franco, J.; Pérez, V. A Marine Biotic Index to Establish the Ecological Quality of Soft-Bottom Benthos Within European Estuarine and Coastal Environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- George, J.D. The polychaetes of Lewis and Harris with notes on other marine invertebrates. Proc. R. Soc. Edinburgh. Sect. B. Biol. Sci. 1979, 77, 189–216. [Google Scholar] [CrossRef]
- Johnson, S.B.; Attramadal, Y.G. Reproductive behaviour and larval development of Tanais cavolinii (Crustacea: Tanaidacea). Mar. Biol. 1982, 71, 11–16. [Google Scholar] [CrossRef]
- López, C.A.; Stotz, W.B. Description of the fauna associated with Corallina officinalis L. in the intertidal of the rocky shore of Palo Colorado (Los Vilos, IV-region, Chile). Oceanogr. Lit. Rev. 1998, 3, 512. [Google Scholar]
- Bussell, J.A.; Lucas, I.A.; Seed, R. Patterns in the invertebrate assemblage associated with Corallina officinalis in tide pools. J. Mar. Biol. Assoc. UK 2007, 87, 383–388. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Castilla, J.C.; Prado, L.; York, P.; Schwindt, E.; Bortolus, A. Spatial variation in molluscan assemblages from coralline turfs of Argentinean Patagonia. J. Molluscan Stud. 2007, 73, 139–146. [Google Scholar] [CrossRef]
- Buršić, M.; Iveša, L.; Jaklin, A.; Pijevac, M.A. A preliminary study on the diversity of invertebrates associated with Corallina officinalis Linnaeus in southern Istrian peninsula. Acta Adriat. 2019, 60, 127–136. [Google Scholar] [CrossRef]
- Buršić, M.; Iveša, L.; Jaklin, A.; Pijevac, M.A.; Kučinić, M.; Štifanić, M.; Neal, L.; Mađarić, B.B. DNA Barcoding of Marine Mollusks Associated with Corallina officinalis Turfs in Southern Istria (Adriatic Sea). Diversity 2021, 13, 196. [Google Scholar] [CrossRef]
- Nordsieck, F. Die Europäischen Meeres-Gehäuseschnecken (Prosobranchia); Gustav Fischer Verlag: Stuttgart, Germany, 1968; 273p. [Google Scholar]
- Nordsieck, F. Die Europäischen Meeresmuscheln (Bivalvia); Gustav Fisher Verlag: Stuttgart, Germany, 1969; 256p. [Google Scholar]
- Parenzan, P. Carta D’identità delle Conchiglie del Mediterraneo. Vol. I. Gasteropodi; Bios Taras: Taranto, Italy, 1970; 283p. [Google Scholar]
- Parenzan, P. Carta D’identità delle Conchiglie del Mediterraneo. Vol. II; Bivalvi, Prima Parte; Bios Taras: Taranto, Italy, 1974; 277p. [Google Scholar]
