
Optimizing Growth and Rearing Techniques for Larvae and Juveniles of the Sea Cucumber Holothuria arguinensis


 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Spawning Induction
2.2. Larval Development and Settlement
2.3. Juvenile Growth
2.4. Statistical Analyses
3. Results
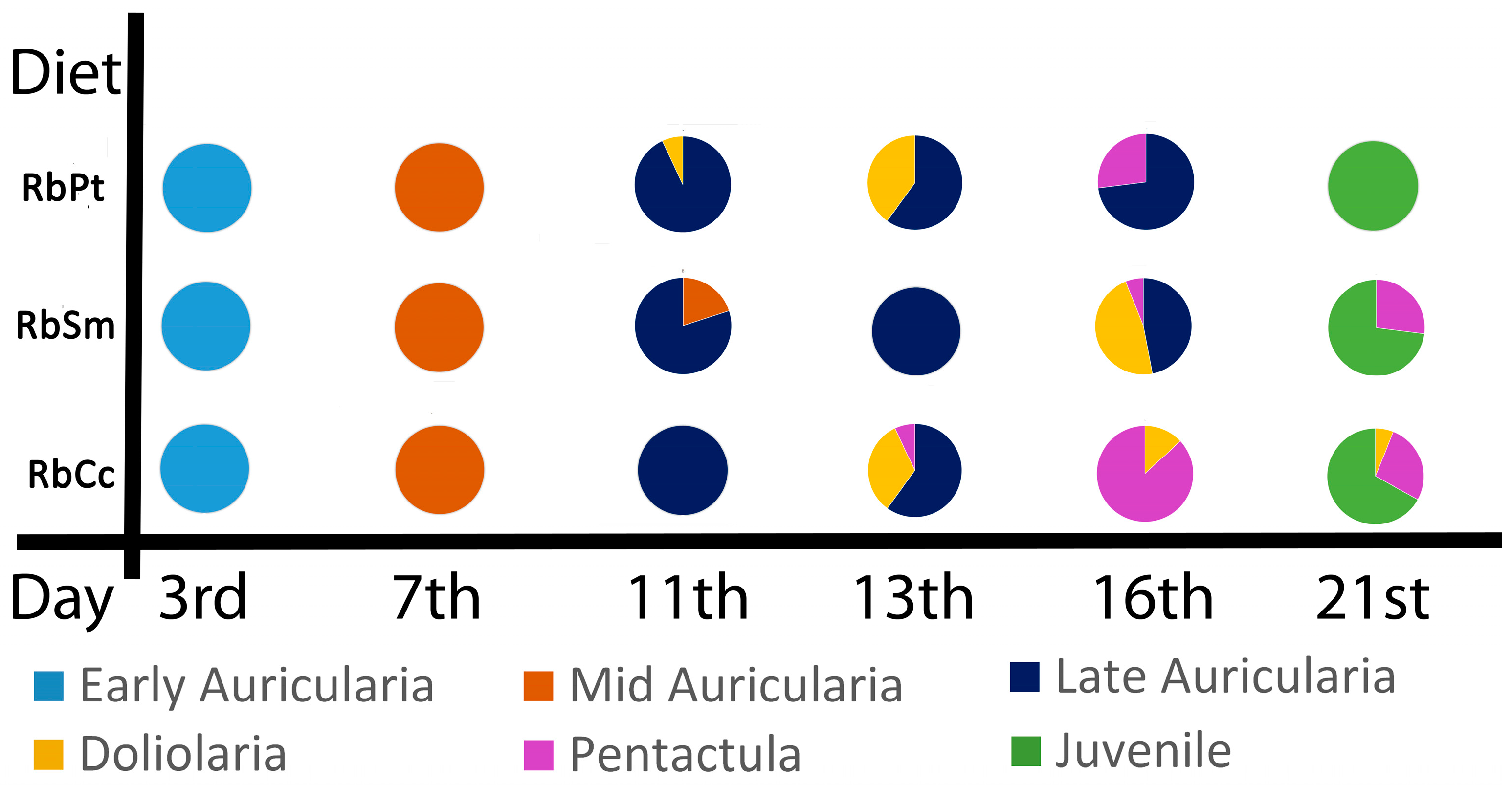
3.1. Larval Development
3.2. Larval Settlement and Survival
3.3. Juvenile Growth
4. Discussion
4.1. Larval Development
4.2. Larval Survival and Settlement
4.3. Juvenile Rearing and Growth
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, X.; McCoy, S.J.; Du, Y.; Widdicombe, S.; Hall-Spencer, J.M. Physiological and Behavioral Plasticity of the sea cucumber Holothuria forskali (Echinodermata, Holothuroidea) to Acidified Seawater. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Purcell, S.W. Value, market preferences and trade of beche-de-mer from Pacific Island sea cucumbers. PLoS ONE 2014, 9, e95075. [Google Scholar] [CrossRef]
- FAO. Sea Cucumber: A Global Review of Fisheries and Trade; Fisheries and Aquaculture Technical Paper. No. 516; FAO: Rome, Italy, 2008. [Google Scholar]
- Purcell, S.W.; Mercier, A.; Conand, C.; Hamel, J.; Toral-Granda, M.V.; Lovatelli, A.; Uthicke, S. Sea cucumber fisheries: Global analysis of stocks, management measures and drivers of overfishing. Fish Fish. 2011, 14, 34–59. [Google Scholar] [CrossRef]
- Conand, C.; Preston, G.; Hoffschir, C. Trends in tropical sea-cucumber fisheries. In Biology of Echinodermata; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar] [CrossRef]
- Dereli, H.; Haydin, M. Sea cucumber fishery in Turkey: Management regulations and their efficiency. Reg. Stud. Mar. Sci. 2020, 4, 101551. [Google Scholar] [CrossRef]
- Hair, C.; Mills, D.J.; McIntyre, R.; Southgate, P.C. Optimising methods for community-based sea cucumber ranching: Experimental releases of cultured juvenile Holothuria scabra into seagrass meadows in Papua New Guinea. Aquacult. Rep. 2016, 3, 198e208. [Google Scholar] [CrossRef]
- Han, Q.; Keesing, J.K.; Liu, D. A review of sea cucumber aquaculture, ranching, and stock enhancement in China. Rev. Fish Sci. Aquac. 2016, 24, 326–341. [Google Scholar] [CrossRef]
- Gianasi, B.L.; Hamel, J.H.; Montgomery, E.M.; Sun, J.; Mercier, A. Current Knowledge on the Biology, Ecology, and Commercial Exploitation of the Sea Cucumber Cucumaria frondose. Rev. Fish. Sci. Aquac. 2020, 29, 582–653. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Huang, W.; Huo, D.; Yu, Z.; Ren, C.; Jiang, X.; Luo, P.; Chen, T.; Hu, C. Spawning, larval development and juvenile growth of the tropical sea cucumber Holothuria leucospilota. Aquaculture 2018, 488, 22–29. [Google Scholar] [CrossRef]
- Rakaj, A.; Fianchini, A.; Boncagni, P.; Scardi, M.; Cataudella, S. Artificial reproduction of Holothuria polii: A new candidate for aquaculture. Aquaculture 2019, 498, 444–453. [Google Scholar] [CrossRef]
