

Spatial and Temporal Variations in Autumn Fish Assemblages in the Offshore Waters of the Yangtze Estuary



Abstract
1. Introduction
2. Materials and Methods
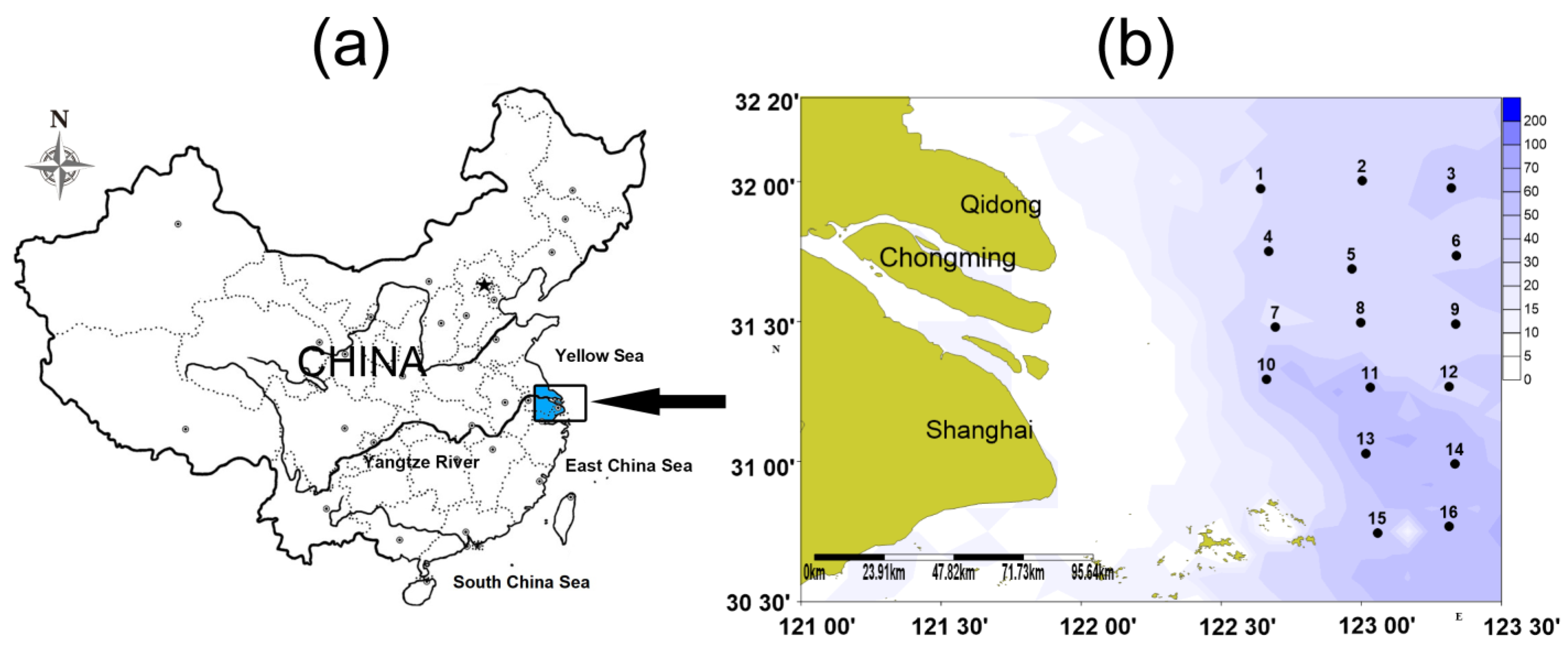
2.1. Sample Collection
2.2. Data Analysis
3. Results
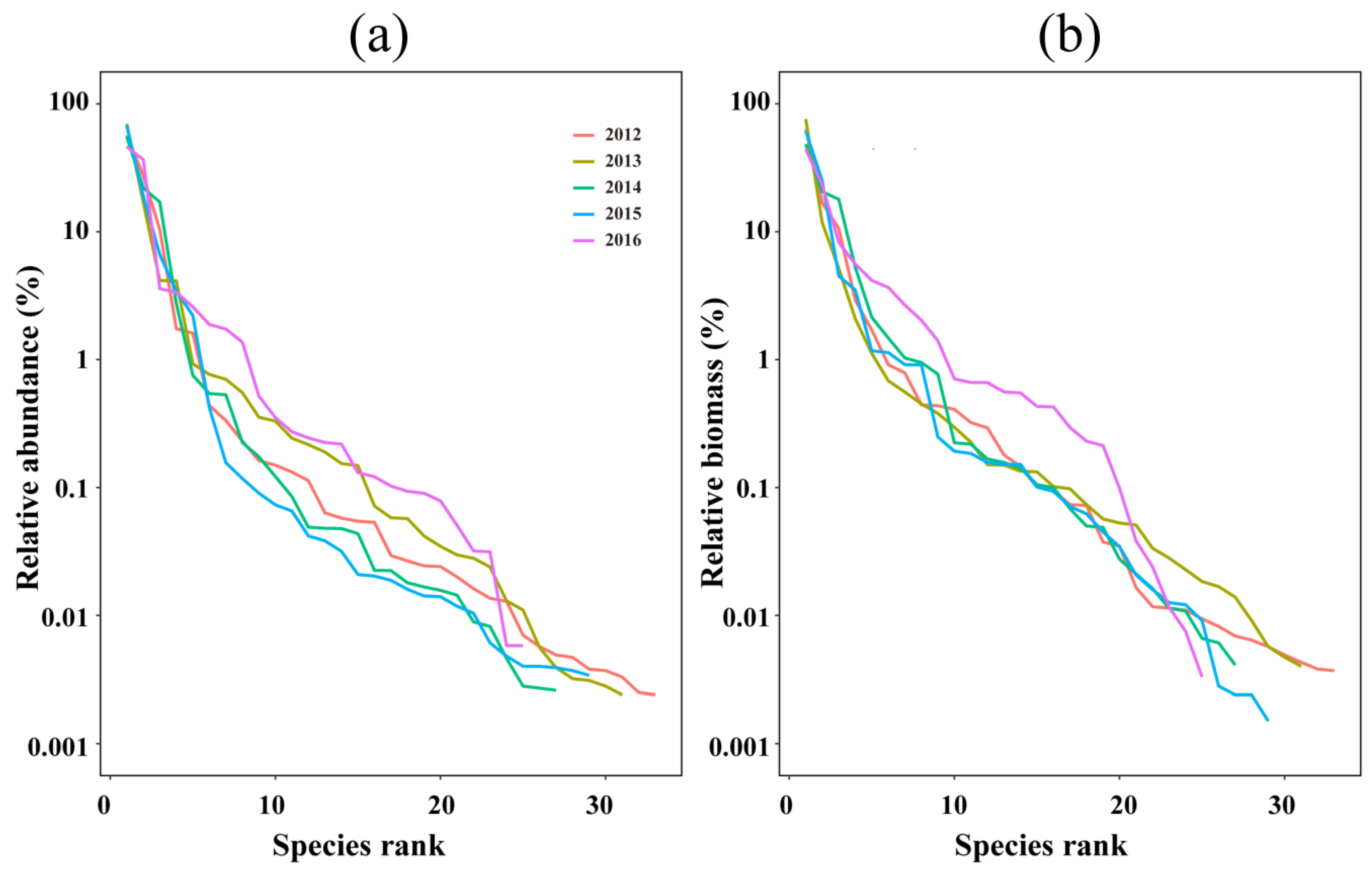
3.1. Faunal Composition
3.2. Environment Variables
3.3. Annual Variation in Fish Assemblages and Relationships with Environmental Factors
3.4. Spatial Characteristics of the Fish Assemblages
4. Discussion
4.1. Composition of the Fish Assemblages
4.2. Annual Variation in Fish Assemblages
4.3. Spatial Characteristics of the Fish Assemblages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.Q.; Xian, W.W.; Li, W.L. Fish Assemblage Structure in Adjacent Sea of Changjiang Estuary in Spring of 2004 and 2007 and Its Association with Environmental Factors. Period. Ocean Univ. China 2013, 43, 67–74. [Google Scholar]
- Elliott, M. The forth estuary: A nursery and overwintering area for North Sea fishes. Hydrobiologia 1990, 195, 89–103. [Google Scholar] [CrossRef]
- Wasserman, R.J.; Strydom, N.A. The importance of estuary head waters as nursery areas for young estuary– and marine–spawned fishes in temperate South Africa. Estuar. Coast. Shelf Sci. 2011, 94, 56–67. [Google Scholar] [CrossRef]
- Sheaves, M.; Johnston, R.; Connolly, R.M.; Baker, R. Importance of estuarine mangroves to juvenile banana prawns. Estuar. Coast. Shelf Sci. 2012, 114, 208–219. [Google Scholar] [CrossRef]
- Shao, H.B. Research on Composition, Distribution and Influencing Factors of Suspended Matters in the Yangtze River Estuary in Fall. Master’s Thesis, Ocean University of China, Qingdao, China, 2012. [Google Scholar]
- Luo, B.Z.; Shen, H.T. The Three Gorges Project and the Ecological Environment of Yangtze Estuary; Science Press: Beijing, China, 1994; pp. 1–32. [Google Scholar]
- Xin, M. Long–Term Variations of the Key Environmental Factors and Their Ecological Effects in the Changjiang Estuary. Ph.D. Thesis, Ocean University of China, Qingdao, China, 2014. [Google Scholar]
- Vaquer-Sunyer, R.; Duarte, C.M. Thresholds of hypoxia for marine biodiversity. Proc. Natl. Acad. Sci. USA 2008, 105, 15452–15457. [Google Scholar] [CrossRef]