- Sabelli, B.; Gianuzzi-Savelli, R.; Bedulli, D. Catalogo Annotato dei Molluschi Marini del Mediterraneo. Vol. I; Libreria Naturalistica Bolognese: Bologna, Italy, 1990; 348p. [Google Scholar]
- Poppe, G.T.; Goto, Y. European Seashells. Vol. II (Scaphopoda, Bivalvia, Cephalopoda); Verlag Christa Hemmen: Wiesbaden, Germany, 1993; 221p. [Google Scholar]
- Gianuzzi-Savelli, R.; Pusateri, F.; Palmeri, A.; Ebreo, C. Atlante delle Conchiglie Marine del Mediterraneo; La Conchiglia: Roma, Italy, 1996; 258p. [Google Scholar]
- Gofas, S.; Moreno, D.; Salas, C. (Eds.) Moluscos Marinos de Andalucía. Vol. I; Universidad de Málaga: Málaga, Spain, 2011; 342p. [Google Scholar]
- Gofas, S.; Moreno, D.; Salas, C. (Eds.) Moluscos marinos de Andalucía. Vol. II; Universidad de Málaga: Málaga, Spain, 2011; 455p. [Google Scholar]
- Flo, E.; Garcés, E.; Camp, J. Land Uses Simplified Index (LUSI): Determining Land Pressures and Their Link with Coastal Eutrophication. Front. Mar. Sci. 2019, 6, 18. [Google Scholar] [CrossRef]
- Borja, Á.; Mader, J.; Muxika, I. Instructions for the use of the AMBI index software (version 5.0). Rev. Invest. Mar. 2012, 19, 71–82. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth Marine Laboratory: Plymouth, UK, 2006; p. 190. [Google Scholar]
- Mehdipour, N.; Gerami, M.H.; Nemati, H. Assessing benthic health of hard substratum macrobenthic community using soft bottom indicators and their relationship with environmental condition. Iran. Fish. J. Sci. 2018, 17, 641–656. [Google Scholar] [CrossRef]
- Urra, J.; Rueda, J.; Ramírez, M.; Marina, P.; Tirado, C.; Salas, C.; Gofas, S. Seasonal variation of molluscan assemblages in different strata of photophilous algae in the Alboran Sea (western Mediterranean). J. Sea Res. 2013, 83, 83–93. [Google Scholar] [CrossRef]
- Terlizzi, A.; Scuderi, D.; Fraschetti, S.; Anderson, M.J. Quantifying effects of pollution on biodiversity: A case study of highly diverse molluscan assemblages in the Mediterranean. Mar. Biol. 2005, 148, 293–305. [Google Scholar] [CrossRef]
- Sánchez-Moyano, J.; Estacio, F.; García-Adiego, E.; García-Gómez, J. The Molluscan Epifauna of the alga Halopteris scoparia in southern Spain as a bioindicator of coastal environmental conditions. J. Molluscan Stud. 2000, 66, 431–448. [Google Scholar] [CrossRef]