- Laguerre, H.; Raymond, G.; Plan, P.; Améziane, N.; Bailly, X.; Le Chevalier, P. First description of embryonic and larval development, juvenile growth of the black sea-cucumber Holothuria forskali (Echinodermata: Holothuroidea), a new species for aquaculture in the north-eastern Atlantic. Aquaculture 2020, 521, 734961. [Google Scholar] [CrossRef]
- Juinio-Meñez, M.A.; Tech, E.D.; Ticao, I.P.; Gorospe, J.R.C.; Edullantes, C.M.A.; Rioja, R.A.V. Adaptive and integrated culture production systems for the tropical sea cucumber Holothuria scabra. Fish. Res. 2017, 186, 502–513. [Google Scholar] [CrossRef]
- Tuwo, A.; Yasir, I.; Syafiuddin; Aprianto, R.; Yanti, A.; Bestari, A.; Tresnati, J. Low salinity reduces survival rate of a commercially important sea cucumber (Sandfish: Holothuria scabra). In Proceedings of the IOP Conference Series: Earth and Environmental Science, Changsha, China, 18–20 September 2020. [Google Scholar] [CrossRef]
- Venâncio, E.; Félix, P.M.; Brito, A.C.; Sousa, J.; Azevedo e Silva, F.; Simões, T.; Narciso, L.; Amorim, A.; Dâmaso, L.; Pombo, A. Do broodstock diets influence viability and larval development of Holothuria mammata? Aquaculture 2021, 536, 736431. [Google Scholar] [CrossRef]
- Domínguez-Godino, J.A.; González-Wangüemert, M. Holothuria arguinensis: As new sea cucumber species for aquaculture. SPC Beche-De-Mer Inf. Bull. 2019, 39, 60–64. [Google Scholar]
- Godino, J.A.; Slater, M.J.; Hannon, C.; González-Wangüermert, M. A New Species for Sea Cucumber Ranching and Aquaculture: Breeding and Rearing of Holothuria Arguinensis. Aquaculture 2015, 438, 122–128. [Google Scholar] [CrossRef]
- Domínguez-Godino, J.A.; González-Wangüemert, M. Habitat associations and seasonal abundance patterns of the sea cucumber Holothuria arguinensis at Ria Formosa coastal lagoon (South Portugal). Aquat. Ecol. 2020, 54, 337–354. [Google Scholar] [CrossRef]
- González-Wangüemert, M.; Domínguez-Godino, J.A.; Cánovas, F. The fast development of sea cucumber fisheries in the Mediterranean and NE Atlantic waters: From a new marine resource to its over-exploitation. Ocean. Coast. Manag. 2017, 151, 165–177. [Google Scholar] [CrossRef]
- Domínguez-Godino, J.A.; González-Wangüemert, M. Improving the fitness of Holothuria arguinensis larvae through different microalgae diets. Aquac. Res. 2019, 50, 3130–3137. [Google Scholar] [CrossRef]
- Gianasi, B.L.; Jean-François Hamel, J.F.; Mercier, A. Morphometric and behavioural changes in the early life stages of the sea cucumber Cucumaria frondose. Aquaculture 2018, 490, 5–18. [Google Scholar] [CrossRef]
