

Structure, Growth and Histology of Gnathal Elements in Dunkleosteus (Arthrodira, Placodermi), with a Description of a New Species from the Famennian (Upper Devonian) of the Tver Region (North-Western Russia)

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
Institutional Abbreviations
3. Geographical and Geological Setting
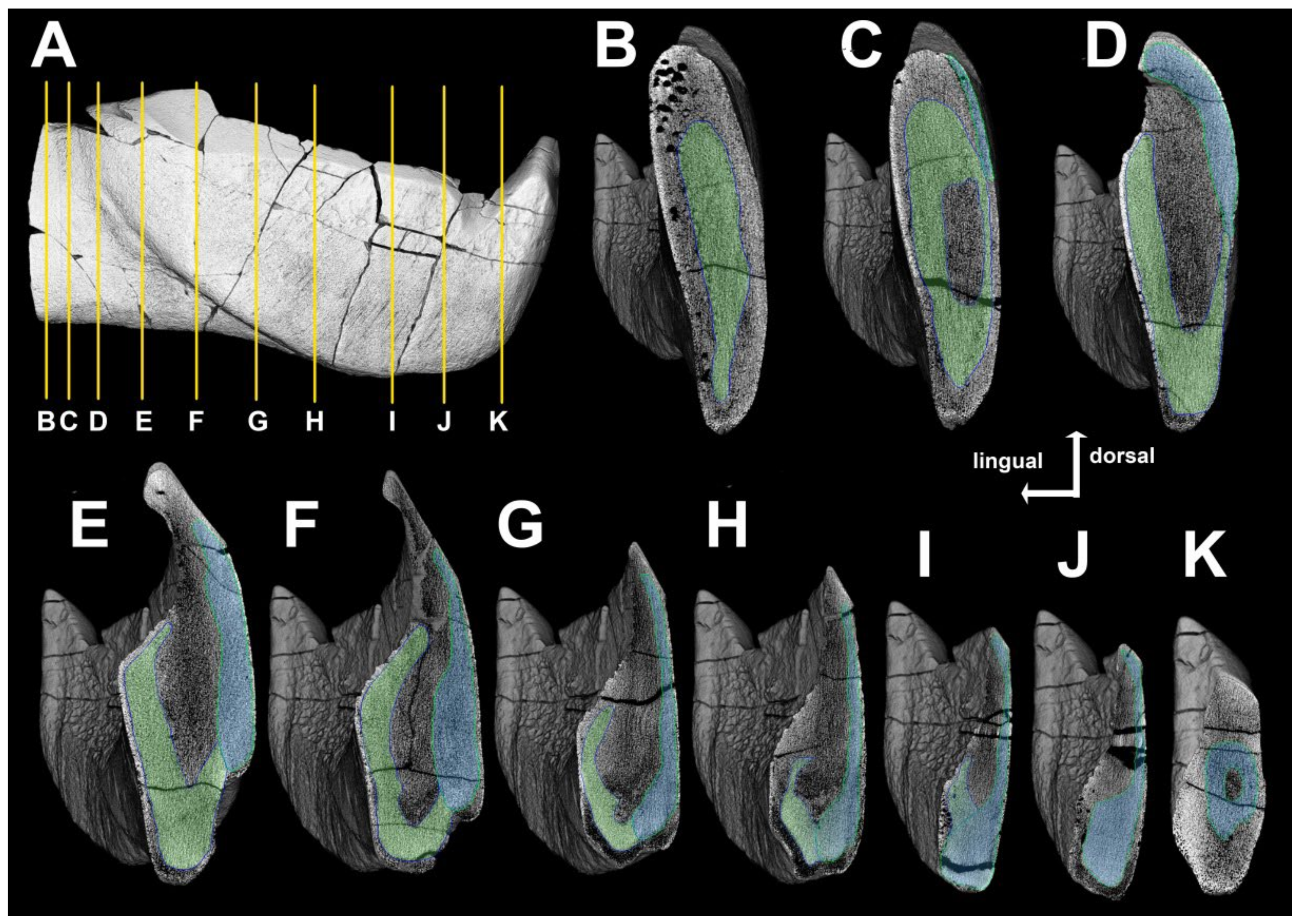
