



Ecological Basis of Ecosystem Services and Management of Wetlands Dominated by Common Reed (Phragmites australis): European Perspective


Abstract
1. Introduction
2. The Genetic Delineation and Ecological Niche of P. australis in Europe
3. Vegetation with Phragmites australis
3.1. General Overview
3.2. Regional Survey
- Freshwater reedbeds usually hosting species-poor plant communities including P. australis, other marsh dominants such as Typha latifolia, T. angustifolia, Schoenoplectus lacustris, Bolboschoenus maritimus and tall sedges.
- Tall-herb species-rich fens with Cladium mariscus and Calamagrostis canescens or some other species (Juncus subnodulosus, Carex elata, C. acutifomis, C. appropinquata, C. lasiocarpa, C. diandra) as co-dominants.
- Saline brackish marshes in which more halophlous species such as Atriplex prostrata, Juncus gerardii, and Aster tripolium co-occur with P. australis.
- A tall-herb vegetation of abandoned moist-to-wet meadows, including tall herbaceous dicotyledons such as Eupatorium cannabinum, Angelica sylvestris, Lythrum salicaria, Cirsium palustre, Filipendula ulmaria, and Epilobium hirsutum.
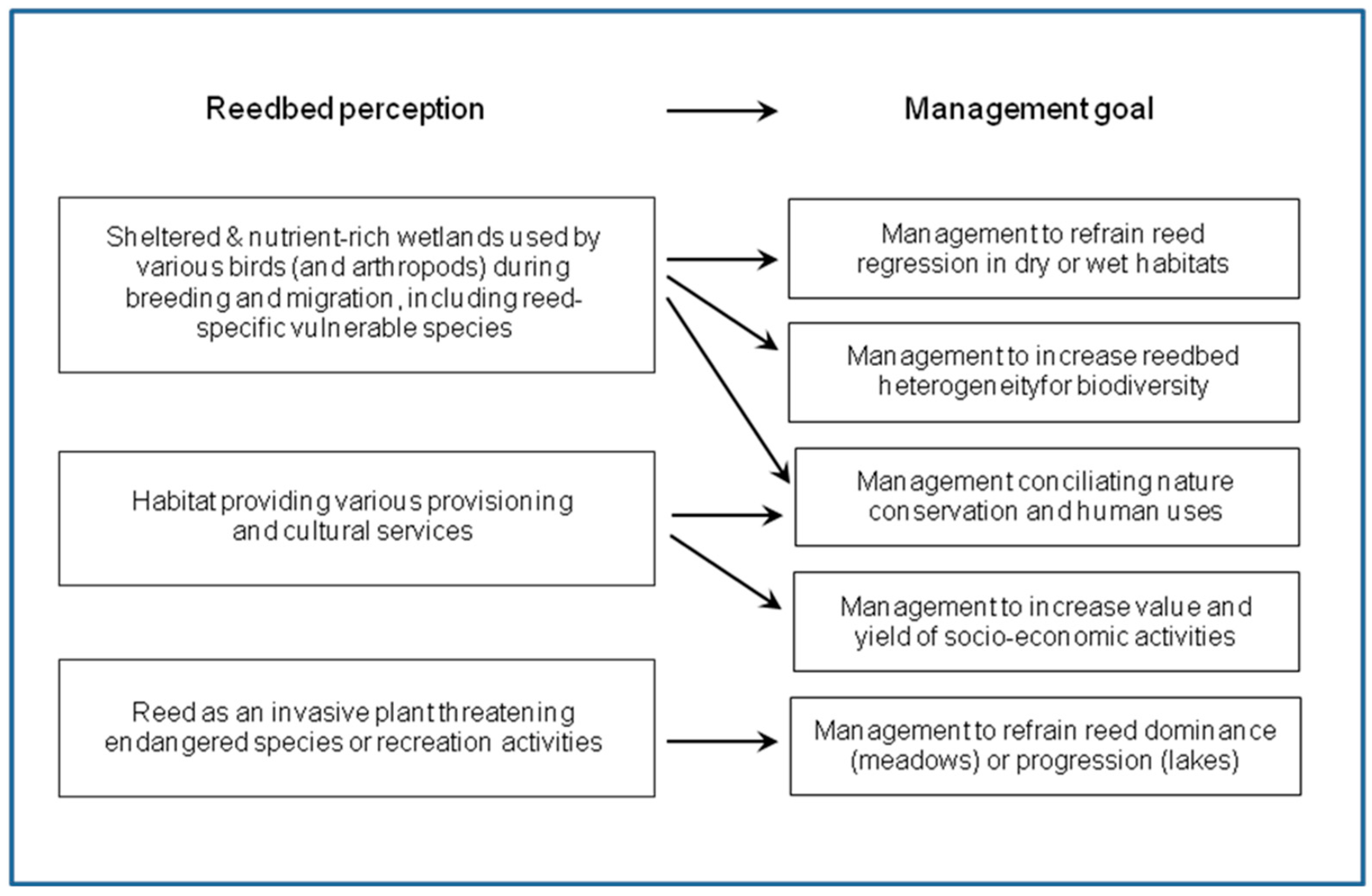
4. Use and Management of P. australis Habitats for Biodiversity
4.1. P. australis Stands as Habitats of Birds and Invertebrates
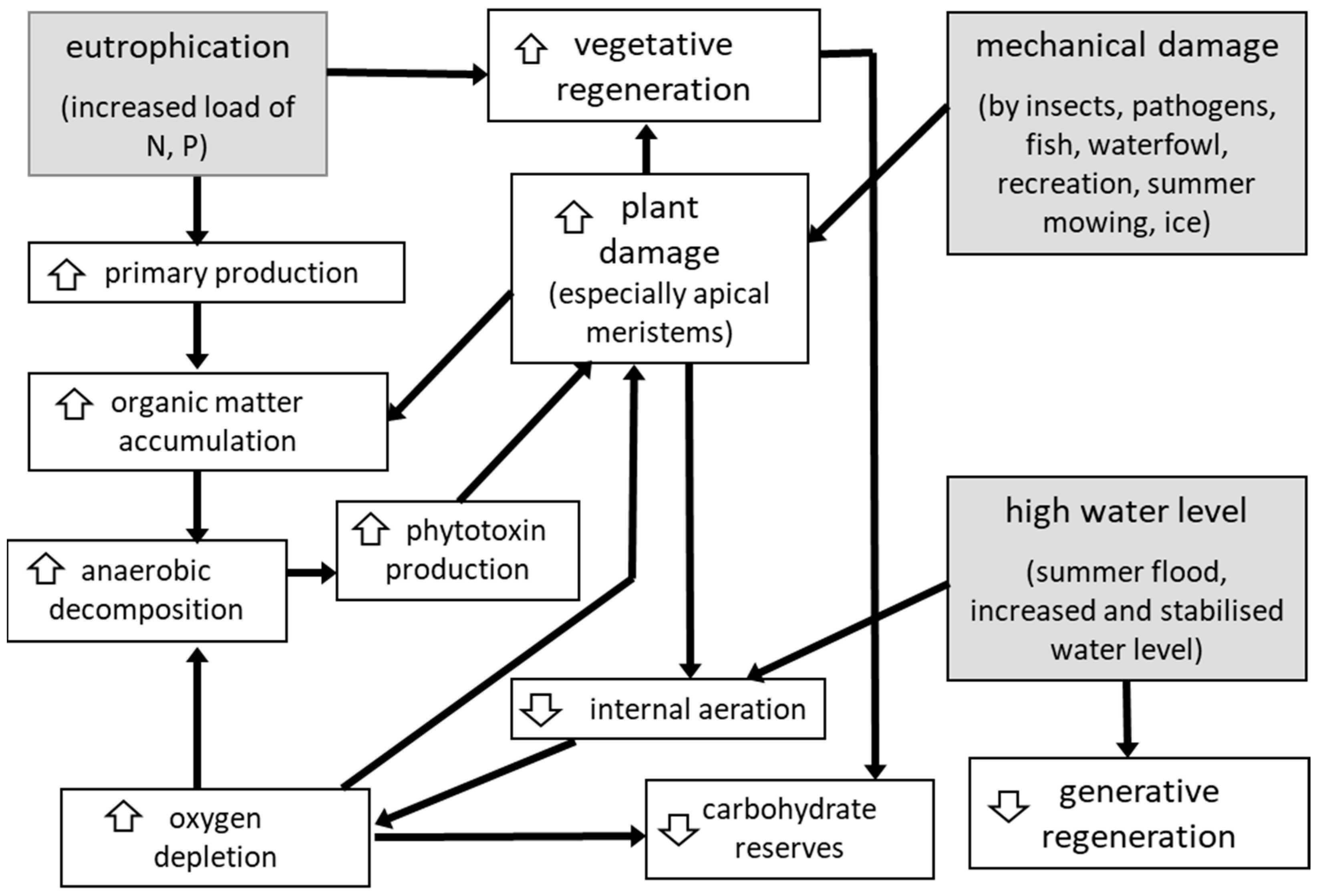