- Domínguez-Godino, J.A.; Santos, T.F.; Pereira, H.; Custódio, L.; González-Wangüemert, M. Seagrass debris as potential food source to enhance Holothuria arguinensis’ growth in aquaculture. Aquac. Res. 2020, 51, 1487–1499. [Google Scholar] [CrossRef]
- Ru, X.; Zhang, L.; Li, X.; Liu, S.; Yang, H. Development strategies for the sea cucumber industry in China. J. Oceanol. Limnol. 2019, 37, 300–312. [Google Scholar] [CrossRef]
- Nora’aini, A.; Wahab, M.A.; Jusoh, A.; Hasan, M.R.; Ghazali, N.; Kamaruzaman, K. Treatment of aquaculture wastewater using ultra-low pressure asymmetric polyethersulfone (PES) membrane. Desalination 2005, 185, 317–326. [Google Scholar] [CrossRef]
- Lee, J.S.F.; Pierce, M.L.; Cook, M.A.; Berejikian, B.A.; Goetz, F.W.; Poretsky, R.S. Effects of tank cleaning frequency and sea cucumber co-culture on larval sablefish growth and survival, water quality and microbial communities. Aquaculture 2021, 545, 737213. [Google Scholar] [CrossRef]
- Cheng, C.; Wu, F.; Ren, C.; Jiang, X.; Zhang, X.; Li, X.; Peng, L.; Hu, C.; Chen, T. Aquaculture of the tropical sea cucumber Stichopus monotuberculatus: Induced spawning, detailed records of gonadal and embryonic development, and improvements in larval breeding by digestive enzyme supply in diet. Aquaculture 2021, 540, 736690. [Google Scholar] [CrossRef]
- Rakaj, A.; Fianchini, A.; Boncagni, P.; Lovatelli, A.; Scardi, M.; Cataudella, S. Spawning and rearing of Holothuria tubulosa: A new candidate for aquaculture in the Mediterranean region. Aquac. Res. 2017, 49, 557–568. [Google Scholar] [CrossRef]
- Domínguez-Godino, J.A.; González-Wangüemert, M. Breeding and larval development of Holothuria mammata, a new target species for aquaculture. Aquac. Res. 2018, 49, 578–1430. [Google Scholar] [CrossRef]
- Yamamoto, S.; Okauchi, M.; Yoshimatsu, T. Dietary value of microalga Rhodomonas sp. as a live food for sea cucumber Apostichopus japonicus larvae. Nippon. Suisan Gakkaishi 2015, 81, 973–978. [Google Scholar] [CrossRef][Green Version]
- Sibonga, R.C.; Laureta, L.V.; Lebata-Ramos, M.J.H.; Nievales MF, J.; Pedroso, F.L. Single and mixed species of microalgae as larval food for the tropical sea cucumber Holothuria scabra. J. Appl. Phycol. 2021, 33, 3103–3112. [Google Scholar] [CrossRef]
- Agudo, N. Sandfish Hatchery Techniques; Australian Centre for International Agricultural Research (ACIAR), Secretariat of the Pacific Community (SPC) & WorldFish Center: Noumea, Australia, 2006; p. 43. [Google Scholar]
- Santos, M.; Amorim, A.; Cruz, J.P.C.; Veloso, V.; Favareto, L.R.; Dâmaso-Rodrigues, M.L.; Palma, C.; Borges, C.; Chainho, P.; Félix, P.M.; et al. Phytoplankton community in the Sado estuary. In Proceedings of the Actas das 6as Jornadas de Engenharia Hidrográfica/1as Jornadas Luso-Espanholas de Hidrografia, Lisboa, Portugal, 3–5 November 2020; pp. 247–250. [Google Scholar]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Future Microbiol. 2002, 5, 917–933. [Google Scholar] [CrossRef]