- Meier, H.E.M.; Eilola, K.; Almroth-Rosell, E.; Schimanke, S.; Kniebusch, M.; Hoglund, A.; Pemberton, P.; Liu, Y.; Vali, G.; Saraiva, S. Disentangling the impact of nutrient load and climate changes on Baltic Sea hypoxia and eutrophication since 1850. Clim. Dynam. 2019, 53, 1145–1166. [Google Scholar] [CrossRef]
- Zhuang, P. Fishes of the Yangtze Estuary; China Agriculture Press: Beijing, China, 2018. [Google Scholar]
- Shen, X.Q.; Shi, Y.R.; Chao, M.; Quan, W.M.; Huang, H.J.; Wu, Q.Y. Analysis of taxonomic diversity of fish community in Yangtze River estuary. Prog. Mater. Sci. 2013, 34, 1–7. [Google Scholar]
- Wu, J.H.; Wang, J.Q.; Dai, X.J.; Tian, S.Q.; Liu, J.; Chen, J.H.; Wang, X.F. An analysis of spatial co–occurrence pattern of fish species of Yangtze River estuary based on probabilistic model. South China Fish. Sci. 2019, 15, 1–9. [Google Scholar]
- Wang, Y.C.; Liang, C.; Xian, W.W.; Wang, Y.B. Using the LBB Method for the Assessments of Seven Fish Stocks From the Yangtze Estuary and Its Adjacent Waters. Front. Mar. Sci. 2021, 8, 679299. [Google Scholar] [CrossRef]
- Liu, S.H.; Yang, Y.Y.; He, Y.L.; Ji, X.; Wang, Y.T.; Zhang, H.J.; Mao, R.Y.; Jiang, X.S.; Cheng, X.S. Morphological classification of ichthyoplankton in the Changjiang River Estuary based on DNA barcoding. Acta. Oceanol. Sin. 2021, 43, 93–104. [Google Scholar]
- Yu, H.C.; Xian, W.W. The environment effect on fish assemblage structure in waters adjacent to the Changjiang (Yangtze) River estuary (1998–2001). Chin. J. Oceanol. Limnol. 2009, 327, 443–456. [Google Scholar] [CrossRef]
- Zhang, H.; Xian, W.W.; Liu, S.D. Ichthyoplankton assemblage structure of springs in the Yangtze Estuary revealed by biological and environmental visions. PeerJ 2015, 3, e1186. [Google Scholar] [CrossRef]
- Li, J.S.; Li, S.F.; Cheng, J.H. Analysis on the annual variations of fish resources in the offshore water of Yangtze Estuary in autumn. Mar. Fish. 2008, 2, 120–125. [Google Scholar]
- Chen, Z.M.; Ren, Q.Q.; Liu, C.L.; Xian, W.W. Seasonal and Spatial Variations in Fish Assemblage in the Yangtze Estuary and Adjacent Waters and Their Relationship with Environmental Factors. J. Mar. Sci. Eng. 2022, 10, 1679. [Google Scholar] [CrossRef]
- Almeida, C.; Coelho, R.; Silva, M.; Bentes, L.; Monteiro, P.; Ribeiro, J.; Erzini, K.; Goncalves, J.M.S. Use of different intertidal habitats by faunal communities in a temperate coastal lagoon. Estuar. Coast. Shelf Sci. 2008, 80, 357–364. [Google Scholar] [CrossRef]