- Terlizzi, A.; Scuderi, D.; Fraschetti, S.; Guidetti, P.; Boero, F. Molluscs on subtidal cliffs: Patterns of spatial distribution. J. Mar. Biol. Assoc. UK 2003, 83, 165–172. [Google Scholar] [CrossRef]
- Avila, S.P. The littoral molluscs (Gastropoda, Bivalvia and Polyplacophora) of Sao Vicente, Capelas (Sao Miguel island, Azores): Ecology and biological associations to algae. Iberus 2003, 21, 11–33. [Google Scholar]
- Tamburello, L.; Papa, L.; Guarnieri, G.; Basconi, L.; Zampardi, S.; Scipione, M.B.; Terlizzi, A.; Zupo, V.; Fraschetti, S. Are we ready for scaling up restoration actions? An insight from Mediterranean macroalgal canopies. PLoS ONE 2019, 14, e0224477. [Google Scholar] [CrossRef]
- Chiarore, A.; Fioretti, S.; Meccariello, A.; Saccone, G.; Patti, F.P. Molluscs community associated with the brown algae of the genus Cystoseira in the Gulf of Naples (South Tyrrhenian Sea). bioRxiv 2017, 160200. [Google Scholar] [CrossRef]
- Dommasnes, A. On the fauna of Corallina officinalis L. in western Norway. Sarsia 1969, 38, 71–86. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; García-Gómez, J.C. Soft bottom mollusc assemblages and pollution in a harbour with two opposing entrances. Estuar. Coast. Shelf Sci. 2004, 60, 273–283. [Google Scholar] [CrossRef]
- Marchini, A.; Gauzer, K.; Occhipinti-Ambrogi, A. Spatial and temporal variability of hard-bottom macrofauna in a disturbed coastal lagoon (Sacca di Goro, Po River Delta, Northwestern Adriatic Sea). Mar. Pollut. Bull. 2004, 48, 1084–1095. [Google Scholar] [CrossRef]
- Liuzzi, M.; Gappa, J.L. Macrofaunal assemblages associated with coralline turf: Species turnover and changes in structure at different spatial scales. Mar. Ecol. Prog. Ser. 2008, 363, 147–156. [Google Scholar] [CrossRef]
- Lehane, C.; Davenport, J. Ingestion of bivalve larvae by Mytilus edulis: Experimental and field demonstrations of larviphagy in farmed blue mussels. Mar. Biol. 2004, 145, 101–107. [Google Scholar] [CrossRef]
- Lehane, C.; Davenport, J. A 15-month study of zooplankton ingestion by farmed mussels (Mytilus edulis) in Bantry Bay, Southwest Ireland. Estuar. Coast. Shelf Sci. 2006, 67, 645–652. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Impact | Freshwater Impact | Pressure Score | ||