4.2. Management to Stop P. australis Regression in Dry Habitats
4.3. Management to Revert the Regression of Reed in Wet Habitats
- More efficient purification of wastewater discharged into the lake [104];
- Reduction of nutrient input from neighbouring agricultural areas [105];
- Increased nutrient stripping in the inflowing water by enhancing the mineral nutrient uptake by a dense water and bank vegetation upstream; its thereby enhanced cumulative nutrient uptake deprives the reeds growing downstream of a part of their mineral nutrient supply [106];
- Removal of accumulated nutrient-rich mud by suction dredging [102].
4.4. Management to Increase Reedbed Heterogeneity
4.5. Management to Stop the Spread of Reed in Wet Grasslands
5. Use and Management for Direct Economic Benefits
5.1. Overview of Economic Benefits
5.2. Management for Reed Harvesting
5.3. Management for Waterfowl Hunting
5.4. Sustainable Grazing
5.5. Compatibility with Fish Farming
6. Restoration and Construction of P. australis-Dominated Wetlands
6.1. Rewetting of Agricultural Peat Soils
6.2. Constructed Wetlands for Wastewater Treatment
7. Multiple Uses
8. Future Prospects of P. australis in Europe
9. Conclusions
- This review of knowledge on European P. australis populations indicates that it is a plastic and versatile species, forming part of varied plant communities all over Europe.