- López, Y.; Soto, S.M. The Usefulness of Microalgae Compounds for Preventing Biofilm Infections. Antibiotics 2019, 9, 9. [Google Scholar] [CrossRef]
- van Houcke, J.; Medina, I.; Maehre, H.K.; Cornette, J.; Cardinal, M.; Linssen, J.; Luten, J. The effect of algae diets (Skeletonema costatum and Rhodomonas baltica) on the biochemical composition and sensory characteristics of Pacific cupped oysters (Crassostrea gigas) during land-based refinement. Food Res. Int. 2017, 100, 151–160. [Google Scholar] [CrossRef]
- Chronakis, I.; Madsen, M. Algal proteins. In Handbook of Food Proteins; Woodhead Publishing Series in Food Sciences, Technology and Nutrition; Elsevier: Cambridge, UK, 2011. [Google Scholar] [CrossRef]
- Bertozzini, E.; Galluzzi, L.; Ricci, F.; Penna, A.; Magnani, M. Neutral lipid content and biomass production in Skeletonema marinoi (Bacillariophyceae) culture in response to nitrate limitation. Appl. Biochem. Biotechnol. 2013, 170, 1624–1636. [Google Scholar] [CrossRef]
- Wang, X.; Fosse, H.K.; Li, K.; Chauton, M.S.; Vadstein, O.; Reitan, K.I. Influence of Nitrogen Limitation on Lipid Accumulation and EPA and DHA Content in Four Marine Microalgae for Possible use in Aquafeed. Front. Mar. Sci. 2019, 6, 95. [Google Scholar] [CrossRef]
- Mitani, E.; Nakayama, F.; Matsuwaki, I.; Ichi, I.; Kawabata, A.; Kawachi, M.; Kato, M. Fatty acid composition profiles of 235 strains of three microalgal divisions within the NIES Microbial Culture Collection. Microb. Resour. Syst. 2017, 33, 19–29. [Google Scholar]
- Napolitano, G.E.; Ackman, R.G.; Ratnayake, W.M.N. Fatty Acid Composition of Three Cultured Algal Species (Isochvysis galbana, Chaetoceros gracilis and Chaetoceros calcitrans) Used as Food for Bivalve Larvae. J. World Aquac. Soc. 1990, 21, 122–130. [Google Scholar] [CrossRef]
- Félix, P.M.; Pombo, A.; Azevedo e Silva, F.; Simões, T.; Marques, T.A.; Melo, R.; Rocha, C.; Sousa, J.; Venâncio, E.; Costa, J.L.; et al. Modelling the Distribution of a Commercial NE-Atlantic Sea Cucumber, Holothuria mammata: Demographic and Abundance Spatio-Temporal Patterns. Front. Mar. Sci. 2021, 8, 675330. [Google Scholar] [CrossRef]
- Rodrigues, T.; Azevedo e Silva, F.; Sousa, J.; Félix, P.M.; Pombo, A. Effect of Enriched Substrate on the Growth of the Sea Cucumber Holothuria arguinensis Koehler and Vaney, 1906 Juveniles. Diversity 2023, 15, 458. [Google Scholar] [CrossRef]
- Tweedie, M.C.K. An index which distinguishes between some important exponential families. In Statistics: Applications and New Directions: Proceedings of the Indian Statistical Institute Golden Jubilee International Conference, Calcutta, India; Ghosh, J.K., Roy, J., Eds.; Indian Statistical Institute: Calcutta, India, 1984; pp. 579–604. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: https://www.r-project.org/ (accessed on 12 March 2023).