|---|---|---|---|---|
| Urban Area | Agricultural Area | Industrial Area | Salinity | |
| ≤10% | ≤10% | ≥37.5 | 0 | |
| ≤33% | 10–40% | >10% | 34.5–37.5 | 1 |
| 33–66% | >40% | <34.5 | 2 | |
| >66% | 3 | |||
| Correction factor for coastline morphology | concave | ×1.25 | ||
| convex | ×0.75 | |||
| straight | ×1.00 | |||
| Area | Location | Individual Values | Score | ||||
|---|---|---|---|---|---|---|---|
| PULA | Verudela | 1 | 0 | 1 | 0 | 0.75 | 1.50 |
| Saccorgiana * | 2 | 1 | 1 | 0 | 1.25 | 5.00 | |
| BANJOLE | Bumbište | 1 | 0 | 0 | 0 | 0.75 | 0.75 |
| Cintinera * | 2 | 1 | 1 | 0 | 1.25 | 5.00 | |
| PREMANTURA | Kamenjak | 1 | 0 | 0 | 0 | 1.25 | 1.25 |
| Stupice * | 2 | 1 | 0 | 0 | 1.25 | 3.75 | |
| NP BRIJUNI | Javorika | 1 | 0 | 0 | 0 | 1.00 | 1.00 |
| Dražice | 1 | 0 | 0 | 0 | 1.25 | 1.25 | |
| Verige * | 1 | 0 | 1 | 0 | 1.00 | 2.00 | |
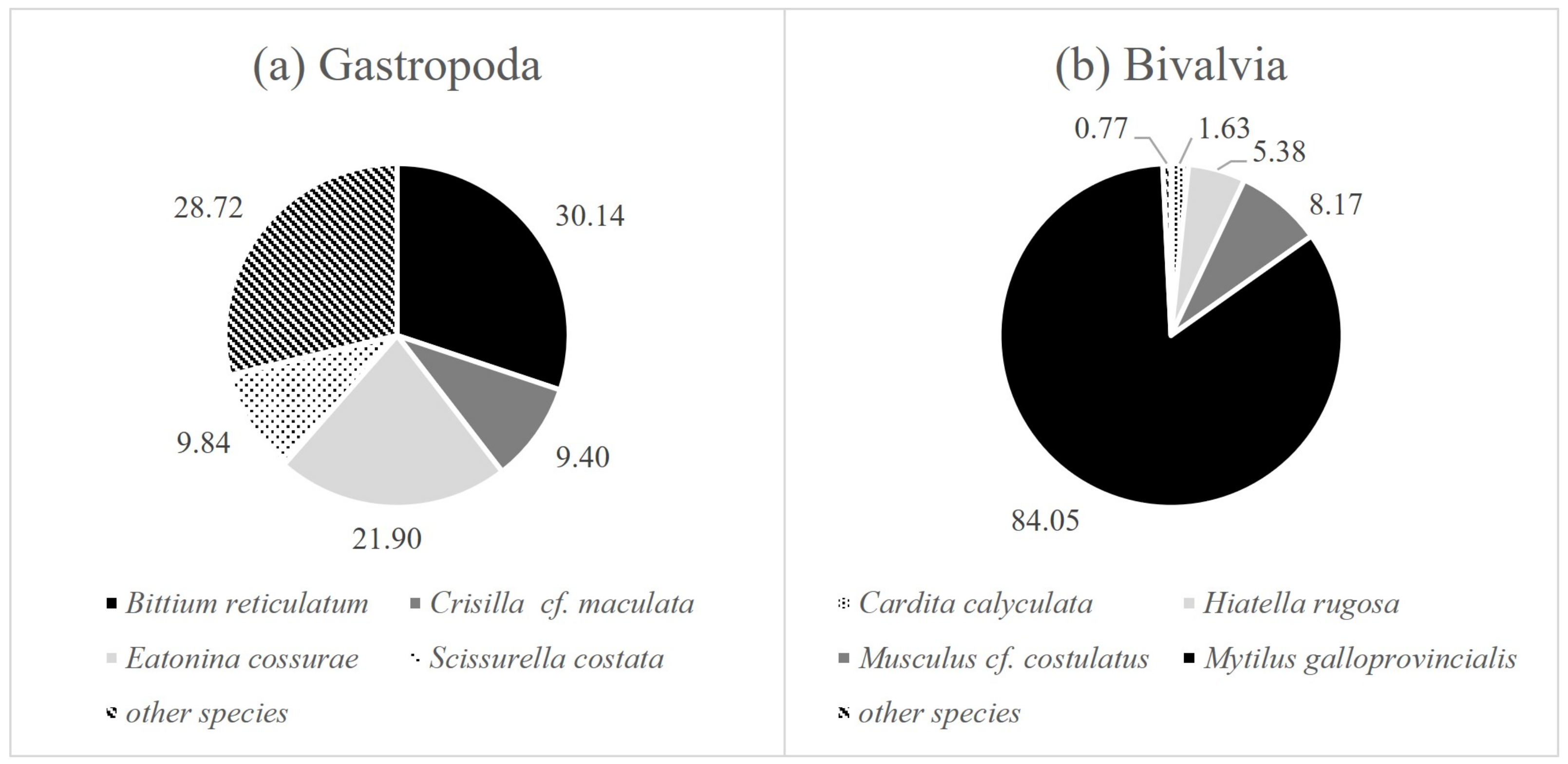
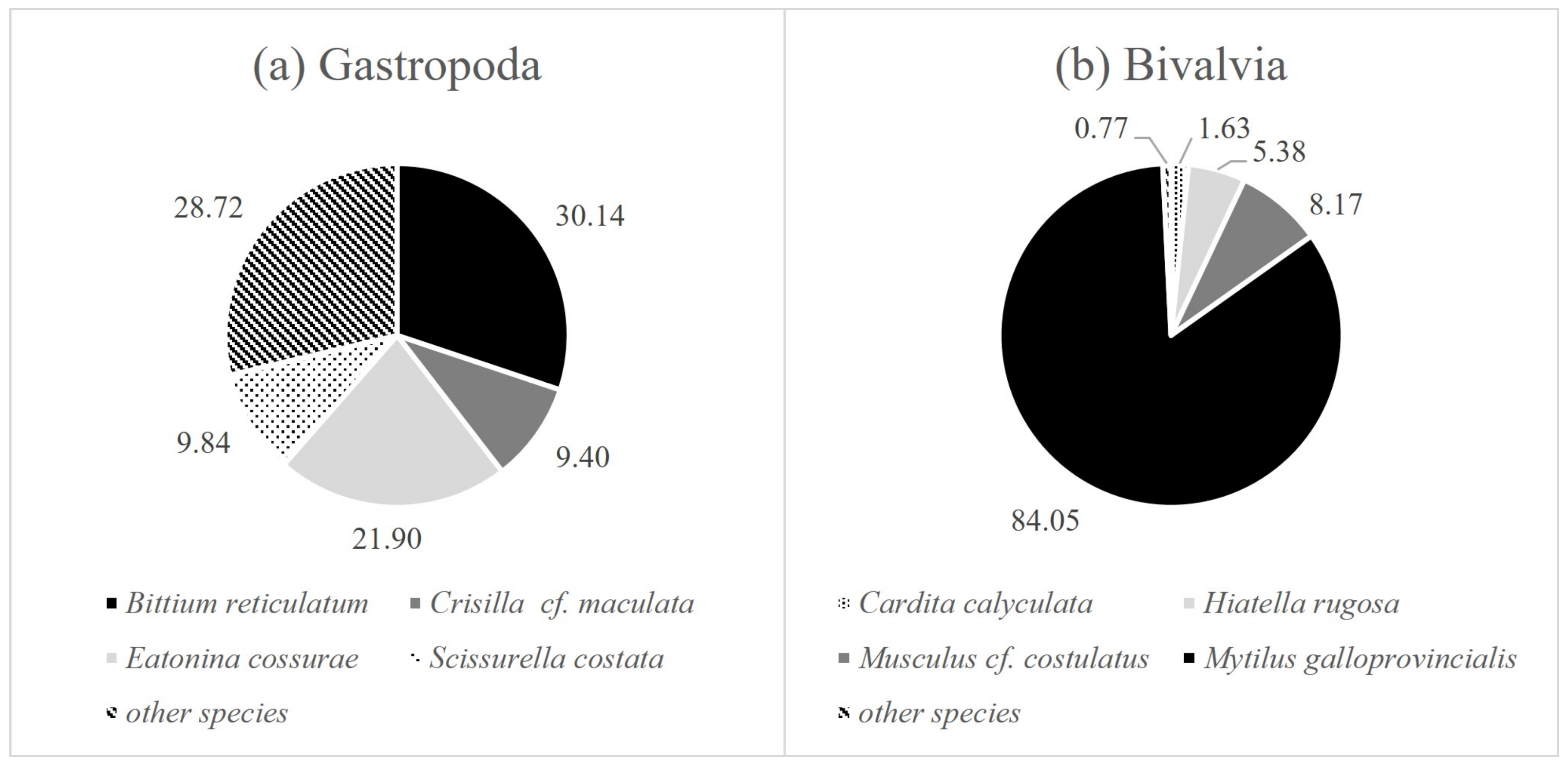
| Family | Species | EG | Share in the Total Number of Mollusks (%) | |
|---|---|---|---|---|
| Outside A.P. | Under A.P. | |||
| GASTROPODA | ||||
| Aplysiidae | Aplysia sp.juv. (Linnaeus, 1767) | I | 0.01 | |
| Cerithiidae | Bittium reticulatum (da Costa, 1778) | I | 7.21 | 2.55 |
| Cerithiidae | Cerithium vulgatum (Bruguière, 1792) | II | 0.01 | |
| Cerithiopsidae | Cerithiopsis tubercularis (Montagu, 1803) | I | 0.01 | |
| Cingulopsidae | Eatonina cossurae (Calcara, 1841) | 3.72 | 3.82 | |
| Cingulopsidae | Eatonina sp. (Thiele, 1912) | 0.04 | 0.01 | |
| Columbellidae | Columbella rustica (Linnaeus, 1758) | I | 0.02 | |
| Dorididae | Dorididae indet.juv. (Rafinesque, 1815) | I | 0.01 | |
| Eulimidae | Vitreolina antiflexa (Monterosato, 1884) | I | 0.02 | 0.02 |
| Fissurellidae | Fissurellidae indet.juv (J. Fleming, 1822) | 0.01 | 0.02 | |
| Mitridae | Episcomitra cornicula (Linnaeus, 1758) | 0.01 | ||
| Muricidae | Hexaplex trunculus (Linnaeus, 1758) | I | 0.01 | |
| Muricidae | Muricopsis cristata (Brocchi, 1814) | 0.01 | ||
| Muricidae | Ocenebra cf. edwardsii (Payraudeau, 1826) | 0.04 | 0.05 | |
| Omalogyridae | Ammonicera fischeriana (Monterosato, 1869) | 0.00 | 0.05 | |
| Patellidae | Patella caerulea (Linnaeus, 1758) | 0.12 | 0.05 | |