- Clarke, K.; Warwick, R. Similarity-Based testing for community pattern—The 2-way layout with no replication. Mar. Biol. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Chu, C.; Jones, N.E. Spatial variability of thermal regimes and other environmental determinants of stream fish communities in the Great Lakes Basin, Ontario, Canada. River Res. Appl. 2011, 27, 646–662. [Google Scholar] [CrossRef]
- Chen, Y.L. Spatio–Temporal Variation of Fishery Resources in the Yellow Sea and Yangtze River Estuary. Ph.D. Thesis, Institute of Oceanography, Chinese Academy of Sciences, Qingdao, China, 2017. [Google Scholar]
- Yang, D.; Wu, G.; Sun, J. The investigation ok pelagic eggs, larvae and juveniles of fishes at the mouth of the Changjiang river and adjacent areas. Oceanol. Limnol. Sin. 1990, 4, 346–355. [Google Scholar]
- Wang, Y.B.; Liang, C.; Chen, Z.M.; Liu, S.D.; Zhang, H.; Xian, W.W. Spring Ichthyoplankton Assemblage Structure in the Yangtze Estuary Under Environmental Factors. Front. Mar. Sci. 2021, 8, 806096. [Google Scholar] [CrossRef]
- Whitfield, A.K. Fish species diversity in Southern African estuarine systems: An evolutionary perspective. Environ. Biol. Fishes 1994, 40, 37–48. [Google Scholar] [CrossRef]
- Zheng, L.; Zhen, B.; Li, F.; Xu, B.Q.; Li, M.; Yang, F.Z. Comparison of ontaxonomic diversity of fish community among the Yellow River estuary, Yangtze River estuary, Pearl River estuary and their adjacent waters. J. Dalian Ocean Univ. 2014, 29, 530–535. [Google Scholar]
- Sreekanth, G.B.; Jaiswar, A.K.; Zacharia, P.U.; Pazhayamadom, D.G.; Chakraborty, S.K. Effect of environment on spatio–temporal structuring of fish assemblages in a monsoon–influenced tropical estuary. Environ. Monit. Assess. 2019, 191, 305. [Google Scholar] [CrossRef] [PubMed]
- Drake, P.; Arias, A.M.; Baldó, F.; Cuesta, J.A.; Rodríguez, A.; Silva–Garcia, A.; Sobrino, I.; García–González, D.; Fernández–Delgado, C. Spatial and temporal variation of the nekton and hyperbenthos from a temperate European estuary with regulated freshwater inflow. Estuaries 2002, 25, 451. [Google Scholar] [CrossRef]
- Jackson, G.; Jones, G. Spatial and temporal variation in nearshore fish and macroinvertebrate assemblages from a temperate Australian estuary over a decade. Mar. Ecol. Prog. Ser. 1999, 182, 253–268. [Google Scholar] [CrossRef]