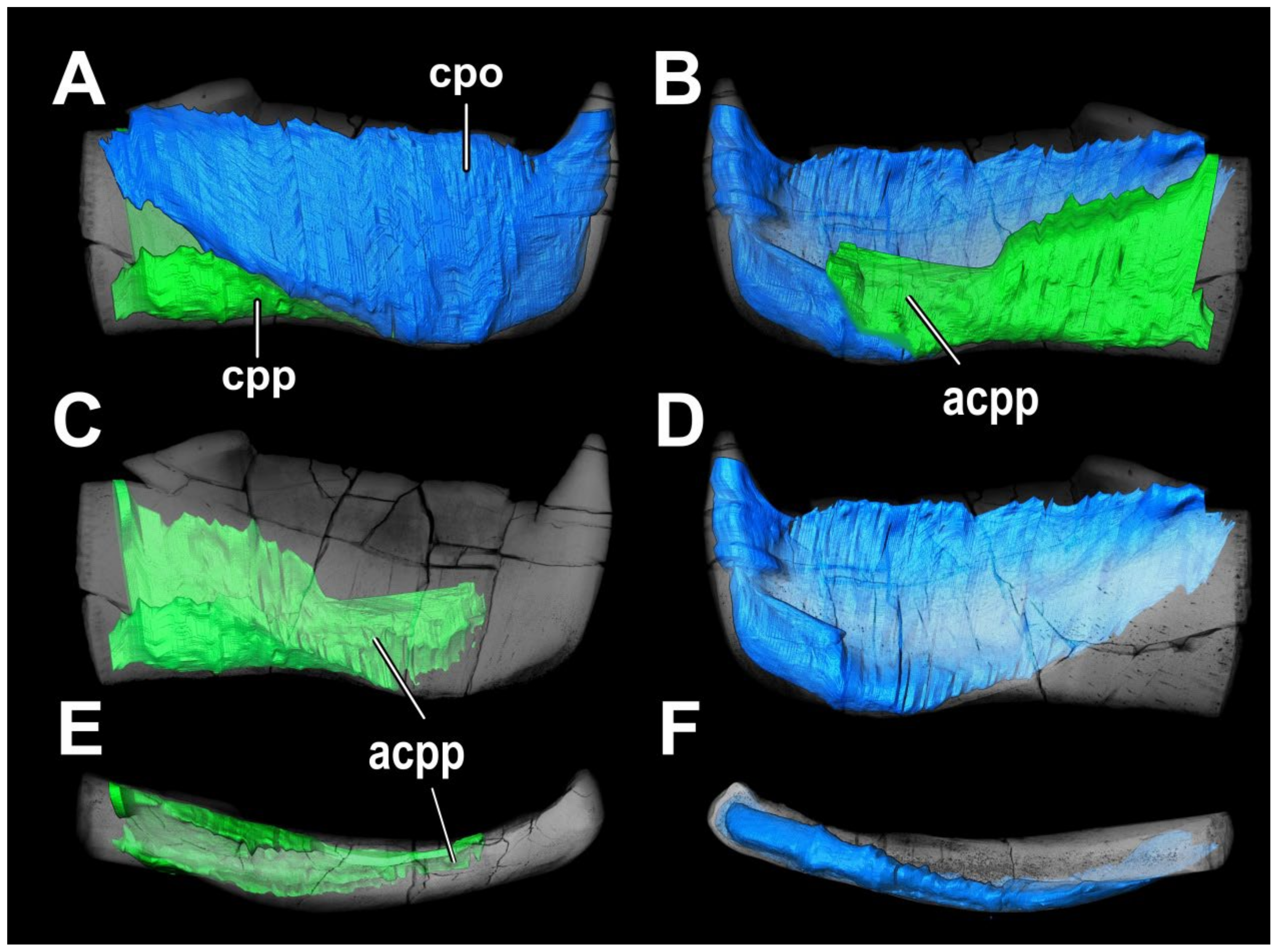
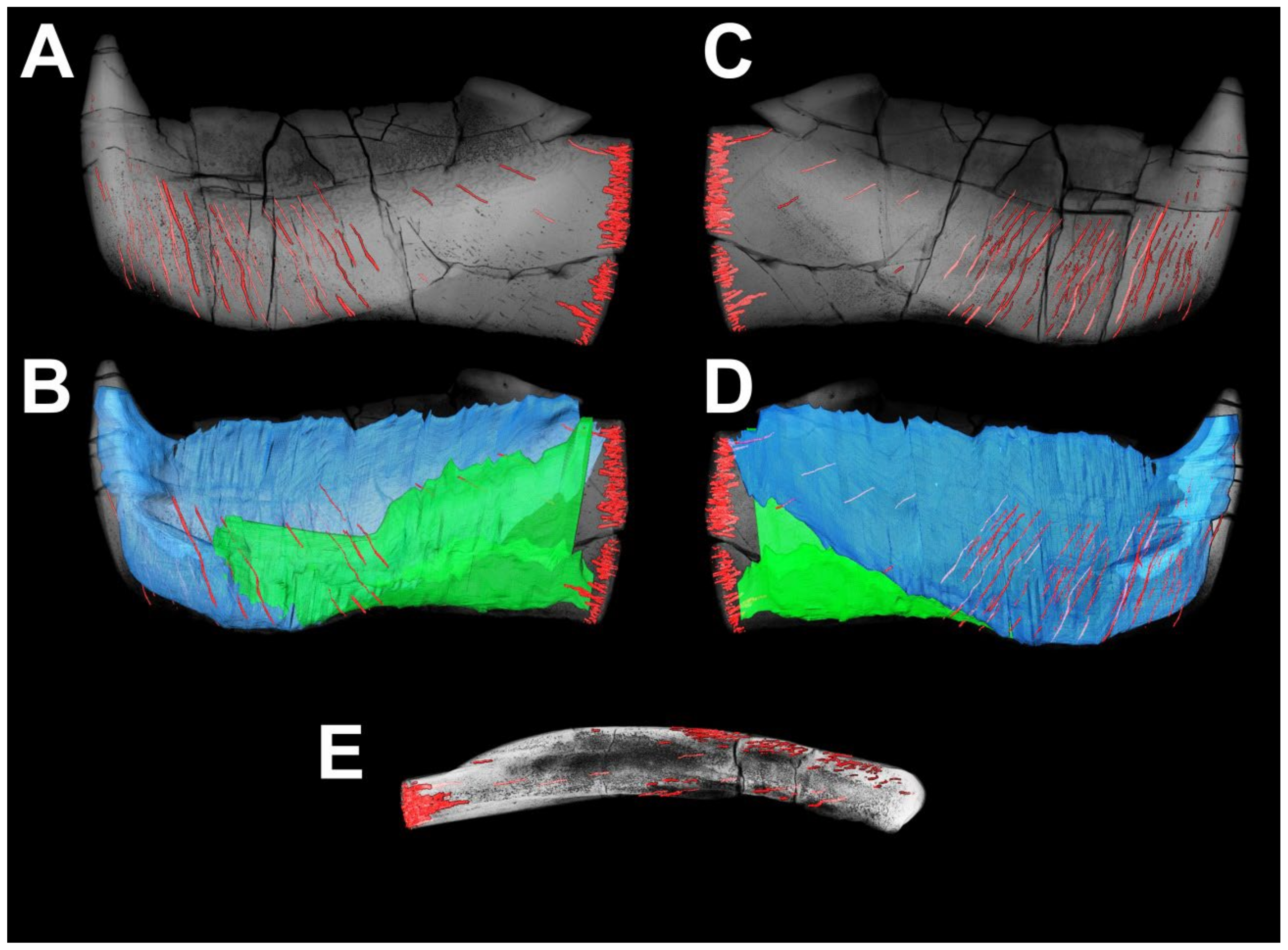
4. Results
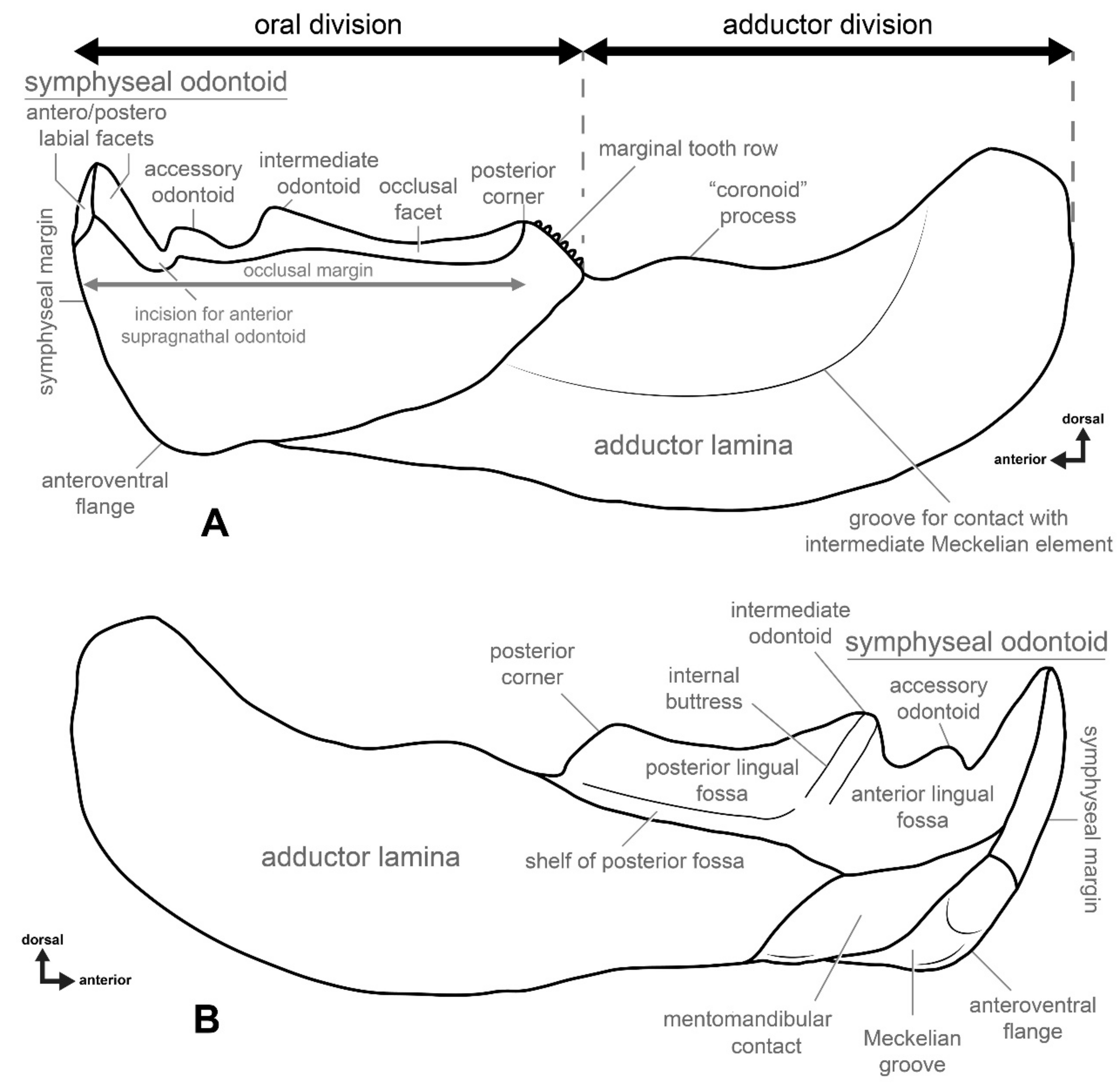
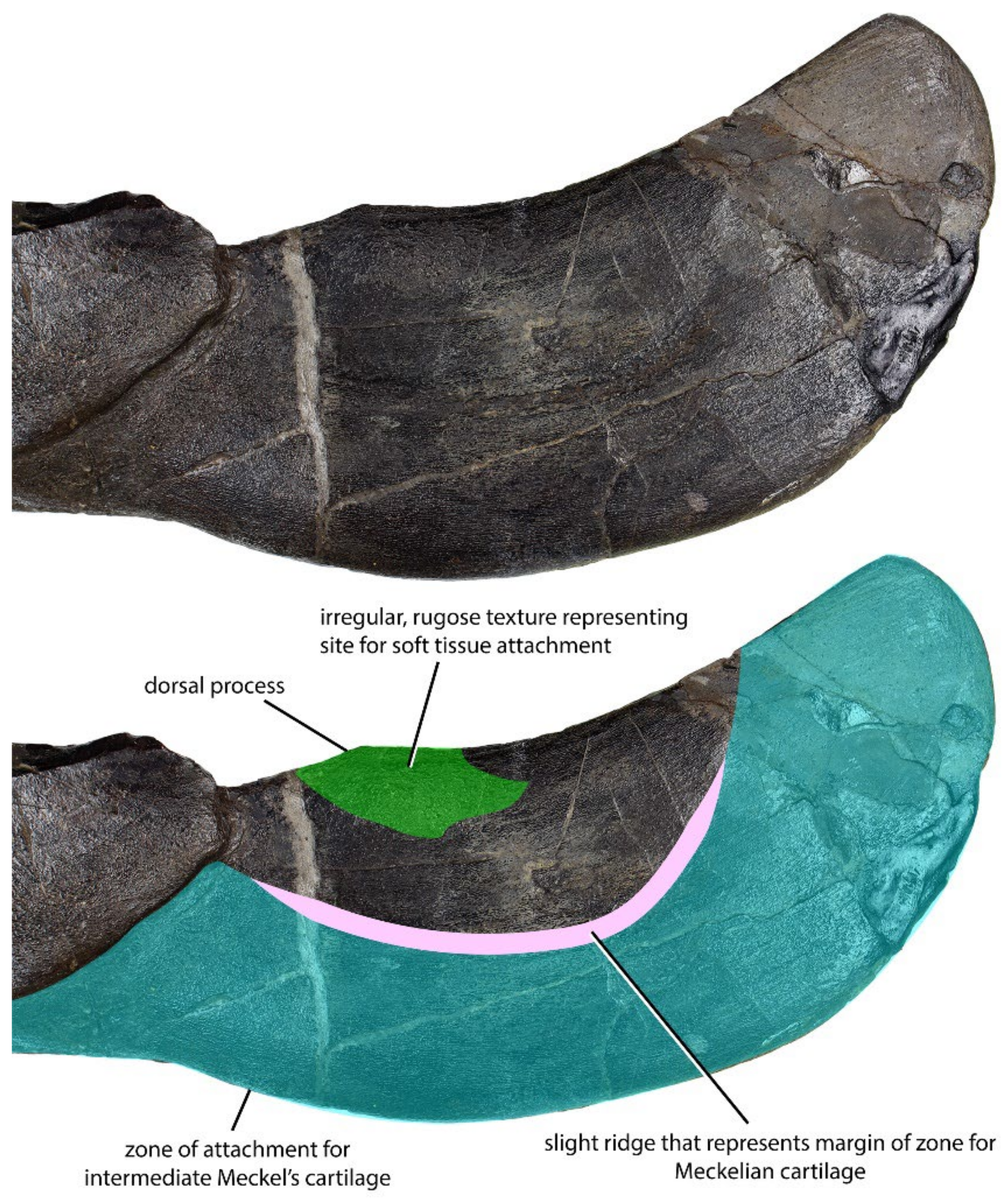
4.1. General Morphology and Terminology
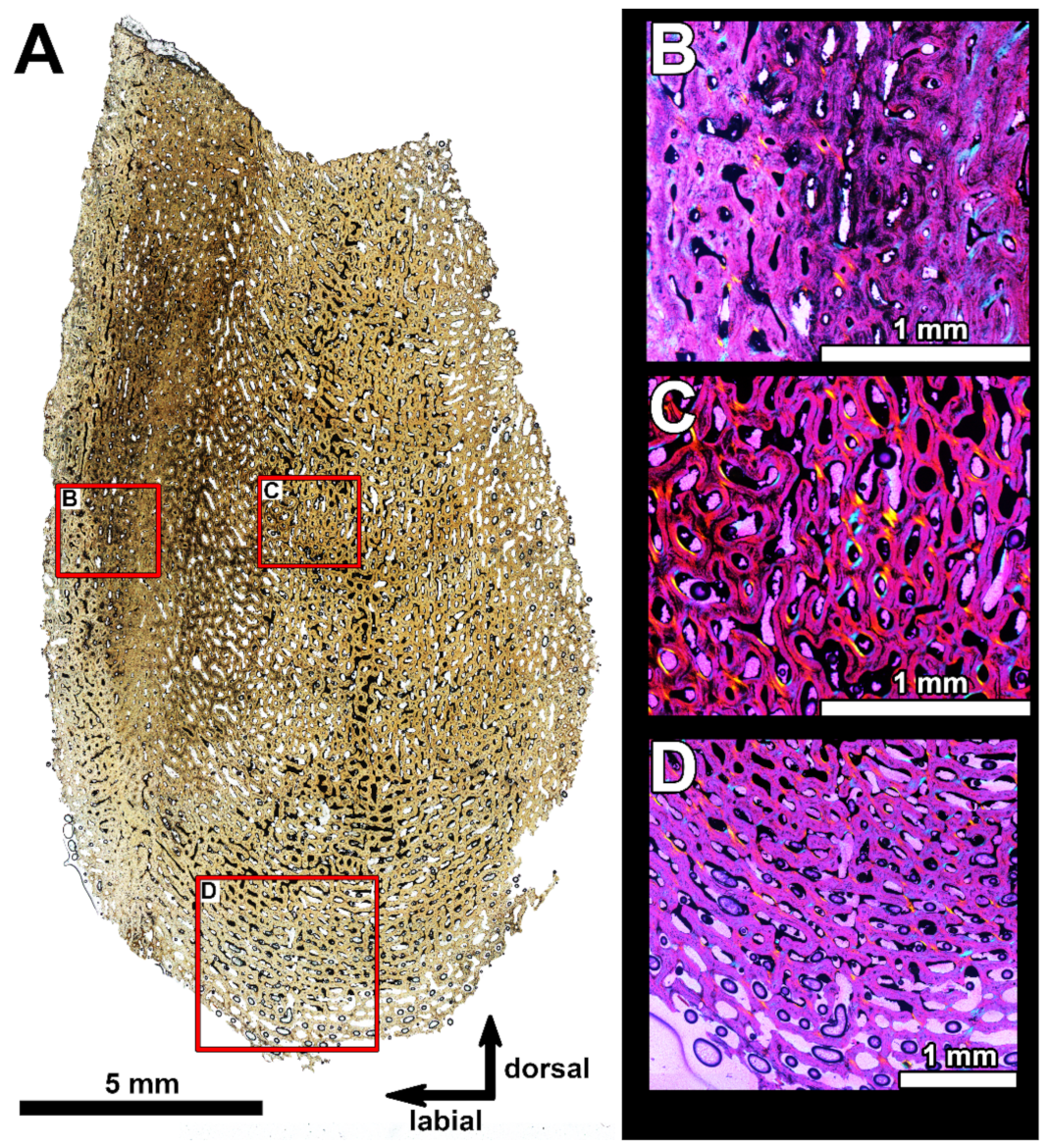
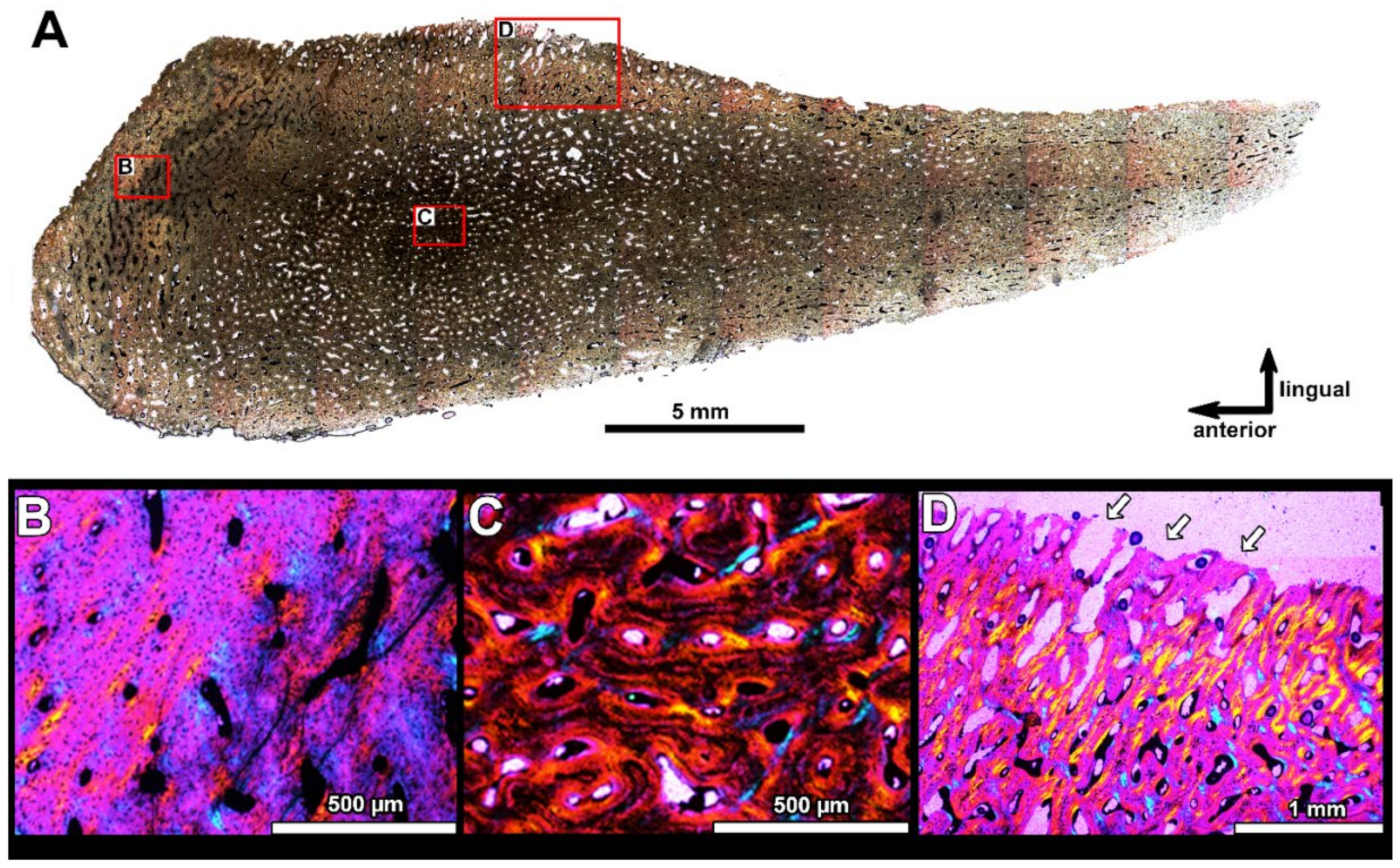
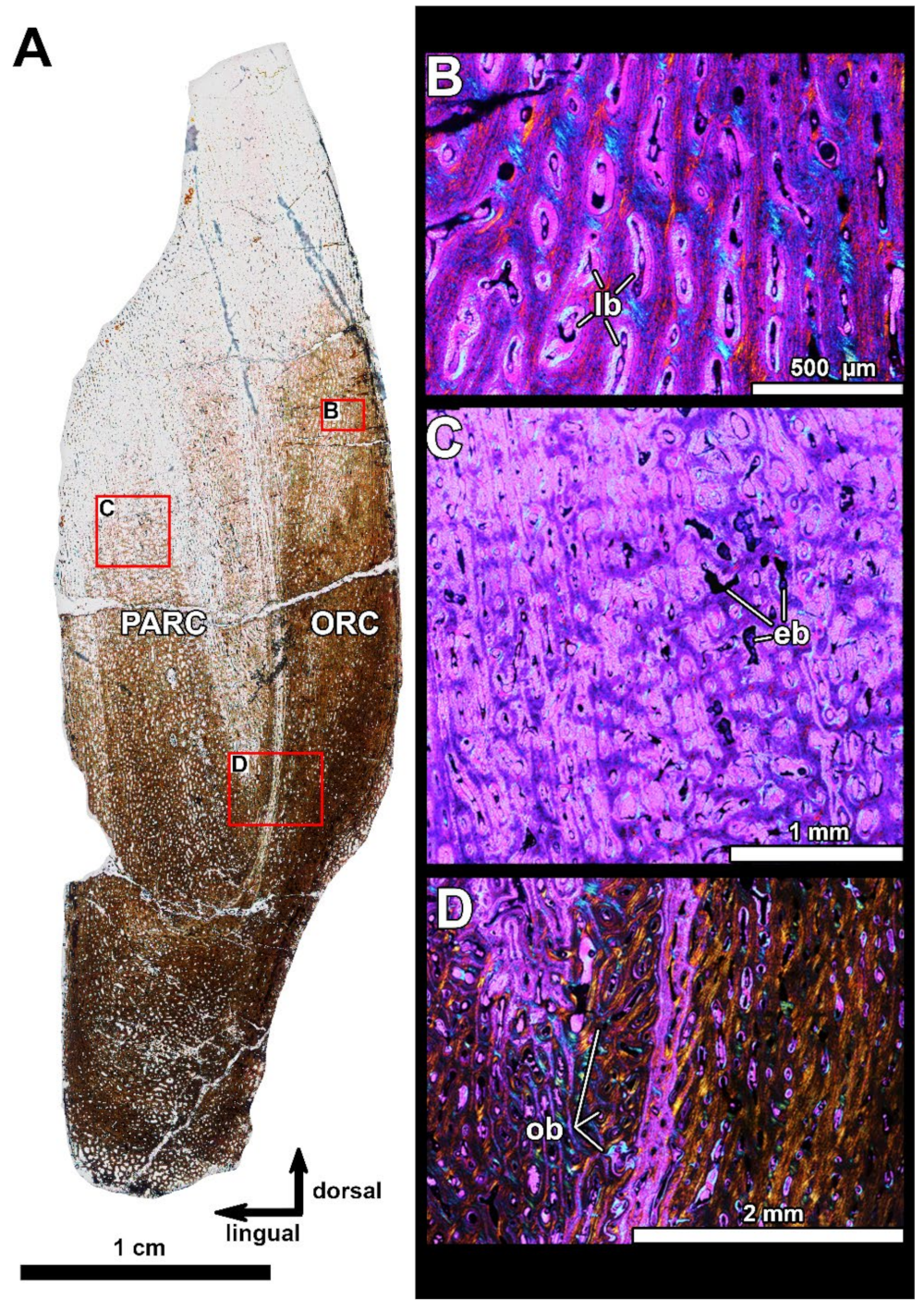