- The analysis of the ecophysiological response to multiple stressors is used as a tool for understanding the population dynamics of P. australis in the main habitat types in Europe. Its decline at deep-water sites, stable performance in constructed wetlands with subsurface horizontal flow and expansion in wet grasslands are given as examples.
- Of various human uses, the role of P. australis as a habitat former has gained an increasing value. Vulnerable birds are major drivers of reedbed management, especially in northwestern Europe, where large reedbeds have deteriorated or disappeared, which was followed by intensive habitat management (‘gardening’), restoration and creation. Traditional socioeconomic uses are being abandoned, intensified or replaced by more lucrative activities (e.g., waterfowl hunting). Uses of common reed as energy crop and renewable eco-material for green buildings are limited but promising.
- Each of the uses should be based on management practices that include both natural and human-driven processes. Nevertheless, the long-term maintenance or intensification of the economic uses often leads to practices that are not sustainable and get into conflict with nature conservation. Harmonisation of multiple uses with the help of innovative approaches (modelling) can assure a more sustainable future of P. australis wetlands.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- den Hartog, C.; Květ, J.; Sukopp, H. Reed. A common species in decline. Aquat. Bot. 1989, 35, 1–4. [Google Scholar] [CrossRef]
- Kiviat, E. Organisms using Phragmites australis are diverse and similar on three continents. J. Nat. Hist. 2019, 53, 1975–2010. [Google Scholar] [CrossRef]
- Köbbing, J.F.; Thevs, N.; Zerbe, S.; Wichtmann, W.; Couwenberg, J. The utilisation of reed (Phragmites australis): A review. Mires Peat 2013, 13, 1–14. [Google Scholar]
- Koppitz, H. Analysis of genetic diversity among selected populations of Phragmites australis world-wide. Aquat. Bot. 1999, 64, 209–221. [Google Scholar] [CrossRef]
- Lambertini, C.; Gustafsson, M.H.G.; Frydenberg, J.; Lissner, J.; Speranza, M.; Brix, H. A phylogeographic study of the cosmopolitan genus Phragmites (Poaceae) based on AFLPs. Plant Syst. Evol. 2006, 258, 161–182. [Google Scholar] [CrossRef]
- Tho, B.T.; Sorrell, B.K.; Lambertini, C.; Eller, F.; Brix, H. Phragmites australis: How do genotypes of different phylogeographic origins differ from their invasive genotypes in growth, nitrogen allocation and gas exchange? Biol. Invasions 2016, 18, 2563–2576. [Google Scholar] [CrossRef]
- Pyšek, P.; Skálová, H.; Čuda, J.; Guo, W.; Doležal, J.; Kauzál, O.; Lambertini, C.; Pyšková, K.; Brix, H.; Meyerson, L.A. Physiology of a plant invasion: Biomass production, growth and tissue chemistry of invasive and native Phragmites australis populations. Preslia 2019, 91, 51–75. [Google Scholar] [CrossRef]
- Kettenring, K.M.; McCormick, M.K.; Baron, H.M.; Whigham, D.F. Mechanisms of Phragmites australis invasion: Feedbacks among genetic diversity, nutrients, and sexual reproduction. J. Appl. Ecol. 2011, 48, 1305–1313. [Google Scholar] [CrossRef]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Guo, W.Y.; Guo, X.; Hazelton, E.L.; Kettenring, K.M.; et al. Cosmopolitan species as models for ecophysiological responses to global change: The common reed Phragmites australis. Front. Plant Sci. 2017, 8, 1833. [Google Scholar] [CrossRef]
- Hazelton, E.L.; Mozdzer, T.J.; Burdick, D.M.; Kettenring, K.M.; Whigham, D.F. Phragmites australis management in the United States: 40 years of methods and outcomes. AoB Plants 2014, 6, plu001. [Google Scholar] [CrossRef] [PubMed]
- De Groot, R.S.; Wilson, M.A.; Boumans, R.M. A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecol. Econ. 2002, 41, 393–408. [Google Scholar] [CrossRef]
- Marks, M.; Lapin, B.; Randall, J. Phragmites australis (p communis): Threats, management, and monitoring. Nat. Areas J. 1994, 14, 285–294. [Google Scholar]
- Güsewell, S.; Klötzli, F. Assessment of aquatic and terrestrial reed (Phragmites australis) stands. Wetl. Ecol. Manag. 2000, 8, 367–373. [Google Scholar] [CrossRef]
- Ludwig, D.E.; Iannuzzi, T.J.; Esposito, A.N. Phragmites and environmental management: A question of values. Estuaries 2003, 26, 624–630. [Google Scholar] [CrossRef]