- Battaglene, S.C. Culture of tropical sea cucumbers for stock restoration and enhancement. Naga ICLARM Q. 1999, 22, 4–11. [Google Scholar]
- Restrepo, J.; González-Wangüemert, M.; Erzini, K. Estimation of growth parameters for the exploited sea cucumber Holothuria arguinensis from South Portugal. Fish. Bull. Natl. Ocean. Atmos. Adm. 2018, 116, 1–8. [Google Scholar] [CrossRef]
- Ribalet, F.; Vidoudez, C.; Cassin, D.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. High plasticity in the production of diatom-derived polyunsaturated aldehydes under nutrient limitation: Physiological and ecological implications. Protist 2009, 160, 444–451. [Google Scholar] [CrossRef]
- Banerjee, S.; Hew, W.; Khatoon, H.; Shariff, M.; Yusoff, F. Growth and proximate composition of tropical marine Chaetoceros calcitrans and Nannochloropsis oculata cultured outdoors and under laboratory conditions. Afr. J. Biotechnol. 2011, 10, 1375–1383. [Google Scholar] [CrossRef]
- Duy ND, Q.; Francis, D.S.; Southgate, P.C. Development of hyaline spheres in late auriculariae of sandfish, Holothuria scabra: Is it a reliable indicator of subsequent performance? Aquaculture 2016, 461, 144–151. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, L.; Xia, S.; Liu, S.; Ru, X.; Xu, Q.; Zhang, T.; Yang, H. Effects of dietary protein levels on the growth, energy budget, and physiological and immunological performance of green, white and purple color morphs of sea cucumber, Apostichopus japonicus. Aquaculture 2016, 450, 375–382. [Google Scholar] [CrossRef]
- Gianasi, B.L.; Parrish, C.C.; Hamel, J.F.; Mercier, A. Influence of diet on growth, reproduction and lipid and fatty acid composition in the sea cucumber Cucumaria frondosa. Aquac. Res. 2017, 48, 3413–3432. [Google Scholar] [CrossRef]
- Li, L.; Li, Q. Effects of stocking density, temperature, and salinity on larval survival and growth of the red race of the sea cucumber Apostichopus japonicus (Selenka). Aquacult. Int. 2010, 18, 447–460. [Google Scholar] [CrossRef]
- Ito, S.; Kitamura, H. Induction of larval metamorphosis in the sea cucumber Stichopus japonicus by periphitic diatoms. Hydrobiologica 1997, 358, 281–284. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Kong, L. The Effect of Different Substrates on Larvae Settlement in Sea Cucumber, Apostichopus japonicus Selenka. J. World Aquac. Soc. 2010, 41, 123–130. [Google Scholar] [CrossRef]
- Roca, C.; Alves, V.D.; Freitas, F.; Reis, M.A. Exopolysaccharides enriched in rare sugars: Bacterial sources, production, and applications. Front. Microbiol. 2015, 6, 288. [Google Scholar] [CrossRef]