- Luo, B.Z. Ecological characteristics and fishery resources of the Yangtze River Estuary and adjacent sea. Resour. Environ. Yangtze Val. 1992, 1, 24–30. [Google Scholar]
- Li, H.J. Effects of Broodstock Enhancement on Fish Larval Resources and Genetic Diversity of the Four Major Chinese Carps in the Middle Reaches of the Yangtze River. Master’s Thesis, Southwest University, Chongqing, China, 2019. [Google Scholar]
- Luo, H.Z. Study of Main Biology Character and Analysis of Resource Status on the Harpadon nehereus. Master’s Thesis, Zhejiang Ocean University, Zhoushan, China, 2012. [Google Scholar]
- Du, X.X. Biological Characteristics and Spatial Distribution Patter of Harpadon nehereus in Offshore Water of Southern Zhejiang. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2018. [Google Scholar]
- Flint, R.W. Long–term estuarine variability and associated biological response. Estuaries 1985, 8, 158–169. [Google Scholar] [CrossRef]
- Harris, S.A.; Cyrus, D.P. Occurrence of fish larvae in the St Lucia Estuary, Kwazulu–Natal, South Africa. S. Afr. J. Mar. Sci. 1995, 16, 333–350. [Google Scholar] [CrossRef]
- Selleslagh, J.; Amara, R. Environmental factors structuring fish composition and assemblages in a small macrotidal estuary (eastern English Channel). Estuar. Coast. Shelf Sci. 2008, 79, 507–517. [Google Scholar] [CrossRef]
- Prista, N.; Vasconcelos, R.P.; Costa, M.J.; Cabral, H. The demersal fish assemblage of the coastal area adjacent to the Tagus estuary (Portugal): Relationships with environmental conditions. Oceanol. Acta 2003, 26, 525–536. [Google Scholar] [CrossRef]
- Zhang, Y.Q. Environmental Impact on the Fish Assemblage Structure in Adjacent Sea Area of the Yangtze River Estuary. Master’s Thesis, Institute of Oceanography, Chinese Academy of Sciences, Qingdao, China, 2012. [Google Scholar]
- Suzumura, M.; Kokubun, H.; Arata, N. Distribution and characteristics of suspended particulate matter in a heavily eutrophic estuary, Tokyo Bay, Japan. Mar. Pollut. Bull. 2004, 49, 496–503. [Google Scholar] [CrossRef]
- Qin, Y.S.; Li, F.; Xu, S.M.; Milliman, J.; Limeburner, R. Suspended matter in the south yellow sea. Oceanol. Limnol. Sin. 1989, 20, 101–112. [Google Scholar]
- Xue, W.J.; Qiao, L.L.; Zhong, Y.; Xue, C.; Chen, S.G.; Li, S.H.; Liu, P.; Gao, F. Multiple timescale variation in concentration of surface suspended sediment in Changjiang river estuary. Oceanol. Limnol. Sin. 2019, 50, 1002–1013. [Google Scholar]