- Valkamaa, E.; Lyytinena, S.; Koricheva, J. The impact of reed management on wildlife: A meta-analytical review of European studies. Biol. Conserv. 2008, 141, 364–374. [Google Scholar] [CrossRef]
- Haslam, S.M. A Book of Reed: (Phragmites australis (Cav.) Trin. ex Steudel, Formerly Phragmites Communis Trin.); Forrest Press: Cardigan, UK, 2010. [Google Scholar]
- Lambertini, C.; Sorrell, B.K.; Riis, T.; Olesen, B.; Brix, H. Exploring the borders of European Phragmites within a cosmopolitan genus. AoB Plants 2012, pls020. [Google Scholar] [CrossRef]
- Clayton, W.D. The correct name of the common reed. Taxon 1968, 17, 168–169. [Google Scholar] [CrossRef]
- Björk, S. Ecological investigations of Phragmites communis. Folia Limnol. Scand. 1967, 14, 1–248. [Google Scholar]
- Rodewald-Rudescu, L. Das Schilfrohr, Phragmites communis Trin. In Binnengewässer Bd. 27; Elster, H.D., Ohle, J.W., Eds.; Schweizerbart’scher Verlag: Stuttgart, Germany, 1974; 307p. [Google Scholar]
- Haslam, S.M. Biological flora of the British Isles. Phragmites communis Trin. J. Ecol. 1972, 60, 585–610. [Google Scholar]
- Packer, J.G.; Meyerson, L.A.; Skálová, H.; Pyšek, P.; Kueffer, C. Biological flora of the British Isles: Phragmites australis. J. Ecol. 2017, 105, 1123–1162. [Google Scholar] [CrossRef]
- Mal, T.K.; Narine, L. The biology of Canadian weeds. 129. Phragmites australis (Cav.) Trin. ex Steud. Can. J. Plant Sci. 2004, 84, 365–396. [Google Scholar] [CrossRef]
- Engloner, A.I. Structure, growth dynamics and biomass of reed (Phragmites australis)—A review. Flora 2009, 204, 331–346. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Hejný, S.; Remy, D.; Pott, R. Tall reed vegetation and tall sedge swamps, aquatic vegetation. In Karte der Natürlichen Vegetation Europas/Map of the Natural Vegetation of Europe. Maßstab/Scale 1:2,500,000; Bohn, U., Gollub, G., Hettwer, C., Neuhäuslová, Z., Raus, T., Schlüter, H., Eds.; Bundesamt für Naturschutz: Bonn, Germany, 2004; pp. 437–449. [Google Scholar]
- Bohn, U.; Gollub, G.; Hettwer, C.; Neuhäuslová, Z.; Raus, T.; Schlüter, H. (Eds.) Karte der Natürlichen Vegetation Europas/Map of the Natural Vegetation of Europe. Maßstab/Scale 1:2,500,000; Bundesamt für Naturschutz: Bonn, Germany, 2004. [Google Scholar]
- Hobbs, R.J.; Arico, S.; Aronson, J.; Baron, J.S.; Bridgewater, P.; Cramer, V.A.; Epstein, P.R.; Ewel, J.J.; Klink, C.A.; Lugo, A.E.; et al. Novel ecosystems: Theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 2006, 15, 1–7. [Google Scholar] [CrossRef]
- Solomeshch, A.I.; Mirkin, B.M.; Ermakov, N.; Ishbirdin, A.; Golub, V.; Saitov, M.; Zhuravliova, S.; Rodwell, J. Red Data Book of Plant Communities in the Former USSR; Unit of Vegetation Science: Lancaster, UK, 1997. [Google Scholar]
- Chytrý, M.; Hennekens, S.M.; Jiménez-Alfaro, B.; Knollová, I.; Dengler, J.; Jansen, F.; Landucci, F.; Schaminée, J.H.; Aćić, S.; Agrillo, E.; et al. European Vegetation Archive (EVA): An integrated database of European vegetation plots. Appl. Veg. Sci. 2016, 19, 173–180. [Google Scholar] [CrossRef]
- Rodwell, J. (Ed.) British Plant Communities; Cambridge University Press: Cambridge, UK, 2000; Volume 1–5. [Google Scholar]
- Schaminée, J.H.J.; Stortelder, A.H.F.; Westhoff, V.; Weeda, E.J.; Hommel, P.W.F.M. (Eds.) De Vegetatie van Nederland. Deel 1–5; Opulus Press: Uppsala, Sweden; Leiden, The Netherlands, 1995–1999. [Google Scholar]
- Dierssen, K. Vegetation Nordeuropas; Verlag Eugen Ulmer: Stuttgart, Germany, 1996. [Google Scholar]
- Pott, R. Die Pflanzengesellschften Deutschlands; Verlag Eugen Ulmer: Stuttgart, Germany, 1995. [Google Scholar]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roslinnych Polski; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2002. [Google Scholar]
- Chytrý, M. (Ed.) Vegetation of the Czech Republic; Academia: Praha, Czech Republic, 2007–2011; Volume 1–3. (In Czech) [Google Scholar]
- Mucina, L.; Grabherr, G.; Ellmauer, T.; Wallnöfer, S. (Eds.) Die Pflanzengesellschaften Österreichs 1–3; Gustav Fischer: Jena, Germany, 1993. [Google Scholar]
- Borhidi, A. An annotated checklist of the Hungarian plant communities 1. The non-forest vegetation. In Critical Revision of the Hungarian Plant Communities; Borhidi, A., Ed.; Janus Pannonius University: Pécs, Hungary, 1996; pp. 43–94. [Google Scholar]