| Phasianellidae | Tricolia pullus (Linnaeus, 1758) | I | 0.01 | |
| Pyramidellidae | Brachystomia eulimoides (Hanley, 1844) | 0.03 | 0.06 | |
| Pyramidellidae | Megastomia winfriedi (Peñas and Rolán, 1999) | 0.22 | 0.16 | |
| Pyramidellidae | Odostomia plicata (Montagu, 1803) | II | 0.01 | 0.01 |
| Pyramidellidae | Parthenina emaciata (Brusina, 1866) | I | 0.03 | 0.01 |
| Pyramidellidae | Spiralina alpinoligustica (Sacco, 1892) | I | 0.02 | 0.03 |
| Rissoellidae | Rissoella sp. (Gray, 1847) | I | 1.38 | 0.16 |
| Rissoidae | Alvania poucheti (Dautzenberg, 1889) | I | 0.17 | 0.03 |
| Rissoidae | Alvania rudis(Philippi, 1844) | I | 0.01 | |
| Rissoidae | Alvania sp. 1 (Risso, 1826) | I | 0.03 | |
| Rissoidae | Alvania sp. 2 (Risso, 1826) | I | 0.01 | |
| Rissoidae | Alvania cf. carinata juv. (da Costa, 1778) | 0.02 | ||
| Rissoidae | Cingula trifasciata (J. Adams, 1800) | I | 0.03 | 0.01 |
| Rissoidae | Crisilla beniamina (Monterosato, 1884) | 0.09 | 0.01 | |
| Rissoidae | Crisilla innominata (R. B. Watson, 1897) | 3.60 | 0.68 | |
| Rissoidae | Crisilla iunoniae (Palazzi, 1988) | 0.13 | 0.03 | |
| Rissoidae | Crisilla cf. maculata (Monterosato, 1869) | 2.13 | 0.95 | |
| Rissoidae | Pusillina philippi (Aradas and Maggiore, 1844) | 0.31 | 0.06 | |
| Rissoidae | Rissoa splendida (Eichwald, 1830) | 0.07 | 0.02 | |
| Rissoidae | Setia sp. (H. Adams and A. Adams, 1852) | 0.01 | ||
| Scissurellidae | Scissurella costata (d’Orbigny, 1824) | 1.62 | 1.79 | |
| Scissurellidae | Sinezona cingulata (O. G. Costa, 1861) | 0.19 | 0.06 | |
| Siphonariidae | Siphonaria cf. pectinata juv. (Linnaeus, 1758) | I | 0.05 | 0.11 |
| Triphoridae | Monophorus perversus (Linnaeus, 1758) | I | 0.01 | 0.00 |
| Tritoniidae | Duvaucelia manicata (Deshayes, 1853) | I | 0.01 | 0.03 |
| Trochidae | Clanculus sp.juv. (Montfort, 1810) | 0.01 | ||
| Trochidae | Gibbula sp.juv. (Risso, 1826) | I | 0.01 | |
| Trochidae | Gibbula cf. turbinoides (Deshayes, 1835) | I | 0.04 | 0.03 |
| Trochidae | Phorcus turbinatus (Born, 1778) | 0.27 | 0.16 | |
| Trochidae | Steromphala adriatica (Philippi, 1844) | I | 0.00 | 0.01 |
| Gastrpoda indet.juv. | 0.25 | 0.02 | ||
| BIVALVIA | ||||
| Arcidae | Arca noae (Linnaeus, 1758) | I | 0.01 | 0.02 |
| Arcidae | Arca sp. (Linnaeus, 1758) | I | 0.05 | 0.05 |