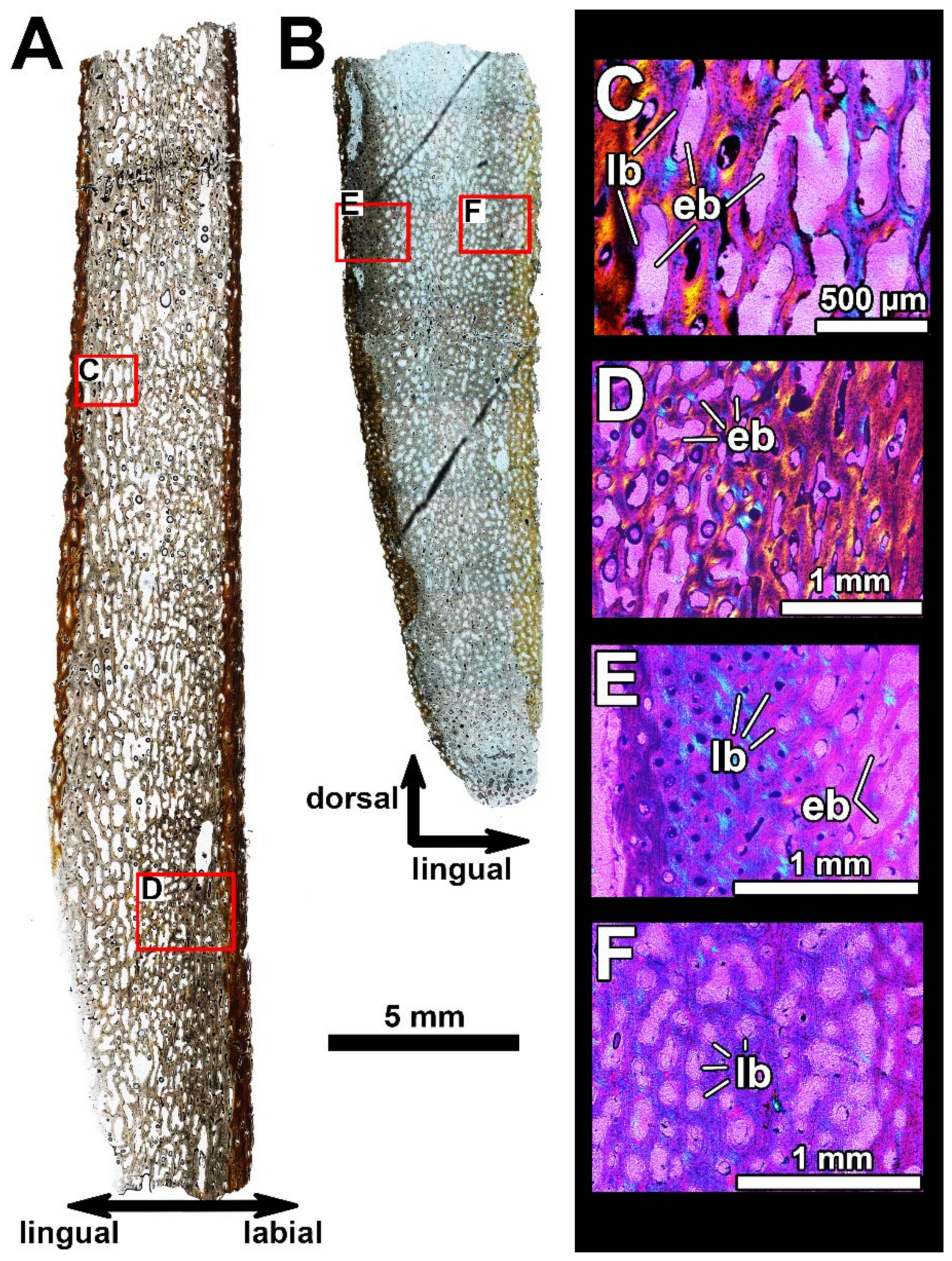
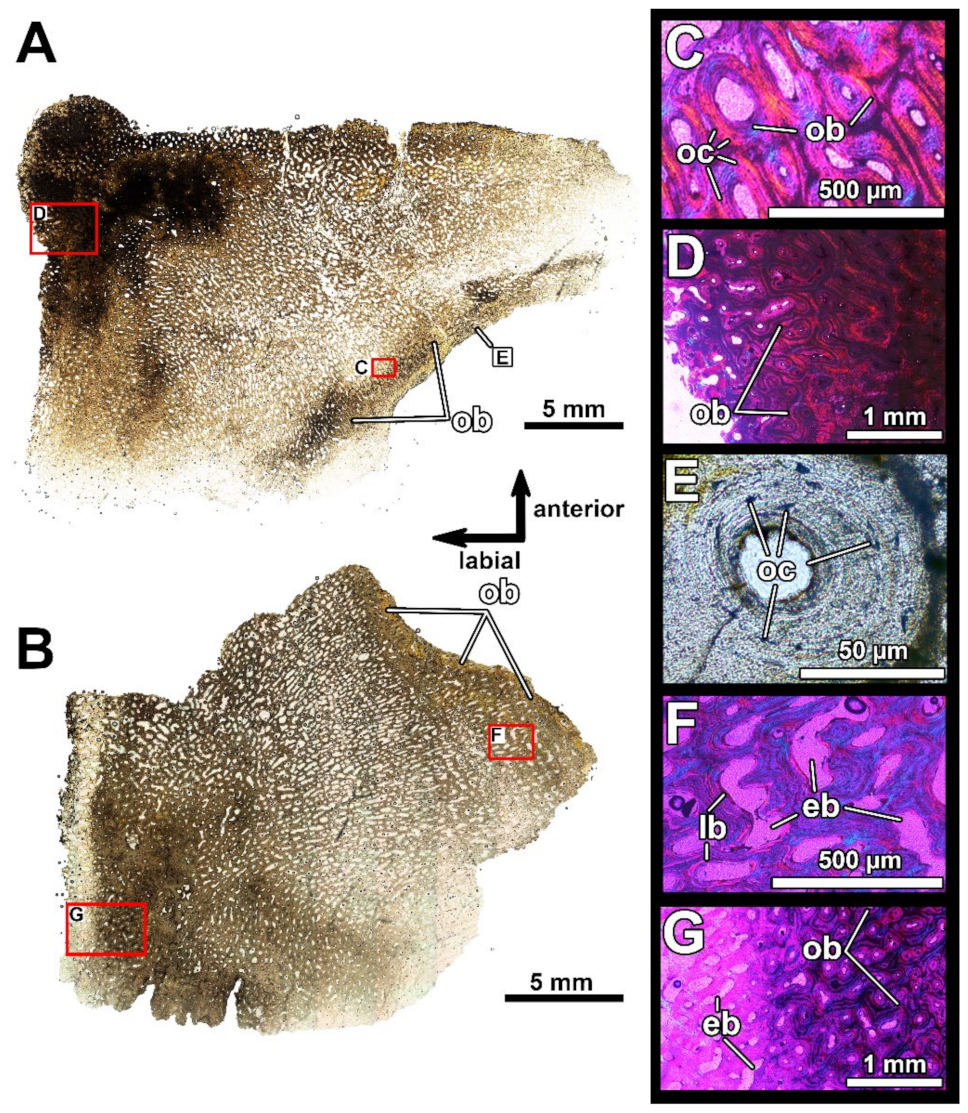
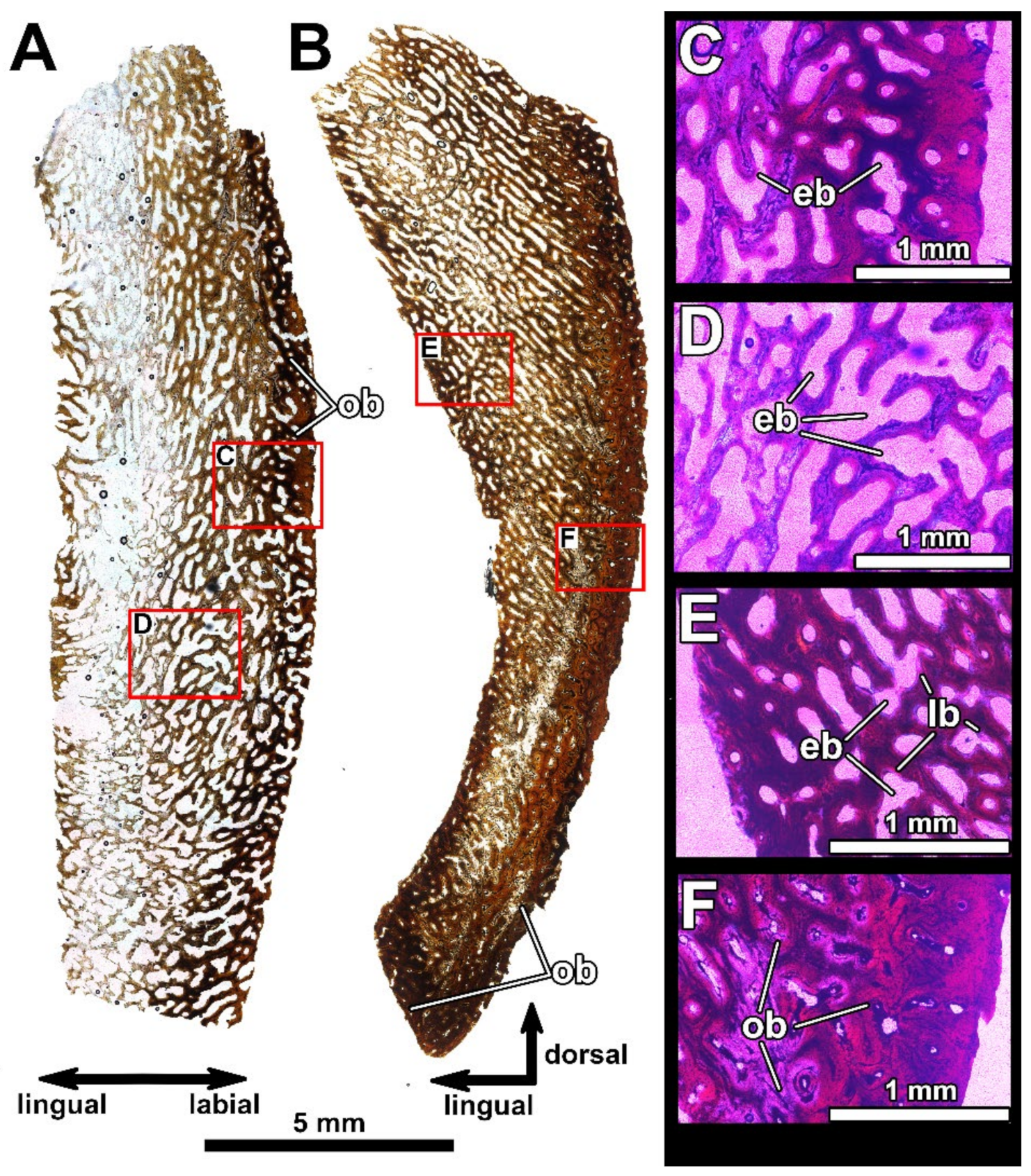
4.2. Histological Studies
4.2.1. Previous Histological Studies of Gnathal Elements in the Dunkleosteidae
4.2.2. Histology of the Gnathal Elements in Dunkleosteidae Indet
4.3. Systematic Status of KMA 4155
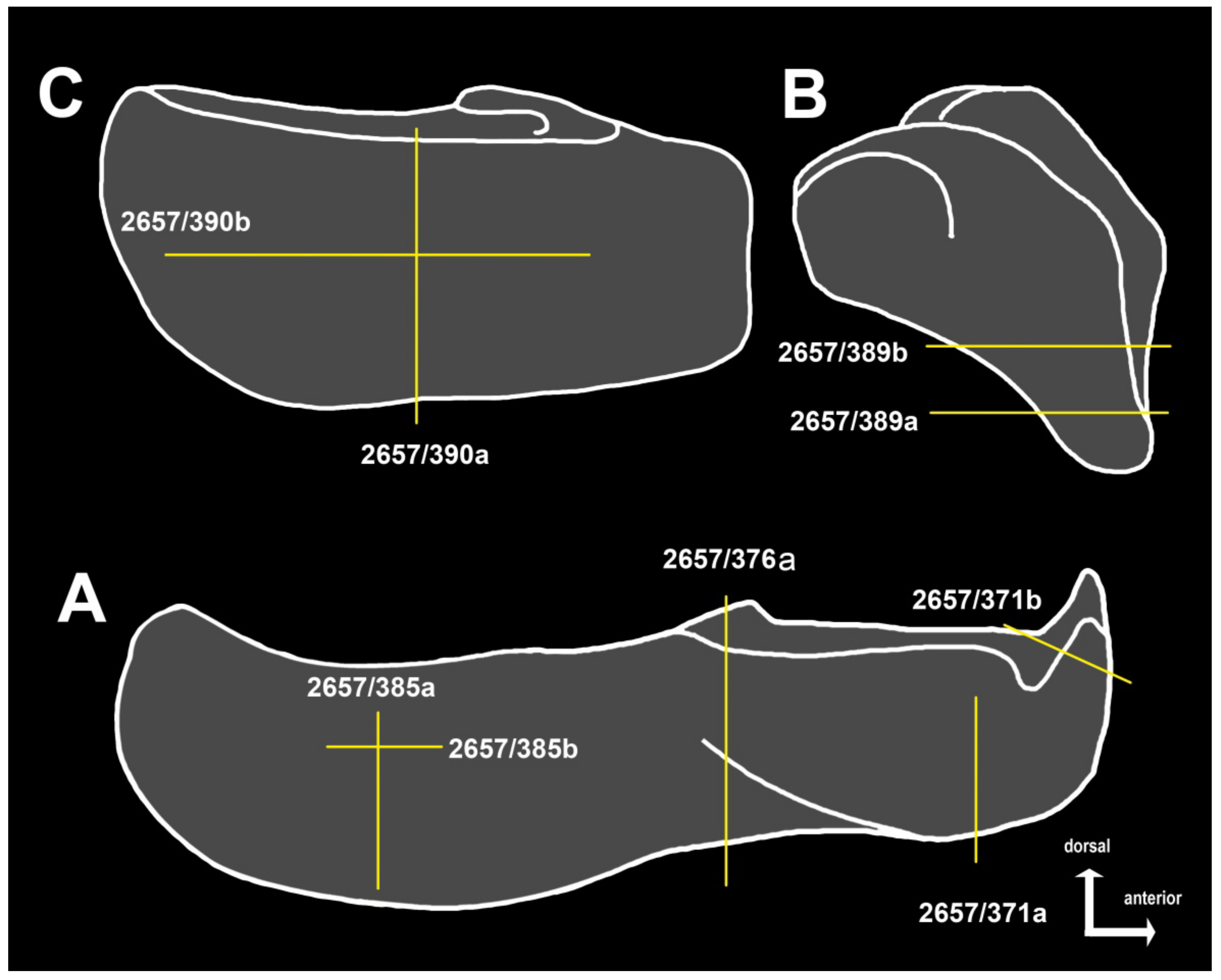