- Pomogyi, P. Changes in the Reed Communities of Lake Fenéki in the Little Balaton Protection System Based on the Results of the Vegetation Mapping; Research Report; West Danube Water Directorate: Készthély, Hungary, 1996. (In Hungarian) [Google Scholar]
- Horvat, I.; Glavač, V.; Ellenberg, H. Vegetation Südosteuropas; Gustav Fischer Verlag: Stuttgart, Germany, 1974. [Google Scholar]
- Stančić, Z. Marshland vegetation of the class Phagmito-Magnocaricetea in Croatia. Biologia 2007, 62, 297–314. [Google Scholar] [CrossRef]
- Solomakha, V.A. The Syntaxonomy of Vegetation of the Ukraine; Ukrainian Phytosociological Centre: Kyiv, Ukraine, 1996. [Google Scholar]
- Shelyag-Sosonko, J.R. Zelenaya kniga Ukrajinskoi SSR; Naukova Dumka: Kiev, Ukraine, 1987. (In Russian) [Google Scholar]
- Golub, V.B.; Mirkin, B.M. Grasslands of the lower Volga valley. Folia Geobot. Et Phytotaxon. 1986, 21, 337–395. [Google Scholar] [CrossRef]
- Raspopov, I.M. Higher Aquatic Plant Communities of the Great Lakes in the Northwestern USSR; Izdatelstvo Nauka: Leningrad, Russia, 1985. (In Russian) [Google Scholar]
- Gejny, S.; Sytnik, K.M. Macrophytes as Environmental Indicators; Naukova Dumka: Kiev, Ukraine, 1993. (In Russian) [Google Scholar]
- Rivas-Martínez, S.; Díaz, T.E.; Fernández-González, F.; Izco, J.; Loidi, J.; Lousã, M.; Penas, Á. Vascular plant communities of Spain and Portugal. Addenda to the Syntaxonomical checklist of 2001. Itinera Geobot. 2002, 15, 5–922. [Google Scholar]
- Landucci, F.; Gigante, D.; Venanzoni, R.; Chytrý, M. Wetland vegetation of the class Phragmito-Magno-Caricetea in central Italy. Phytocoenologia 2013, 43, 67–100. [Google Scholar] [CrossRef]
- Hutchinson, G.E. A treatise on limnology. In Limnological Botany; John Wiley & Sons: New York, NY, USA; London, UK; Sydney, Australia; Toronto, ON, Canada, 1977; Volume 3, 660p. [Google Scholar]
- Hroudová, Z.; Zákravský, P. Vegetation dynamics in a fishpond littoral related to human impact. Hydrobiologia 1999, 415, 139–145. [Google Scholar] [CrossRef]
- Güsewell, S. Management of Phragmites australis in Swiss fen meadows by mowing in early summer. Wetl. Ecol. Manag. 2003, 11, 433–445. [Google Scholar] [CrossRef]
- Rudescu, L. Neue biologische Probleme bei den Phragmiteskulturarbeiten im Donaudelta. Arch. Für Hydrobiol. Suppl. 1965, 30, 80–111. [Google Scholar] [CrossRef]
- Kokin, K.A. Ecology of Higher Aquatic Plants; Izdatelstvo Moskovskogo Universiteta: Moskva, Russia, 1982. (In Russian) [Google Scholar]
- Dubyna, D.V.; Neuhäuslová, Z.; Šeljag-Sosonko, J.R. Coastal vegetation of the Birjučij Island Spit in the Azov Sea, Ukraine. Preslia 1994, 66, 193–216. [Google Scholar]
- Antonellini, M.; Mollema, P.N. Impact of groundwater salinity on vegetation species richness in the coastal pine forests and wetlands of Ravenna, Italy. Ecol. Eng. 2010, 36, 1201–1211. [Google Scholar] [CrossRef]
- Giambastiani, B.M.; Greggio, N.; Nobili, G.; Dinelli, E.; Antonellini, M. Forest fire effects on groundwater in a coastal aquifer (Ravenna, Italy). Hydrol. Process. 2018, 32, 2377–2389. [Google Scholar] [CrossRef]
- Dobrowolski, K.A. Role of birds in Polish wetland ecosystems. Pol. Arch. Hydrobiol. 1973, 20, 217–221. [Google Scholar]
- Bibby, C.J.; Lunn, J. Conservation of reed beds and their avifauna in England and Wales. Biol. Conserv. 1982, 23, 167–186. [Google Scholar] [CrossRef]
- Burgess, N.D.; Evans, C.E. The Management of Reedbeds for Birds; The Royal Society for the Protection of Birds: Sandy, UK, 1989. [Google Scholar]
- Andrews, J.; Ward, D. The management and creation of Reedbeds—Especially for rare birds. Br. Wildl. 1991, 3, 81–91. [Google Scholar]
- Fojt, W.; Foster, A. Reedbeds, their wildlife and requirements. A: Botanical and invertebrate aspects of reedbeds, their ecological requirements and conservation significance. In Reedbeds for Wildlife; Ward, D., Ed.; The Royal Society of Protection for Birds: Oxford, UK; University of Bristol Information Press: Oxford, UK, 1992; pp. 49–56. [Google Scholar]