- Zhai, S.K.; Zhang, H.J.; Fan, D.J.; Yang, R.M.; Cao, L.H. Corresponding relationship between suspended matter concentration and turbidity on Changjiang Estuary and adjacent sea area. Acta. Sci. Circumst. 2005, 25, 693–699. [Google Scholar]
- Whitfield, A.K. Ichthyofaunal assemblages in estuaries: A South African case study. Rev. Fish Biol. Fish 1999, 91, 51–86. [Google Scholar]
- Akin, S.; Buhan, E.; Winemiller, K.O.; Yimaz, H. Fish assemblage structure of Koycegiz Lagoon-Estuary, Turkey: Spatial and temporal distribution patterns in relation to environmental variation. Estuar. Coast Shelf Sci. 2005, 64, 671–684. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhang, J.; Wu, Y.; Zhu, Z.Y. Characteristics of Dissolved Oxygen and Its Affecting Factors in the Yangtze Estuary. J. Environ. Sci. 2007, 28, 1649–1654. [Google Scholar]
- Wu, R.S. Hypoxia: From molecular responses to ecosystem responses. Mar. Pollut. Bull. 2002, 45, 35–45. [Google Scholar] [CrossRef]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef]
- Wang, P.H.; Li, B. Historical Changes and Mechanism of Hypoxia in the Changjiang Estuary and Its Adjacent Waters. J. Zhejiang Ocean. Univ. 2019, 38, 401–406. [Google Scholar]
- Hajisamae, S.; Yeesin, P. Patterns in community structure of trawl catches along coastal area of the south China sea. Raffles Bull. Zool. 2010, 58, 357–368. [Google Scholar]
- Keller, A.A.; Simon, V.; Chan, F.; Wakefield, W.W.; Clarke, M.E.; Barth, J.A.; Kamikawa, D.; Fruh, E.L. Demersal fish and invertebrate biomass in relation to an offshore hypoxic zone along the US west coast. Fish. Oceanogr. 2010, 19, 76–87. [Google Scholar] [CrossRef]
- Hassell, K.L.; Coutin, P.C.; Nugegoda, D. Hypoxia impairs embryo development and survival in black bream (Acanthopagrus butcheri). Mar. Pollut. Bull. 2008, 57, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.B.; Cui, G.C.; Hu, C.L.; Zhu, H.C.; Wang, Y.L.; Jiang, R.J.; Zhang, Y.Z.; Feng, C.L. Effects of environmental factors on growth and distribution of young chub mackerel scomber japonicus along the coast of zhejiang in spring. Oceanol. Limnol. Sin. 2021, 52, 1448–1455. [Google Scholar]
- Liu, X.J.; Huang, G.Q.; Li, J.; Tang, X. Response of Oxidative Stress Indicators in Liver and Muscle of Mullet Liza haematocheila to Variation in Dissolyed Oxygen Levels. Fish. Sci. 2014, 33, 344–349. [Google Scholar]