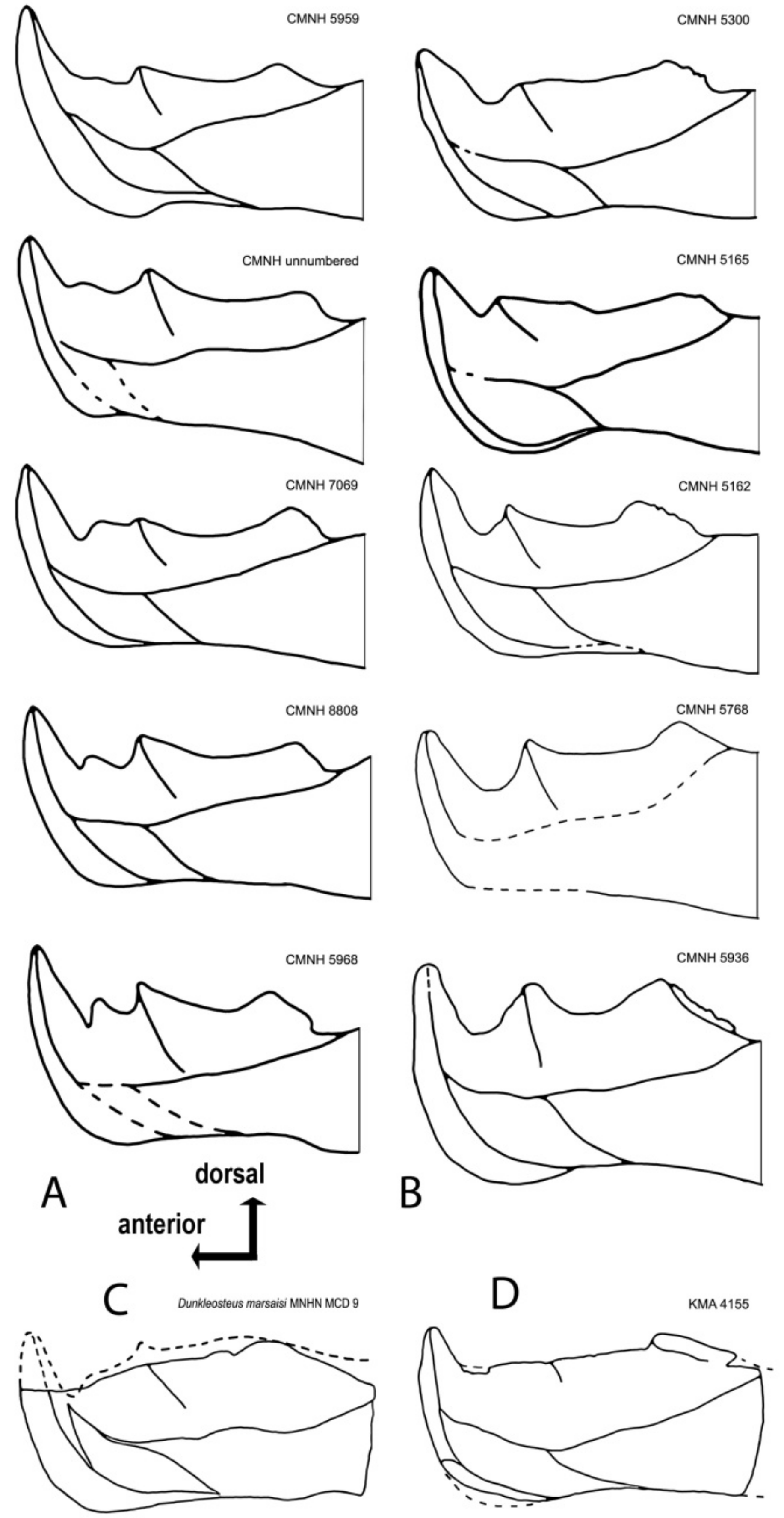
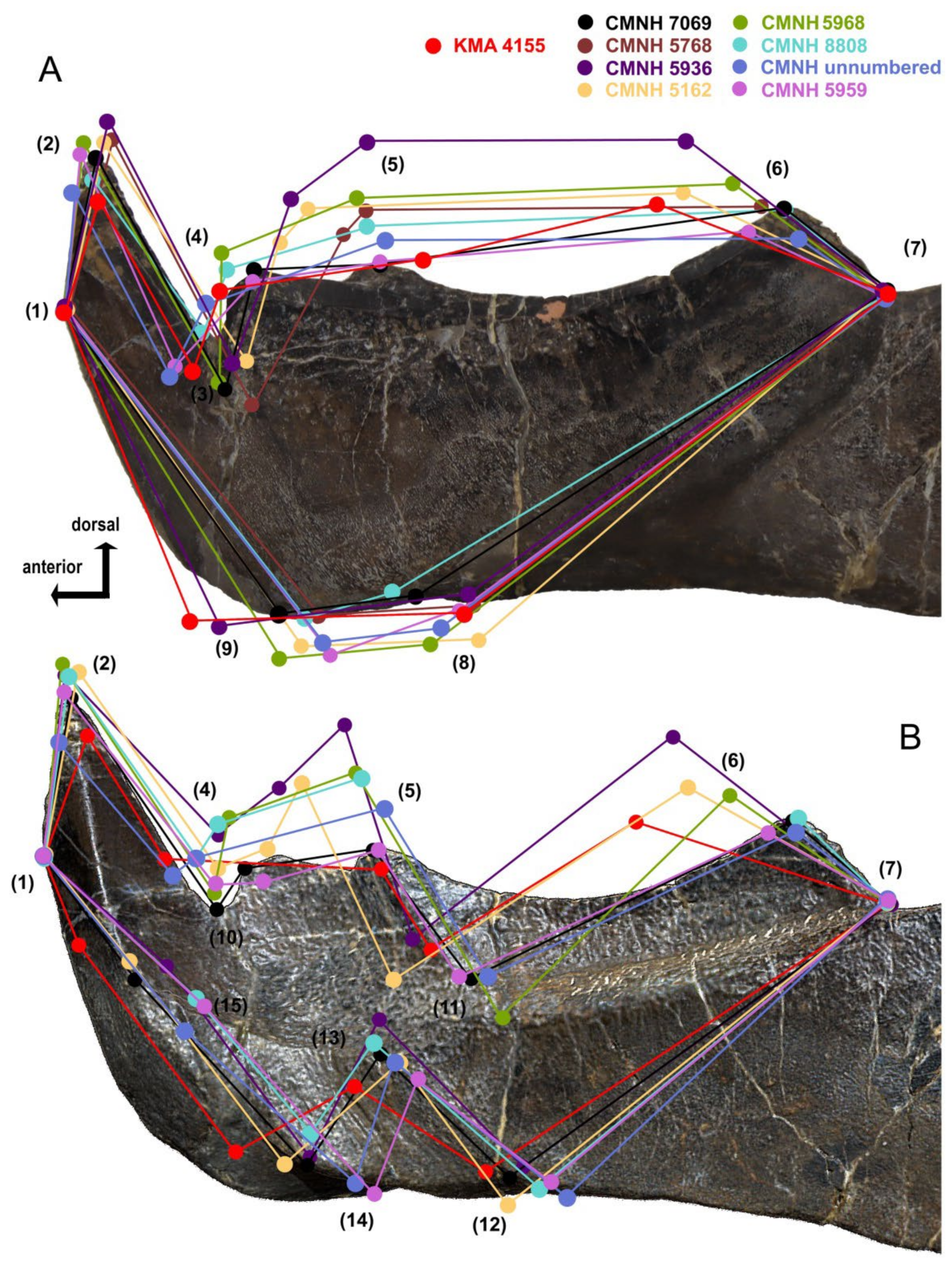
4.4. Intraspecific Variation in the Oral Division of Dunkleosteus terrelli
5. Systematic Paleontology
| Placodermi McCoy, 1848 |
| Arthrodira Woodward, 1891 |
| Pachyosteomorphi Stensiö, 1944 |
| Superfamily Dunkleosteoidea Vézina, 1990 |
| Family Dunkleosteidae Stensiö, 1963 |
| Genus Dunkleosteus Lehman, 1956 |
| Dunkleosteus tuderensis Lebedev, sp. nov. |
- Etymology. The specific epithet tuderensis refers to the locality of the discovery of the holotype specimen on the bank of the Maliy (Minor) Tuder River in the Tver Region of Russia.
- Holotype. KMA 4155, oral division of a left infragnathal.