- Tyler, G. Reedbeds their wildlife requirements, B. Requirements of birds in reedbeds. In Reedbeds for Wildlife; Ward, D., Ed.; The Royal Society for the Protection of Birds: Oxford, UK; University of Bristol Information Press: Oxford, UK, 1992; pp. 57–64. [Google Scholar]
- Graveland, J. Effects of reed cutting on density and breeding success of Reed Warbler Acrocephalus scirpaceus and Sedge Warbler Acrocephalus schoenobaenus. J. Avian Biol. 1999, 30, 469–482. [Google Scholar] [CrossRef]
- Báldi, A.; Kisbenedek, T. Bird species numbers in an archipelago of reeds at Lake Velence, Hungary. Glob. Ecol. Biogeogr. 2000, 9, 451–461. [Google Scholar] [CrossRef]
- Barbraud, C.; Lepley, M.; Mathevet, R.; Mauchamp, A. Reedbed selection and colony size of breeding purple herons Ardea purpurea in southern France. Ibis 2002, 144, 227–235. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Bertolero, A.; Bigas, D.; Paquet, J.Y.; Martinez-Vitalta, A. Habitat selection of passerine birds nesting in the Ebro Delta reedbeds (NE Spain): Management implications. Wetlands 2002, 22, 318–325. [Google Scholar] [CrossRef]
- Poulin, B.; Lefebvre, G. Effect of winter cutting on the passerine breeding assemblage in French Mediterranean reedbeds. Biodivers. Conserv. 2002, 11, 1567–1581. [Google Scholar] [CrossRef]
- Adamo, M.C.; Puglisi, L.; Baldaccini, N.E. Factors affecting Bittern Botaurus stellaris distribution in a Mediterranean wetland. Bird Conserv. Int. 2004, 14, 153–164. [Google Scholar] [CrossRef]
- Gilbert, G.; Tyler, G.A.; Dunn, C.J.; Smith, K.W. Nesting habitat selection by bitterns Botaurus stellaris in Britain and the implications for wetland management. Biol. Conserv. 2005, 124, 547–553. [Google Scholar] [CrossRef]
- Self, M. A review of management for fish and bitterns, Botaurus stellaris, in wetland reserves. Fish. Manag. Ecol. 2005, 12, 387–394. [Google Scholar] [CrossRef]
- Pasinelli, G.; Schiegg, K. Fragmentation within and between wetland reserves: The importance of spatial scales for nest predation in reed buntings. Ecography 2006, 29, 731–732. [Google Scholar] [CrossRef]
- Poulin, B.; Lefebvre, G.; Allard, S.; Mathevet, R. Reed harvest and summer drawdown enhance bittern habitat in the Camargue. Biol. Conserv. 2009, 142, 689–695. [Google Scholar] [CrossRef]
- Provost, P.; Kerbiriou, C.; Jiguet, F. Foraging range and habitat use by Aquatic Warblers Acrocephalus paludicola during a fall migration stopover. Acta Ornithol. 2010, 45, 173–180. [Google Scholar] [CrossRef]
- Andersen, L.H.; Nummi, P.; Rafn, J.; Frederiksen, C.M.S.; Kristjansen, M.P.; Lauridsen, T.L.; Trojelsgaard, K.; Pertoldi, C.; Bruhn, D.; Bahrndorff, S. Can reed harvest be used as a management strategy for improving invertebrate biomass and diversity? J. Environ. Manag. 2021, 300, 113637. [Google Scholar] [CrossRef]
- Musseau, R.; Crépin, M.; Brugulat, C.; Kerbiriou, C. Conservation and Restoration of Coastal Reed Beds in the Context of Global Change: Potential Effects of Habitat Fragmentation for Specialist Marshland Passerines. Wetlands 2021, 41, 70. [Google Scholar] [CrossRef]
- Nemeth, E.; Dvorak, M. Reed die-back and conservation of small reed birds at Lake Neusiedl, Austria. J. Ornithol. 2022, 163, 683–693. [Google Scholar] [CrossRef]
- BirdLife International. European Birds of Conservation Concern: Populations, Trends and National Responsibilities; BirdLife International: Cambridge, UK, 2017. [Google Scholar]
- Smiddy, P.; Cullen, C.; O’Halloran, J. Time of roosting of Barn Swallows Hirundo rustica at an Irish reedbed during autumn migration. Ringing Migr. 2007, 23, 228–230. [Google Scholar] [CrossRef]
- Buckland, S.T.; Hereward, A.C. Trap shyness of Yellow Wagtails Motacilla flava flavissima at a premigratory roost. Ringing Migr. 1982, 4, 15–23. [Google Scholar] [CrossRef]
- Broyer, J.; Calenge, C. Influence of fish-farming management on duck breeding in French fish pond systems. Hydrobiologia 2010, 637, 173–185. [Google Scholar] [CrossRef]
- Wotton, S.; Brown, A.; Burn, A.; Dodd, A.; Droy, N.; Gilbert, G.; Hardiman, N.; Rees, S.; White, G.; Gregory, R. Boom or bust—A sustainable future for reedbeds and Bitterns? British Wildlife 2005, 20, 305–315. [Google Scholar]
- Information Sheet on Ramsar Wetlands. Broadland. Available online: http://jncc.defra.gov.uk/pdf/RIS/UK11010.pdf (accessed on 10 April 2023).