| Carditidae | Cardita calyculata (Linnaeus, 1758) | I | 1.11 | 1.63 |
| Chamidae | Chama gryphoides (Linnaeus, 1758) | 0.01 | ||
| Hiatellidae | Hiatella rugosa (Linnaeus, 1767) | I | 5.46 | 3.04 |
| Lasaeidae | Lasaea cf. rubra (Gmelin, 1791) | II | 0.09 | 0.61 |
| Limidae | Lima lima (Linnaeus, 1758) | I | 0.02 | 0.01 |
| Lucinidae | Lucinella sp. (Monterosato, 1884) | I | 0.01 | |
| Mytilidae | Gregariella semigranata (Reeve, 1858) | I | 0.02 | |
| Mytilidae | Lithophaga lithophaga (Linnaeus, 1758) | 0.01 | ||
| Mytilidae | Modiolus barbatus (Linnaeus, 1758) | I | 0.01 | |
| Mytilidae | Musculus cf. costulatu (Risso, 1826) | I | 7.95 | 5.05 |
| Mytilidae | Musculus sp. (Röding, 1798) | I | 0.01 | 0.02 |
| Mytilidae | Mytilus galloprovincialis (Lamarck, 1819) | III | 62.19 | 77.32 |
| Noetiidae | Striarca lactea (Linnaeus, 1758) | I | 0.03 | 0.02 |
| Pectinidae | Flexopecten glaber (Linnaeus, 1758) | I | 0.01 | |
| Spondylidae | Spondylus sp. Linnaeus, 1758 | 0.01 | ||
| Veneridae | Clausinella sp. (Gray, 1851) | I | 0.01 | |
| Veneridae | Irus irus (Linnaeus, 1758) | I | 0.01 | 0.13 |
| Veneridae | Veneridae indet. 1 | I | 0.01 | 0.02 |
| Veneridae | Veneridae indet. 2 | I | 0.06 | |
| Veneridae | Veneridae indet. 3 | I | 0.05 | 0.08 |
| Veneridae | Veneridae indet. 4 | I | 0.01 | |
| Veneridae | Veneridae indet. 5 | I | 0.01 | |
| Veneridae | Veneridae indet. 6 | I | 0.01 | |
| POLYPLACOPHORA | ||||
| Acanthochitonidae | Acanthochitona fascicularis (Linnaeus, 1767) | I | 1.11 | 0.81 |
| Ecological Group | Share in the Total Number of Mollusks | |
|---|---|---|
| Without Anthropogenic Impact | With Anthropogenic Impact | |
| I | 24.83% | 14.02% |
| II | 0.10% | 0.62% |
| III | 62.19% | 77.32% |
| AMBI indexs | 1.87 | 2.33 |
| Gastropoda | |||||
|---|---|---|---|---|---|
| Source | Df | SS | MS | Pseudo-F | P(MC) |
| Corallina | 1 | 7458.7 | 7458.7 | 3.9186 | 0.0022 |
| Season = Se | 1 | 47,363 | 47,363 | 12.821 | 0.0002 |
| Anthropogenic impact = An | 1 | 4520.4 | 4520.4 | 0.85593 | 0.5389 |
| Area = Ar | 3 | 20,338 | 6779.3 | 2.784 | 0.0037 |
| Se × An | 1 | 4455.5 | 4455.5 | 2.3209 | 0.0822 |
| Se × Ar | 3 | 11,077 | 3692.2 | 1.3807 | 0.1898 |
| An × Ar | 3 | 15,143 | 5047.5 | 2.1675 | 0.0257 |
| Location (Se × Ar) | 8 | 19,251 | 2406.4 | 2.1989 | 0.0002 |