- Wilding, J.L. The oxygen threshold for three species of fish. Ecology 1939, 20, 253–263. [Google Scholar] [CrossRef]
- Sun, Y.; Lv, F.H.; Chen, Z.; Diao, X.Y.; Jiang, J.G.; Wei, C.J.; Pan, J. Spatial–temporal distribution and dynamics of dissolved oxygen in an adjacent area of the Changjiang estuary. Mar. Sci. 2021, 45, 86–96. [Google Scholar]
- Chang, Z.C.; Wen, H.S.; Zhang, M.Z.; Li, J.F.; Li, Y.; Zhang, K.Q.; Wang, W.; Liu, Y.; Tian, Y.; Wang, X.L. Effects of dissolved oxygen levels on oxidative stress response and energy utilization of juvenile Chinese seabass (Lateolabrax maculatus) and associate physiological mechanisms. Period. Ocean Univ. China 2018, 48, 20–28. [Google Scholar]
- Hamilton, B.R.; Peterson, C.T.; Dawdy, A.; Grubbs, R.D. Environmental correlates of elasmobranch and large fish distribution in a river–dominated estuary. Mar. Ecol. Prog. Ser. 2022, 688, 83–98. [Google Scholar] [CrossRef]
- Murillo, F.J.; Serrano, A.; Kenchington, E.; Mora, J. Epibenthic assemblages of the Tail of the Grand Bank and Flemish Cap (northwest Atlantic) in relation to environmental parameters and trawling intensity. Deep Sea Res. Part I 2016, 109, 99–122. [Google Scholar] [CrossRef]
- Beaudreau, A.H.; Bergstrom, C.A.; Whitney, E.J.; Duncan, D.H.; Lundstrom, N.C. Seasonal and interannual variation in high latitude estuarine fish community structure along a glacial to non–glacial watershed gradient in Southeast Alaska. Environ. Biol. Fishes 2022, 105, 431–452. [Google Scholar] [CrossRef]
- Ren, Q.Q.; Xian, W.W.; Liu, C.L.; Li, W.L. Spring–time nektonic invertebrate assemblages of and adjacent to the Yangtze Estuary. Estuar. Coast. Shelf Sci. 2019, 227, 106338. [Google Scholar] [CrossRef]
- Chapman, E.D.; Miller, E.A.; Singer, G.P.; Hearn, A.R.; Thomas, M.J.; Brostoff, W.N.; LaCivita, P.E.; Klimley, A.P. Spatiotemporal occurrence of green sturgeon at dredging and placement sites in the San Francisco estuary. Environ. Biol. Fishes 2019, 102, 27–40. [Google Scholar] [CrossRef]
- Sheaves, M.; Johnston, R. Ecological drivers of spatial variability among fish fauna of 21 tropical Australian estuaries. Mar. Ecol. Prog. Ser. 2009, 385, 245–260. [Google Scholar] [CrossRef]