- Jacoby, H. Die Vögel des Bodenseegebietes; ALA/Schweizerische Gesellschaft für Vogelkunde und Vogelschutz: Sempach, Switzerland, 1970; 260p. [Google Scholar]
- Löffler, H. Neusiedlersee. The Limnology of a Shallow Lake in Central Europe. In Monographiae Biologicae; Dr. W. Junk bv: The Hague, The Netherlands, 1979; 543p. [Google Scholar]
- Dorogman, C. Information Sheet on Ramsar Wetlands: Lake Fertö. 2007. Available online: https://rsis.ramsar.org/RISapp/files/RISrep/HU420RISformer_150330.pdf (accessed on 10 April 2023).
- Metz, H. Information Sheet on Ramsar Wetlands: Neusiedler See–Seewinkel. 2005. Available online: https://rsis.ramsar.org/RISapp/files/RISrep/AT271RIS.pdf (accessed on 6 April 2012).
- Květ, J. Mineral nutrients in shoots of reed (Phragmites communis Trin.). Pol. Arch. Hydrobiol. 1973, 20, 137–147. [Google Scholar]
- Dykyjová, D.; Květ, J. (Eds.) Pond Littoral Ecosystems. Structure and Functioning; Ecological Studies 28; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1978; 464p. [Google Scholar]
- Information Sheet on Ramsar Wetlands: The Camargue. 1993. Available online: https://rsis.ramsar.org/RISapp/files/RISrep/FR786RISformer1993_EN.pdf (accessed on 10 April 2023).
- The Annotated Ramsar List: Romania. Available online: https://rsis.ramsar.org/sites/default/files/rsiswp_search/exports/Ramsar-Sites-annotated-summary-Romania.pdf?1680615226 (accessed on 10 April 2023).
- The Annotated Ramsar List: Hungary. Available online: https://rsis.ramsar.org/sites/default/files/rsiswp_search/exports/Ramsar-Sites-annotated-summary-Hungary.pdf?1622485211 (accessed on 10 April 2023).
- Dömötörfy, Z. Changes in macro-vegetation of the Kis-Balaton wetlands over the last two centuries: A GIS perspective. Hydrobiologia 2003, 506–509, 671–679. [Google Scholar] [CrossRef]
- Lee, S.Y. Net aerial primary productivity, litter production and decomposition of the reed Phragmites communis in a nature reserve in Hong Kong: Management implications. Mar. Ecol. Prog. Ser. 1990, 66, 161–174. [Google Scholar] [CrossRef]
- Cowie, N.R.; Sutherland, W.J.; Ditlhogo, M.K.M.; James, R. The effects of conservation management of reed beds II. The flora and litter disappearance. J. Appl. Ecol. 1992, 29, 277–284. [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Pieczyńska, E.; Úlehlová, B. Further fate of organic matter in wetlands. In The Production Ecology of Wetlands; Westlake, D.F., Květ, J., Szczepański, A., Eds.; Cambridge University Press: Cambridge, UK, 1978; pp. 169–191. [Google Scholar]
- Úlehlová, B. The role and fate of decomposers in wetlands. In The Production Ecology of Wetlands; Westlake, D.F., Květ, J., Szczepański, A., Eds.; Cambridge University Press: Cambridge, UK, 1998; pp. 192–210. [Google Scholar]
- van der Toorn, J.; Mook, J.H. The influence of environmental factors and management on stands of Phragmites australis. I. Effects of burning, frost, and insect damage on shoot density and shoot size. J. Appl. Ecol. 1982, 19, 477–499. [Google Scholar] [CrossRef]
- Ditlhogo, M.K.; James, M.R.; Laurence, B.R.; Sutherland, W.J. The effects of conservation management of reed beds I. The invertebrates. J. Appl. Ecol. 1992, 29, 265–276. [Google Scholar] [CrossRef]
- Ostendorp, W. Impact of winter reed harvesting and burning on the nutrient economy of reed beds. Wetl. Ecol. Manag. 1995, 3, 233–248. [Google Scholar] [CrossRef]
- Schulze Hagen, K.; Pleines, S.; Sennert, G. Rapid increase of Cuckoo parasitism in a local Reed Warbler population. Vogelwelt 1996, 117, 83–86. [Google Scholar]
- Hawke, C.J.; Jose, P.V. Reedbed Management for Commercial and Wildlife Interests; The Royal Society for the Protection of Birds: Sandy, UK, 1996. [Google Scholar]
- White, G.; Purps, J.; Alsbury, S. The Bittern in Europe: A Guide to Species and Habitat Management; The Royal Society for the Protection of Birds: Sandy, UK, 2006. [Google Scholar]