- Provenance. Left bank of Maliy Tuder River about 200 m to the north–northwest of the currently abandoned Bilovo village, Toropets District, Tver Region, north-western Russia; Bilovo Formation, (?) Lower-Middle Famennian, Upper Devonian.
- Diagnosis. Symphyseal odontoid short, its height three times smaller than greatest depth of the oral division from the base of the odontoid. Axis of symphyseal odontoid perpendicular to the occlusal margin. Occlusal margin almost straight, with posterior corner gently elevated. Intermediate odontoid poorly developed (vestigial), its buttress hardly discernible. Lingual shelf of the lingual fossa low, the anterior and posterior fossae grade smoothly into one another.
- Remarks. Characters related to the structure of the occlusal margin are important because, despite their variability in Dunkleosteus terrelli, they reflect the structure of the anterior and posterior supragnathal elements and interaction between the upper and lower jaw elements, reflected in a number of characters.
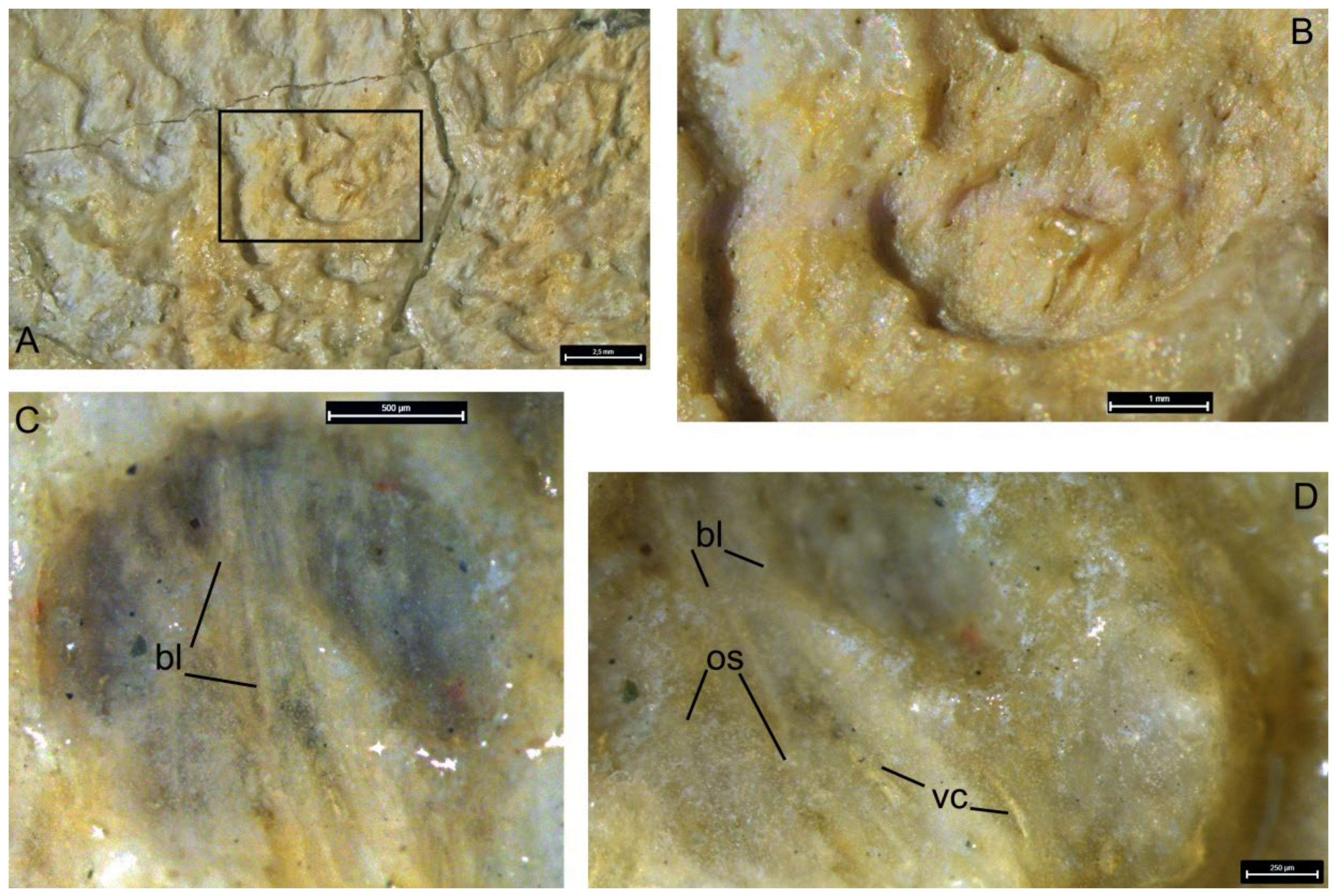
- Description (Figure 14 and Figure 15; Supplementary Figure S2, in greyscale, to present the specimen in higher resolution and contrast).
5.1. General Morphology
5.2. Superficial Microstructure
5.3. Internal Structure
5.4. Vascular System
6. Discussion
6.1. Growth of Arthrodire Gnathal Elements
6.2. Resorption Field
6.3. Microanatomy and Histology of the Gnathal Bones
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engelman, R.K. A Devonian fish tale: A new method of body length estimation suggests much smaller sizes for Dunkleosteus terrelli (Placodermi: Arthrodira). Diversity 2023, 15, 318. [Google Scholar] [CrossRef]
- Zhu, M.; Ahlberg Per, E.; Pan, Z.; Zhu, Y.; Qiao, T.; Zhao, W.; Jia, L.; Lu, J. A Silurian maxillate placoderm illuminates jaw evolution. Science 2016, 354, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Yu, X.; Ahlberg, P.E.; Choo, B.; Lu, J.; Qiao, T.; Qu, Q.; Zhao, W.; Jia, L.; Blom, H.; et al. A Silurian placoderm with osteichthyan-like marginal jaw bones. Nature 2013, 502, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-A.; Ahlberg Per, E.; Zhu, M. The evolution of dermal jaw bones in the light of maxillate placoderms. In Evolution and Development of Fishes; Underwood, C., Richter, M., Johanson, Z., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 71–86. [Google Scholar]
- Carr, R.K.; Hlavin, W.J. Two new species of Dunkleosteus lehman, 1956, from the Ohio Shale Formation (USA, Famennian) and the Kettle Point Formation (Canada, Upper Devonian), and a cladistic analysis of the Eubrachythoraci (Placodermi, Arthrodira). Zool. J. Linn. Soc. 2010, 159, 195–222. [Google Scholar] [CrossRef]
- Newberry, J.S. Paleozoic Fishes of North America. Monogr. U.S. Geol. Surv. 1889, 16, 1–228. [Google Scholar]
- Newberry, J.S. Descriptions of fossil fishes. In Report of the Geological Survey of Ohio. Volume II. Geology and Paleontology; Part II; Nevins and Myers, State Printers: Columbus, OH, USA, 1875; Volume 2, pp. 1–64. [Google Scholar]
- Newberry, J.S. Fossil fishes. In Report of the Geological Survey of Ohio. Volume II. Geology and Paleontology; Part II; Nevins and Myers, State Printers: Columbus, OH, USA, 1873; Volume 1, pp. 245–355. [Google Scholar]
- Eastman, C.R. Devonic fishes of the New York formations. NY Mus. State Mem. 1907, 10, 1–235. [Google Scholar]
- Heintz, A. The structure of Dinichthys: A contribution to our knowledge of the Arthrodira. In The Bashford Dean Memorial Volume; Gudger, E.W., Ed.; American Museum of Natural History: New York, NY, USA, 1932; pp. 115–224. [Google Scholar]
- Miles, R.S. Features of Placoderm Diversification and the Evolution of the Arthrodire Feeding Mechanism. Trans. R. Soc. Edinb. 1969, 68, 123–170. [Google Scholar] [CrossRef]
- Dennis-Bryan, K. A new species of eastmanosteid arthrodire (Pisces: Placodermi) from Gogo, Western Australia. Zool. J. Linn. Soc. 1987, 90, 1–64. [Google Scholar] [CrossRef]
- Anderson, P.S.L. Shape variation between arthrodire morphotypes indicates possible feeding niches. J. Vertebr. Paleontol. 2008, 28, 961–969. [Google Scholar] [CrossRef]
- Anderson, P.S.L.; Westneat, M.W. Feeding mechanics and bite force modelling of the skull of Dunkleosteus terrelli, an ancient apex predator. Biol. Lett. 2007, 3, 77–80. [Google Scholar] [CrossRef]
- Anderson, P.S.L.; Westneat, M.W. A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi). Paleobiology 2009, 35, 251–269. [Google Scholar] [CrossRef]
- Snively, E.; Anderson, P.S.L.; Ryan, M.J. Functional and ontogenetic implications of bite stress in arthrodire placoderms. Kirtlandia 2010, 57, 53–60. [Google Scholar]
- Anderson, P.S.L. Using linkage models to explore skull kinematic diversity and functional convergence in arthrodire placoderms. J. Morphol. 2010, 271, 990–1005. [Google Scholar] [CrossRef] [PubMed]
- Claypole, E.W. Structure of the bone of Dinichthys. Proc. Am. Microsc. Soc. 1894, 15, 189–191. [Google Scholar] [CrossRef]
- Stetson, H. Notes on the structure of Dinichthys and Macropetalichthys. Bull. Mus. Comp. Zool. 1930, 71, 19–39. [Google Scholar]
- Johanson, Z.; Smith, M.M. Origin and evolution of gnathostome dentitions: A question of teeth and pharyngeal denticles in placoderms. Biol. Rev. 2005, 80, 303–345. [Google Scholar] [CrossRef]
- Obrucheva, O.P. Remains of Dinichthys (Arthrodira) from the Upper Devonian of USSR. Dokl. Akad. Nauk. SSSR 1956, 108, 333–336. (In Russian) [Google Scholar]
- Denison, R.H. Placodermi. In Handbook of Palaeoichthyology; Schultze, H.-P., Ed.; Gustav Fischer Verlag: Stuttgart, Germany, 1978; Volume 2, pp. 1–128. [Google Scholar]
- Obrucheva, O.P. Pantsyrniye Ryby Devona SSSR. Kokkosteidy, Dinichthidy [Placoderms from the Devonian of USSR (Coccosteids and Dinichthyids)]; Moscow State University Publishers: Moscow, Russia, 1962; pp. 3–190. (In Russian) [Google Scholar]
- Moloshnikov, S.V. Finds of the dunkleosteid placoderms (Pisces, Placodermi) in the European part of Russia (Central Devonian field). Zhizn’ Zemli 2021, 43, 67–76, (In Russian with English Abstract). [Google Scholar] [CrossRef]
- Schultze, H.-P. Large upper Devonian arthrodires from Iran. Fieldiana Geol. 1973, 23, 53–78. [Google Scholar]
- Lebedev, O.A.; Zakharenko, G.V.; Silantiev, V.V.; Evdokimova, I.O. New finds of fishes in the lower uppermost Famennian (Upper Devonian) of Central Russia and habitats of the Khovanshchinian vertebrate assemblages. Est. J. Earth Sci. 2018, 67, 59–75. [Google Scholar] [CrossRef]
- Esin, D.; Ginter, M.; Ivanov, A.; Lebedev, O.; Luksevics, E.; Avkhimovich, V.; Golubtsov, V.; Petukhova, L. Vertebrate correlation of the Upper Devonian and Lower Carboniferous on the East European Platform. Cour. Forsch. Inst. Senckenberg 2000, 233, 341–359. [Google Scholar]
- Obrucheva, O.P.; Obrucheva, E.D. Fishes of the Central Devonian field. In Ocherki po Filogenii i Sistematike Iskopaemyh Ryb [Essays on the Phylogeny and Systematics of Fossil Fishes]; Novitskaya, L.I., Ed.; Nauka: Moscow, Russia, 1977; pp. 24–28. (In Russian) [Google Scholar]
- Moloshnikov, S.V. A unique vertebrate locality in the Upper Devonian deposits (Orel Region). Izv. Vyss. Uchebnykh Zavedeniy. Geol. I Razved. 2001, 3, 29–33. (In Russian) [Google Scholar]
- Hecker, R.F.; Filippova, M.F., VII. A section by the Lovat’ river and its tributaries, bolshoy i maliy tuder, Kunya and Smota rivers. In Deposits of the Main Devonian Field; Hecker, R.F., Obruchev, D.V., Filippova, M.F., Eds.; Trudy Leningradskogo Geologo-Gidro-Geodezicheskogo Tresta; Transactions of the Leningrad Geological: Leningrad, Russia, 1935; Volume 9, pp. 40–82. [Google Scholar]
- Verbitsky, V.R.; Verbitsky, I.V.; Vasilyeva, O.V.; Savanin, V.V. Gosud0arstvennaia Geologicheskaia Karta Rossiiskoi Federatsii. Masshtab 1: 1000000 (tretye pokolenie). Seriya Tsentral’noEvropeiskaia. Listy O-35—Pskov, (N-35), O-36—SanktPetersburg. Ob’iasnitel’naia zapiska [National Geological Map of the Russian Federation. Scale 1: 1000000. Central European Series. Sheets O-35 Pskov, (N-35), O-36 St. Petersburg. Descriptive Notes]; Cartographic Factory VSEGEI: St. Petersburg, Russia, 2012; pp. 3–508. (In Russian) [Google Scholar]