- Ravelo, A.M.; Konar, B.; Bluhm, B.A. Spatial variability of epibenthic communities on the Alaska Beaufort Shelf. Polar Biol. 2015, 38, 1783–1804. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | 2012 | 2013 | 2014 | 2015 | 2016 |
|---|---|---|---|---|---|
| Depth (D) | 42.6 ± 11.12 | 41.14 ± 11.93 | 38.47 ± 8.41 | 37.67 ± 8.05 | 40.4 ± 9.53 |
| Salinity (S) | 32.3 ± 0.57 bc | 33.51 ± 0.48 a | 30.36 ± 2.32 c | 31.01 ± 3.12 b | 32.56 ± 1.12 b |
| Temperature (T) | 19.58 ± 0.56 c | 21.36 ± 0.34 a | 20.6 ± 0.51 b | 21.32 ± 0.97 a | 21.56 ± 0.86 a |
| Dissolved oxygen (DO) | 7.4 ± 0.41 a | 7.26 ± 0.17 ab | 7.37 ± 1.14 b | 6.41 ± 0.74 b | 7.13 ± 0.55 b |
| pH | 8.46 ± 0.09 a | 8.23 ± 0.05 b | 7.99 ± 0.29 bc | 7.93 ± 0.05 c | 8.36 ± 0.13 a |
| Chemical oxygen demand (COD) | 0.63 ± 0.31 b | 0.93 ± 0.26 a | 1.22 ± 0.82 a | 1.38 ± 0.83 a | 1.12 ± 0.38 a |
| Total nitrogen (TN) | 13.22 ± 4.5 b | 16.52 ± 9.16 b | 28.45 ± 15.05 a | 19.17 ± 11.48 ab | 8.37 ± 10.21 c |
| Total phosphorus (TP) | 0.69 ± 0.2 cd | 0.9 ± 0.45 bc | 0.95 ± 0.21 ab | 1.13 ± 0.31 a | 0.6 ± 0.28 d |
| Total suspended particles (TSP) | 7.28 ± 5.18 a | 13.53 ± 17 a | 2.66 ± 0.85 b | 2.05 ± 1.53 b | 5.21 ± 2.15 a |
| Chlorophyll a (Chl a) | 0.2 ± 0.04 b | 0.52 ± 0.15 a | 0.32 ± 0.5 b | 0.55 ± 0.41 a | 0.37 ± 0.16 a |
| Groups | ANOSIM | SIMPER | |
|---|---|---|---|
| R | P | Average Dissimilarity | |
| 2012 vs. 2013 | 0.073 | 0.057 | 54.32 |
| 2012 vs. 2014 | −0.005 | 0.457 | 54.64 |
| 2012 vs. 2015 | 0.188 | 0.006 | 76.42 |
| 2012 vs. 2016 | 0.095 | 0.04 | 62.34 |
| 2013 vs. 2014 | 0.149 | 0.01 | 51.64 |
| 2013 vs. 2015 | 0.433 | 0.001 | 77.9 |
| 2013 vs. 2016 | 0.374 | 0.001 | 62.67 |
| 2014 vs. 2015 | 0.28 | 0.001 | 73.68 |
| 2014 vs. 2016 | 0.055 | 0.097 | 56.15 |
| 2015 vs. 2016 | 0.164 | 0.005 | 72.66 |
| IRI | |||||
|---|---|---|---|---|---|
| Dominant Species | 2012 | 2013 | 2014 | 2015 | 2016 |
| Collichthys lucidus | 253.12 | 533.08 | 337.46 | 1.08 | 327.92 |
| Larimichthys polyactis | 391.90 | 88.95 | 289.55 | 2940.67 | 603.06 |
| Trichiurus japonicus | 4437.75 | 2871.92 | 3692.76 | 647.09 | 5590.32 |
| Setipinna taty | 1956.62 | 866.09 | 3269.56 | 207.99 | 672.35 |
| Pampus argenteus | 108.01 | 116.63 | 579.06 | 427.41 | 929.86 |
| Harpadon nehereus | 10,420.92 | 14,588.79 | 10,384.48 | 8580.33 | 9031.50 |
| Axis | 1 | 2 | 3 | Total Inertia |
|---|---|---|---|---|
| Eigenvalues | 0.1538 | 0.0182 | 0.0092 | |