- Mercier, A.; Battaglene, S.C.; Hamel, J. Settlement preferences and early migration of the tropical sea cucumber Holothuria scabra. J. Exp. Mar. Biol. Ecol. 2000, 249, 89–110. [Google Scholar] [CrossRef]
- Joyce, A.; Utting, S. The role of exopolymers in hatcheries: An overlooked factor in hatchery hygiene and feed quality. Aquaculture 2015, 446, 122–131. [Google Scholar] [CrossRef]
- Slater, M.J.; Jeffs, A.G.; Carton, A.G. The use of the waste from green-lipped mussels as a food source for juvenile sea cucumber, Australostichopus mollis. Aquaculture 2009, 292, 219–224. [Google Scholar] [CrossRef]
- Garcia, J.S.; Palacios, V.; Roldán, A. Nutritional potential of four seaweed species collected in the barbate estuary (Gulf of Cadiz, Spain). J. Nutr. Food Sci. 2016, 6, 1–7. [Google Scholar]
- Sánchez-Machado, D.I.; López-Cervantes, J.; Lopez-Hernandez, J.; Paseiro-Losada, P. Fatty acids, total lipid, protein and ash contents of processed edible seaweeds. Food Chem. 2004, 85, 439–444. [Google Scholar] [CrossRef]
- Bruckner, A.W.; Johnson, K.A.; Field, J.D. Conservation strategies for sea cucumbers: Can a CITES Appendix II listing promote sustainable international trade. SPC Bêche De Mer. Inf. Bull. 2003, 18, 24–33. [Google Scholar]
- Preston, G.L. Beche-de-mer. In Nearshore Marine Resources of the South Pacific: Information for Fisheries Development and Management; Forum Fisheries Agency: Honiara, Solomon Islands, 1993; pp. 371–401. [Google Scholar]
- Uthicke, S.; Karez, R. Sediment patch selectivity in tropical sea cucumbers (Holothurioidea: Aspidochirotida) analysed with multiple choice experiments. J. Exp. Mar. Biol. Ecol. 1999, 236, 69–87. [Google Scholar] [CrossRef]
- Asha, P.S.; Muthiah, P.S. Growth of the hatchery-produced juveniles of commercial sea cucumber Holothuria (Theelothuria) spinifera Theel. Aquac. Res. 2007, 38, 1082–1087. [Google Scholar] [CrossRef]
- Yang, H.; Yuan, X.; Zhou, Y.; Mao, Y.; Zhang, T.; Liu, Y. Effects of body size and water temperature on food consumption and growth in the sea cucumber Apostichopus japonicus (Selenka) with special reference to aestivation. Aquac. Res. 2005, 36, 1085–1092. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | RbPt | RbSm | RbCc | Source |
|---|---|---|---|---|
| Protein | 65% | 49% | 72% | [36,37,38] |
| Lipids | 28% | 16% | 24% | [36,37,38,39] |
| EPA * | 12% | 7% | 14% | [36,37,38,39,40,41] |
| DHA * | 3% | 3% | 3% | [36,37,38,39,40,41] |
| ARA * | 2% | 1% | <1% | [36,37,38,39,40,41] |
| ∑PUFA * | 36% | 41% | 47% | [36,37,38,39,40,41] |
| Larval Stage | Measurement | Diet RbPt | Diet RbSm | Diet RbCc | Statistical Analysis |
|---|---|---|---|---|---|
| Mid Auricularia | Length | 468 ± 74 ab | 376 ± 179 a | 543 ± 84 b | [F(2, 46) = 7.1906; p = 0.0020; Tukey < 0.01] |
| Width | 255 ± 57 ab | 217 ± 105 a | 301 ± 75 b | [F(2, 46) = 4.1700; p = 0.0218; Tukey < 0.05] | |