| Se × An × Ar | 3 | 5543.9 | 1848 | 0.71144 | 0.7675 |
| Se × Location (An × Ar) | 8 | 20,816 | 2602.1 | 2.3777 | 0.0001 |
| Error | 63 | 68,946 | 1094.4 | ||
| Bivalvia | |||||
|---|---|---|---|---|---|
| Source | Df | SS | MS | Pseudo-F | P(MC) |
| Corallina | 1 | 1451.7 | 1451.7 | 2.2751 | 0.078 |
| Season = Se | 1 | 8411.5 | 8411.5 | 4.656 | 0.0263 |
| Anthropogenic = An | 1 | 859.8 | 859.8 | 0.89527 | 0.511 |
| Area = Ar | 3 | 8668.5 | 2889.5 | 3.7172 | 0.0031 |
| Se × An | 1 | 1673.3 | 1673.3 | 1.4399 | 0.2881 |
| Se × Ar | 3 | 5473.5 | 1824.5 | 1.8771 | 0.0809 |
| An × Ar | 3 | 2775 | 924.99 | 1.2457 | 0.3078 |
| Location (An × Ar) | 8 | 6141.8 | 767.73 | 2.2408 | 0.0009 |
| Se × An × Ar | 3 | 3350.9 | 1117 | 1.1873 | 0.343 |
| Se × Location (An × Ar) | 8 | 7540.9 | 942.61 | 2.7512 | 0.0001 |
| Error | 63 | 21585 | 342.62 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buršić, M.; Iveša, L.; Jaklin, A.; Arko Pijevac, M.; Bruvo Mađarić, B.; Neal, L.; Pustijanac, E.; Burić, P.; Iveša, N.; Paliaga, P. Changes in Composition of Mollusks within Corallina officinalis Turfs in South Istria, Adriatic Sea, as a Response to Anthropogenic Impact. Diversity 2023, 15, 939. https://doi.org/10.3390/d15080939
Buršić M, Iveša L, Jaklin A, Arko Pijevac M, Bruvo Mađarić B, Neal L, Pustijanac E, Burić P, Iveša N, Paliaga P. Changes in Composition of Mollusks within Corallina officinalis Turfs in South Istria, Adriatic Sea, as a Response to Anthropogenic Impact. Diversity. 2023; 15(8):939. https://doi.org/10.3390/d15080939
Chicago/Turabian StyleBuršić, Moira, Ljiljana Iveša, Andrej Jaklin, Milvana Arko Pijevac, Branka Bruvo Mađarić, Lucija Neal, Emina Pustijanac, Petra Burić, Neven Iveša, and Paolo Paliaga. 2023. "Changes in Composition of Mollusks within Corallina officinalis Turfs in South Istria, Adriatic Sea, as a Response to Anthropogenic Impact" Diversity 15, no. 8: 939. https://doi.org/10.3390/d15080939
APA StyleBuršić, M., Iveša, L., Jaklin, A., Arko Pijevac, M., Bruvo Mađarić, B., Neal, L., Pustijanac, E., Burić, P., Iveša, N., & Paliaga, P. (2023). Changes in Composition of Mollusks within Corallina officinalis Turfs in South Istria, Adriatic Sea, as a Response to Anthropogenic Impact. Diversity, 15(8), 939. https://doi.org/10.3390/d15080939