| Species–environment correlations | 0.6058 | 0.2833 | 0.3755 | |
| Cumulative percentage variance | ||||
| of species data | 15.38 | 17.2 | 18.12 | |
| of species–environment relation | 84.89 | 94.91 | 100 | |
| Sum of all eigenvalues | 1 | |||
| Sum of all canonical eigenvalues | 0.18 |
| Typical Species | Contribution Percentage for Average Similarity (%) | |
|---|---|---|
| Deep Area | Shallow Area | |
| Trichiurus japonicus | 36.47 | 16.65 |
| Harpadon nehereus | 30.96 | 52.27 |
| Setipinna taty | 9.72 | 17.36 |
| Larimichthys polyactis | 8.63 | 2.91 |
| Apogonichthys lineatus | 5.13 | 0.14 |
| Environmental Factors | Deep Area | Shallow Area | ||
|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | |
| Depth (D) | 47.97 ± 9.15 a | 30–62 | 32.95 ± 3.23 b | 23–41 |
| Salinity (S) | 32.04 ± 2.37 | 24.93–33.82 | 31.85 ± 1.92 | 23.72–33.93 |
| Temperature (T) | 21.16 ± 1.05 | 18.92–22.94 | 20.62 ± 0.90 | 18.60–22.04 |
| Dissolved oxygen (DO) | 7.13 ± 0.56 | 6.24–8.86 | 7.10 ± 0.94 | 3.43–8.57 |
| pH | 8.18 ± 0.28 | 7.46–8.61 | 8.20 ± 0.23 | 7.56–8.58 |
| Chemical oxygen demand (COD) | 1.12 ± 0.70 | 0.26–3.66 | 1.01 ± 0.57 | 0.34–3.40 |
| Total nitrogen (TN) | 17.60 ± 14.02 | 2.16–66.43 | 16.75 ± 11.21 | 3.06–45.55 |
| Total phosphorus (TP) | 0.77 ± 0.34 | 0.27–1.65 | 0.93 ± 0.35 | 0.46–2.04 |
| Total suspended particles (TSP) | 3.22 ± 2.47 a | 0.26–9.90 | 8.59 ± 11.37 b | 0.51–71.56 |
| Chlorophyll a (Chl a) | 0.38 ± 0.36 | 0.01–1.79 | 0.40 ± 0.31 | 0.02–1.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Liang, C.; Xian, W. Spatial and Temporal Variations in Autumn Fish Assemblages in the Offshore Waters of the Yangtze Estuary. Diversity 2023, 15, 669. https://doi.org/10.3390/d15050669
Chen Z, Liang C, Xian W. Spatial and Temporal Variations in Autumn Fish Assemblages in the Offshore Waters of the Yangtze Estuary. Diversity. 2023; 15(5):669. https://doi.org/10.3390/d15050669
Chicago/Turabian StyleChen, Zhaomin, Cui Liang, and Weiwei Xian. 2023. "Spatial and Temporal Variations in Autumn Fish Assemblages in the Offshore Waters of the Yangtze Estuary" Diversity 15, no. 5: 669. https://doi.org/10.3390/d15050669
APA StyleChen, Z., Liang, C., & Xian, W. (2023). Spatial and Temporal Variations in Autumn Fish Assemblages in the Offshore Waters of the Yangtze Estuary. Diversity, 15(5), 669. https://doi.org/10.3390/d15050669