| Late Auricularia | Length | 628 ± 91 | 672 ± 104 | 655 ± 82 | [F(2, 83) = 1.8811; p = 0.1589] |
| Width | 407 ± 70 | 451 ± 72 | 409 ± 61 | [F(2, 83) = 3.9863; p = 0.2230] | |
| Doliolaria | Length | 471 ± 78 | 369 ± 40 | 520 ± 155 | [F(2, 27) = 3.2094; p = 0.0568] |
| Width | 291 ± 56 | 253 ± 32 | 327 ± 93 | [F(2, 27) = 2.1315; p = 0.1389] | |
| Pentactula | Length | 317 ± 11 | 312 ± 27 | 294 ± 25 | [F(2, 24) = 2.2184; p = 0.1315] |
| Width | 180 ± 17 ab | 202 ± 21 b | 171 ± 17 a | [F(2, 24) = 5.3836; p = 0.0121; Tukey < 0.01] | |
| Juvenile | Length | 356 ± 52 | 299 ± 33 | 319 ± 86 | [F(2, 34) = 3.0223; p = 0.0623] |
| Width | 264 ± 43 b | 208 ± 20 a | 201 ± 52 a | [F(2, 34) = 9.0518; p = 0.0007; Tukey < 0.01] |
| β | Std. Error | t-Value | p-Value | |
|---|---|---|---|---|
| (Intercept) | 2.38863 | 0.35585 | 6.713 | 4.16 × 109 *** |
| Depth mid (18 cm) | −0.08683 | 0.47292 | −0.184 | 0.854848 |
| Depth upper (9 cm) | −1.17600 | 0.56863 | −2.068 | 0.042323 * |
| Diet RbSm | −0.45669 | 0.26391 | −1.730 | 0.087951 |
| Diet RbCc | −0.84246 | 0.28181 | −2.990 | 0.003853 ** |
| Substrate Plaque | −0.69056 | 0.52075 | −1.326 | 0.189121 |
| Substrate Roll | 0.22293 | 0.45273 | 0.492 | 0.623973 |
| Depth mid: Substrate Plaque | −0.46939 | 0.78038 | −0.601 | 0.549456 |
| Depth upper: Substrate Plaque | 2.59900 | 0.74636 | 3.482 | 0.000861 *** |
| Depth mid: Substrate Roll | 0.31581 | 0.63537 | 0.497 | 0.620711 |
| Depth upper: Substrate Roll | 0.97460 | 0.72592 | 1.343 | 0.183750 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, J.; Félix, P.M.; Brito, A.C.; Venâncio, E.; Azevedo e Silva, F.; Simões, T.; Amorim, A.; Dâmaso-Rodrigues, M.L.; Pombo, A. Optimizing Growth and Rearing Techniques for Larvae and Juveniles of the Sea Cucumber Holothuria arguinensis. Diversity 2023, 15, 722. https://doi.org/10.3390/d15060722
Sousa J, Félix PM, Brito AC, Venâncio E, Azevedo e Silva F, Simões T, Amorim A, Dâmaso-Rodrigues ML, Pombo A. Optimizing Growth and Rearing Techniques for Larvae and Juveniles of the Sea Cucumber Holothuria arguinensis. Diversity. 2023; 15(6):722. https://doi.org/10.3390/d15060722
Chicago/Turabian StyleSousa, João, Pedro M. Félix, Ana C. Brito, Eliana Venâncio, Francisco Azevedo e Silva, Tomás Simões, Ana Amorim, Maria Luísa Dâmaso-Rodrigues, and Ana Pombo. 2023. "Optimizing Growth and Rearing Techniques for Larvae and Juveniles of the Sea Cucumber Holothuria arguinensis" Diversity 15, no. 6: 722. https://doi.org/10.3390/d15060722
APA StyleSousa, J., Félix, P. M., Brito, A. C., Venâncio, E., Azevedo e Silva, F., Simões, T., Amorim, A., Dâmaso-Rodrigues, M. L., & Pombo, A. (2023). Optimizing Growth and Rearing Techniques for Larvae and Juveniles of the Sea Cucumber Holothuria arguinensis. Diversity, 15(6), 722. https://doi.org/10.3390/d15060722