- Davydov, A.E.; Linkevich, V.V. Brachiopods of the genus Cyrtospirifer from the Bilovo Formation (Famennian, Upper Devonian) in the western part of the Tver Region. Bull. Mosk. Obs. Ispyt. Prirody. Otd. Geol. 2019, 94, 21–32. [Google Scholar]
- Shchedukhin, A.Y. Upper Devonian nautiloids from the Bilovo section (Tver Region). In Proceedings of the Materials of the IX International Scientific Conference of Young Scientists “The Young to Earth Sciences”, Moscow, Russia, 16–20 October 2020; pp. 164–166. (In Russian). [Google Scholar]
- Moloshnikov, S.V.; Linkevich, V.V. On the bothriolepidid (Placodermi, Antiarchi) variability: An unusual specimen of Livnolepis heckeri (Lukševičs) from the Famennian of the Tver Region. Izv. Vyss. Uchebnykh Zavedeniy. Geol. I Razved. 2019, 4, 5–10. [Google Scholar] [CrossRef]
- Moloshnikov, S.V.; Linkevich, V.V. Late Devonian Bothriolepidids (Placodermi, Antiarchi) of the Tver Oblast. Paleontol. J. 2020, 54, 157–165. [Google Scholar] [CrossRef]
- Moloshnikov, S.V.; Linkevich, V.V. Systematic position of the Famennian antiarchs (Vertebrata, Placodermi) of the Tver Region. Izv. Vyss. Uchebnykh Zavedeniy. Geol. I Razved. 2017, 2, 8–14. (In Russian) [Google Scholar] [CrossRef]
- Moloshnikov, S.V.; Linkevich, V.V. Armoured fishes of the family Bothriolepididae Cope (Antiarchi) from the Bilovo locality of Famennian fossils (eastern part of the Main Devonian Field). In Proceedings of the Problems of Regional Geology of Northern Eurasia. Summary of reports at the XII Scientific Session dedicated to Prof. M.V. Muratov, Moscow, Russia, 16–17 April, 2020; pp. 72–75. [Google Scholar]
- Lebedev, O.A.; Krupina, N.I.; Linkevich, V.V. The First Record of a Lungfish (Dipnoi, Sarcopterygii) in the Famennian Deposits (Upper Devonian) of the Tver Oblast. Paleontol. J. 2019, 53, 636–646. [Google Scholar] [CrossRef]
- Lebedev, O.A.; Ivanov, A.O.; Linkevich, V.V. Chondrichthyan spines from the Famennian (Upper Devonian) of Russia. Acta Geol. Pol. 2020, 70, 339–362. [Google Scholar] [CrossRef]
- Vasiļkova, J.; Lukševičs, E.; Stinkulis, G.; Zupiņš, I. Taphonomy of the vertebrate bone beds from the Klūnas fossil site, Upper Devonian Tērvete Formation of Latvia. Est. J. Earth Sci. 2012, 61, 105–119. [Google Scholar] [CrossRef]
- Francillon-Vieillot, H.; De Buffrénil, V.; Castanet, J.; Géraudie, J.; Meunier, F.J.; Sire, J.Y.; Zylberberg, L.; De Ricqlés, A. Microstructure and mineralization of vertebrate skeletal tissues. In Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends; Carter, J.G., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1990; Volume 1, pp. 471–530. [Google Scholar]
- Sobolev, N.N.; Evdokimova, I.O. Devonian System. In State of Knowledge of Precambrian and Phanerozoic Stratigraphy in Russia. Aims for Further Studies; Zhamoida, A.I., Petrov, O.V., Eds.; Decisions of the Interdepartmental Stratigraphic Committee and its Permanent Commissions 2008; Russian Geological Research Institute: St. Petersburg, Rusia, 2008; Volume 38, pp. 52–60. (In Russian) [Google Scholar]
- Hecker, R.F. Deposits, fauna and flora of the main devonian field. In Fauna of the Main Devonian Field; Hecker, R.F.A.B.A., Ed.; Academy of Sciences of USSR Press: Moscow-Leningrad, Rusia, 1941; Volume 1, pp. 17–84. [Google Scholar]
- Sammet, E.Y. The eastern part of the main devonian field. In Stratigrafiya SSSR. Devonskaya Sistema [Stratigraphy of the USSR. The Devonian System]; Nalivkin, D.V., Ed.; Nedra: Moscow, Russia, 1973; Volume 1, pp. 90–100. (In Russian) [Google Scholar]
- Carr, R.K.; Johanson, Z.; Ritchie, A. The phyllolepid placoderm Cowralepis mclachlani: Insights into the evolution of feeding mechanisms in jawed vertebrates. J. Morphol. 2009, 270, 775–804. [Google Scholar] [CrossRef]
- Ørvig, T. Histological studies of ostracoderms, placoderms and fossil elasmobranchs. 3. Structure and growth of the gnathalia of certain arthrodires. Zool. Scr. 1980, 9, 141–159. [Google Scholar] [CrossRef]
- Dunkle, D.H.; Bungart, P.A. On one of the least known of the Cleveland Shale Arthrodira. Kirtlandia 1940, 8, 29–47. [Google Scholar]
- Straka, J.J.; Semken, H.A. A dinichthyid in Middle Devonian of Iowa. J. Paleontol. 1969, 43, 1423–1428. [Google Scholar]
- Vézina, D. Les plaques gnathales de Plourdosteus canadensis (Placodermi, Arthrodira) du Dévonien superieur du Quebec (Canada): Remarques sur la croissance dentaire et la mecanique masticatrice. Bull. Muséum Natl. Hist. Naturelle. Sect. C Sci. TerrePaleontol. Geol. Mineral. 1986, 4, 367–391. [Google Scholar] [CrossRef]
- Fabrezi, M.; Emerson, S.B. Parallelism and convergence in anuran fangs. J Zool 2003, 260, 41–51. [Google Scholar] [CrossRef]
- Trinajstic, K.; Dennis-Bryan, K. Phenotypic plasticity, polymorphism and phylogeny within placoderms. Acta Zool. 2009, 90, 83–102. [Google Scholar] [CrossRef]
- Rücklin, M.; Donoghue, P.C.J.; Johanson, Z.; Trinajstic, K.; Marone, F.; Stampanoni, M. Development of teeth and jaws in the earliest jawed vertebrates. Nature 2012, 491, 748–751. [Google Scholar] [CrossRef]
- Dunkle, D.H.; Bungart, P.A. The antero-supragnathal of Gorgonichthys. Am. Mus. Novit. 1946, 1316, 1–10. [Google Scholar]
- Dunkle, D.H.; Bungart, P.A. The infero-gnathal plates of Titanichthys. Kirtlandia 1942, 8, 49–59. [Google Scholar]
- Miles, R.S.; Westoll, T.S. The Placoderm Fish Coccosteus cuspidatus Miller ex Agassiz from the Middle Old Red Sandstone of Scotland. Part I. Descriptive Morphology. Trans. R. Soc. Edinb. 1968, 67, 373–476. [Google Scholar] [CrossRef]
- Boyle, J.; Ryan, M.J. New information on Titanichthys (Placodermi, Arthrodira) from the Cleveland Shale (Upper Devonian) of Ohio, USA. J. Paleontol. 2017, 91, 318–336. [Google Scholar] [CrossRef]
- Dunkle, D.H.; Bungart, P.A. Comments on Diplognathus mirabilis Newberry. Kirtlandia 1943, 8, 73–84. [Google Scholar]
- Dennis, K.I.M.; Miles, R.S. New durophagous arthrodires from Gogo, Western Australia. Zool. J. Linn. Soc. 1980, 69, 43–85. [Google Scholar] [CrossRef]
- Carr, R.K. Stenosteus angustopectus sp. nov. from the Cleveland Shale (Fammenian) of northern Ohio with a review of selenosteid (Placodermi) systematics. Kirtlandia 1996, 49, 19–43. [Google Scholar]
- Lelièvre, H. Description of Maidenia falipoui n. g., n. sp., a long-snouted brachythoracid (Vertebrata, Placodermi, Arthrodira) from the Givetian of Maider. Bull. Muséum Natl. Hist. Naturelle. Sect. C Sci. TerrePaleontol. Geol. Mineral. 1995, 17, 163–2067. [Google Scholar]
- Zhu, Y.-A.; Zhu, M.; Wang, J.-Q. Redescription of Yinostius major (Arthrodira: Heterostiidae) from the Lower Devonian of China, and the interrelationships of Brachythoraci. Zool. J. Linn. Soc. 2016, 176, 806–834. [Google Scholar] [CrossRef]
- Denison, R. Early devonian fishes from Utah. Pt. III. Arthrodira. Fieldiana Geol. 1958, 11, 461–551. [Google Scholar]
- Hussakof, L. Studies on the Arthrodira. Mem. Am. Mus. Nat. Hist. 1906, 9, 103–155. [Google Scholar]
- Hlavin, W.J. Biostratigraphy of the Late Devonian Black Shales on the Cratonal Margin of the Appalachian Geosyncline. Ph.D. Thesis, Boston University Graduate School, Ann Arbor, MI, USA, 1976. [Google Scholar]
- Johanson, Z.; Smith, M.M. Placoderm fishes, pharyngeal denticles, and the vertebrate dentition. J. Morphol. 2003, 257, 289–307. [Google Scholar] [CrossRef]
- Prondvai, E.; Stein, K.H.W.; De Ricqlès, A.; Cubo, J. Development-based revision of bone tissue classification: The importance of semantics for science. Biol. J. Linn. Soc. 2014, 112, 799–816. [Google Scholar] [CrossRef]
- Hussakof, L. The systematic relationships of certain American arthrodires. Bull. Am. Mus. Nat. Hist. 1909, 26, 263–272. [Google Scholar]
- Lelievre, H.; Janvier, P.; Goujet, D. Les vertébrés dévoniens de l’Iran central: IV: Arthrodires et ptyctodontes. Geobios 1981, 14, 677–709. [Google Scholar] [CrossRef]
- Dunkle, D.H.; Bungart, P.A. A new genus and species of arthrodire from the Cleveland Shale. Kirtlandia 1942, 8, 65–69. [Google Scholar]
- Dunkle, D.H.; Lane, N.G. Devonian fishes from California. Kirtlandia 1971, 15, 1–5. [Google Scholar]
- Dunkle, D.H.; Wilson, J.A. Remains of Devonian fishes from Texas. J. Wash. Acad. Sci. 1952, 42, 213–215. [Google Scholar]
- Szrek, P.; Wilk, O. A large Late Devonian arthrodire (Vertebrata, Placodermi) from Poland. Est. J. Earth Sci. 2018, 67, 33–42. [Google Scholar] [CrossRef]
- Lelièvre, H. Ardennosteus ubaghsi n.g., n. sp. Brachythoraci primitif (vertébré, placoderme) du Famennien d’Esneux (Belgique). Ann. Soc. Géol. Belg. 1982, 105, 1–7. [Google Scholar]
- Hussakof, L.; Bryant, W.L. Catalog of the fossil fishes in the museum of the Buffalo society of natural sciences. Bull. Buffalo Soc. Nat. Sci. 1918, 12, 1–198. [Google Scholar]
- Lehman, J.P. Les arthrodires du Dévonien supérieur du Tafilalet (sud marocain). Notes Mémoires. Serv. Géol. Maroc 1956, 129, 1–170. [Google Scholar]
- Rücklin, M. New finds of placoderms from the late devonian of Morocco. In Proceedings of the 7th European Workshop on Vertebrate Palaeontology, Sibiu, Romania, 2–7 July 2002; p. 31. [Google Scholar]
- Lehman, J.P. Nouveau arthrodires du Tafilalet et de ses environs. Ann. De Paléontologie (Vertébrés) 1977, 63, 105–119. [Google Scholar]
- Carr, R.K.; Hlavin, W.J. Dinichthyidae (Placodermi): A paleontological fiction? Geobios 1995, 28, 85–87. [Google Scholar] [CrossRef]
- Engelman, R.K. Giant, swimming mouths: Oral dimensions of extant sharks do not accurately predict body size in Dunkleosteus terrelli (Placodermi: Arthrodira). PeerJ 2023, 11, e15131. [Google Scholar] [CrossRef]
- Naylor, G.J.P.; Marcus, L.F. Identifying isolated shark teeth of the genus Carcharhinus to species: Relevance for tracking phyletic change through the fossil record. Am. Mus. Novit. 1994, 3109, 1–53. [Google Scholar]
- Garrick, J.A.F. Sharks of the genus Carcharhinus. In NOAA Technical Report NMFS CIRC; NMFS (National Marine Fisheries Service): Woods Hole, MA, USA, 1982; Volume 445, pp. 1–194. [Google Scholar]
- Thulborn, R.A. Thegosis in herbivorous dinosaurs. Nature 1974, 250, 729–731. [Google Scholar] [CrossRef]
- Trinajstic, K.; Hazelton, M. Ontogeny, phenotypic variation and phylogenetic implications of arthrodires from the Gogo Formation, Western Australia. J. Vertebr. Paleontol. 2007, 27, 571–583. [Google Scholar] [CrossRef]
- Carr, R.K. Reanalysis of Heintzichthys gouldii (Newberry), an aspinothoracid arthrodire (Placodermi) from the Famennian of northern Ohio, with a review of brachythoracid systematics. Zool. J. Linn. Soc. 1991, 103, 349–390. [Google Scholar] [CrossRef]
- Hanke, G.F.; Stewart, K.W.; Lammers, G.E. Squamatognathus steeprockensis gen. et sp. nov., an arthrodire inferognathal from the Middle Devonian Elm Point Formation of Manitoba. J. Vertebr. Paleontol. 1996, 16, 617–622. [Google Scholar] [CrossRef]
- Everts, V.; Delaissé, J.M.; Korper, W.; Jansen, D.C.; Tigchelaar-Gutter, W.; Saftig, P.; Beertsen, W. The Bone Lining Cell: Its Role in Cleaning Howship’s Lacunae and Initiating Bone Formation. J. Bone Miner. Res. 2002, 17, 77–90. [Google Scholar] [CrossRef]
- Zanno, L.E.; Makovicky, P.J. Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proc. Natl. Acad. Sci. USA 2011, 108, 232–237. [Google Scholar] [CrossRef]
- Sues, H.-D.; Reisz, R.R. Origins and early evolution of herbivory in tetrapods. Trends Ecol. Evol. 1998, 13, 141–145. [Google Scholar] [CrossRef]
- Thiery, A.P.; Shono, T.; Kurokawa, D.; Britz, R.; Johanson, Z.; Fraser, G.J. Spatially restricted dental regeneration drives pufferfish beak development. Proc. Natl. Acad. Sci. USA 2017, 114, E4425–E4434. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Underwood, C.; Goral, T.; Healy, C.; Johanson, Z. Growth and mineralogy in dental plates of the holocephalan Harriotta raleighana (Chondrichthyes): Novel dentine and conserved patterning combine to create a unique chondrichthyan dentition. Zool. Lett. 2019, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, O.A.; Johanson, Z.; Kuznetsov, A.N.; Tsessarsky, A.; Trinajstic, K.; Isakhodzayev, F.B. Feeding in the Devonian antiarch placoderm fishes: A study based upon morphofunctional analysis of jaws. J. Paleontol. 2022, 96, 1413–1430. [Google Scholar] [CrossRef]
- De Buffrénil, V.; Quilhac, A. Bone remodeling. In Vertebrate Skeletal Histology and Paleohistology; De Buffrénil, V., De Ricqlés, A., Zylberberg, L., Padian, K., Eds.; CRC Press: Boca Raton, CA, USA, 2021; pp. 229–246. [Google Scholar]
- Giles, S.; Rücklin, M.; Donoghue, P.C.J. Histology of “placoderm” dermal skeletons: Implications for the nature of the ancestral gnathostome. J. Morphol. 2013, 274, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.M.; Summers, A.P. The material properties of acellular bone in a teleost fish. J. Exp. Biol. 2009, 212, 1413–1420. [Google Scholar] [CrossRef]
- Davesne, D.; Meunier, F.J.; Schmitt, A.D.; Friedman, M.; Otero, O.; Benson, R.B.J. The phylogenetic origin and evolution of acellular bone in teleost fishes: Insights into osteocyte function in bone metabolism. Biol. Rev. 2019, 94, 1338–1363. [Google Scholar] [CrossRef]
- Fabbri, M.; Navalón, G.; Benson, R.B.J.; Pol, D.; O’Connor, J.; Bhullar, B.-A.S.; Erickson, G.M.; Norell, M.A.; Orkney, A.; Lamanna, M.C.; et al. Subaqueous foraging among carnivorous dinosaurs. Nature 2022, 603, 852–857. [Google Scholar] [CrossRef]
- Houssaye, A.; Martin Sander, P.; Klein, N. Adaptive Patterns in Aquatic Amniote Bone Microanatomy-More Complex than Previously Thought. Integr. Comp. Biol. 2016, 56, 1349–1369. [Google Scholar] [CrossRef]
- De Buffrénil, V.; De Ricqlès, A.; Ray, C.E.; Domning, D.P. Bone histology of the ribs of the archaeocetes (Mammalia: Cetacea). J. Vertebr. Paleontol. 1990, 10, 455–466. [Google Scholar] [CrossRef]
- Allis, E.P. The so-called mandibular artery and the persisting remnant of the mandibular aortic arch in the adult selachian. J. Morphol. 1916, 27, 99–118. [Google Scholar] [CrossRef]




















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebedev, O.A.; Engelman, R.K.; Skutschas, P.P.; Johanson, Z.; Smith, M.M.; Kolchanov, V.V.; Trinajstic, K.; Linkevich, V.V. Structure, Growth and Histology of Gnathal Elements in Dunkleosteus (Arthrodira, Placodermi), with a Description of a New Species from the Famennian (Upper Devonian) of the Tver Region (North-Western Russia). Diversity 2023, 15, 648. https://doi.org/10.3390/d15050648
Lebedev OA, Engelman RK, Skutschas PP, Johanson Z, Smith MM, Kolchanov VV, Trinajstic K, Linkevich VV. Structure, Growth and Histology of Gnathal Elements in Dunkleosteus (Arthrodira, Placodermi), with a Description of a New Species from the Famennian (Upper Devonian) of the Tver Region (North-Western Russia). Diversity. 2023; 15(5):648. https://doi.org/10.3390/d15050648
Chicago/Turabian StyleLebedev, Oleg A., Russell K. Engelman, Pavel P. Skutschas, Zerina Johanson, Moya M. Smith, Veniamin V. Kolchanov, Kate Trinajstic, and Valeriy V. Linkevich. 2023. "Structure, Growth and Histology of Gnathal Elements in Dunkleosteus (Arthrodira, Placodermi), with a Description of a New Species from the Famennian (Upper Devonian) of the Tver Region (North-Western Russia)" Diversity 15, no. 5: 648. https://doi.org/10.3390/d15050648
APA StyleLebedev, O. A., Engelman, R. K., Skutschas, P. P., Johanson, Z., Smith, M. M., Kolchanov, V. V., Trinajstic, K., & Linkevich, V. V. (2023). Structure, Growth and Histology of Gnathal Elements in Dunkleosteus (Arthrodira, Placodermi), with a Description of a New Species from the Famennian (Upper Devonian) of the Tver Region (North-Western Russia). Diversity, 15(5), 648. https://doi.org/10.3390/d15050648