- Ostendorp, W. ‘Die-back’ of reeds—A critical review of literature. Aquat. Bot. 1989, 35, 5–26. [Google Scholar] [CrossRef]
- Kramer, I.; Kapfer, A. Naturnahe Uferbereiche und Flachwasserzonen des Bodensees. Biotope Baden-Württemberg 2001, 13, 1–47. [Google Scholar]
- HELCOM. The review of more specific targets to reach the goal set up in the 1988/1998 Ministerial Declarations regarding nutrients. Balt. Sea Environ. Proc. 2003, 89. [Google Scholar]
- Stamati, F.E.; Chalkias, N.; Moraetis, D.; Nikolaidis, N.P.F. Natural attenuation of nutrients in a Mediterranean drainage canal. J. Environ. Monit. 2010, 12, 164–171. [Google Scholar] [CrossRef]
- Alvarez, M.G.; Tron, F.; Mauchamp, A. Sexual versus asexual colonization by Phragmites australis: 25-year reed dynamics in a Mediterranean marsh, Southern France. Wetlands 2005, 25, 639–647. [Google Scholar] [CrossRef]
- Linthurst, R.A. The effect of aeration on the growth of Spartina alterniflora Loisel. Am. J. Bot. 1979, 66, 685–691. [Google Scholar] [CrossRef]
- van Wijck, C.; De Groot, C.J. The impact of desiccation of a freshwater marsh (Garcines Nord, Camargue, France) on sediment–water–vegetation interactions. Hydrobiologia 1993, 252, 95–103. [Google Scholar] [CrossRef]
- Morris, J.T.; Dacey, J.W.H. Effect of oxygen on ammonium uptake and root respiration by Spartina alternifolia. Am. J. Bot. 1984, 71, 979–985. [Google Scholar] [CrossRef]
- Weisner, S.E.; Granéli, W. Influence of substrate conditions on the growth of Phragmites australis after a reduction in oxygen transport to below-ground parts. Aquat. Bot. 1989, 35, 71–80. [Google Scholar] [CrossRef]
- Armstrong, J.; Armstrong, W.; Van der Putten, W.H. Phragmites die-back: Bud and root death, blockages within the aeration and vascular systems and the possible role of phytotoxins. New Phytol. 1996, 133, 399–414. [Google Scholar] [CrossRef]
- Brix, H.; Sorrel, B.K. Oxygen stress in wetland plants: Comparison of de-oxygenated and reducing root environments. Funct. Ecol. 1996, 10, 521–526. [Google Scholar] [CrossRef]
- Fuchs, C. The beetle Donacia clavipes as possible cause for the reed decline at lake Constance (Untersee). Limnol. Aktuell 1993, 5, 41–48. [Google Scholar]
- Tscharntke, T. Insects on common reed (Phragmites australis): Community structure and the impact of herbivory on shoot growth. Aquat. Bot. 1999, 64, 399–410. [Google Scholar] [CrossRef]
- Boorman, L.A.; Fuller, R.M. The changing status of reedswamp in the Norfolk Broads. J. Appl. Ecol. 1981, 18, 241–269. [Google Scholar] [CrossRef]
- Skuhravý, V.; Pokorný, V.; Pelikán, J.; Skuhravá, M.; Hudec, K.; Rychnovský, B. Invertebrates and Vertebrates Attacking Common Reed Stands (Phragmites communis) in Czechoslovakia; Studie ČSAV; Academia: Praha, Czech Republic, 1981; Volume 1, pp. 1–112. [Google Scholar]
- Bertolino, S.; Perrone, A.; Gola, L. Effectiveness of coypu control in small Italian wetland areas. Wildl. Soc. Bull. 2005, 33, 714–720. [Google Scholar] [CrossRef]
- van der Hut, R.M.G. Habitat choice and temporal differentiation in reed passerines of a Dutch marsh. Ardea 1986, 74, 159–176. [Google Scholar]
- Leisler, B.; Ley, H.W.; Winkler, H. Habitat, behavior and morphology of Acrocephalus warblers: An integrated analysis. Ornis Scand. 1989, 20, 181–186. [Google Scholar] [CrossRef]
- Jedraszko-Dabrowska, D. Reeds as construction supporting great reed warbler (Acrocephalus arundinaceus L.) and reed warbler (Acrocephalus scirpaceus Herm.) nests. Ekol. Pol. 1992, 39, 229–242. [Google Scholar]
- Ward, D. Reedbeds for Wildlife; The Royal Society for the Protection of Birds: Oxford, UK; University of Bristol Information Press: Oxford, UK, 1992. [Google Scholar]
- Poulin, B.; Davranche, A.; Lefebvre, G. Ecological assessment of Phragmites australis wetlands using multi-season SPOT-5 scenes. Remote Sens. Environ. 2010, 114, 1602–1609. [Google Scholar] [CrossRef]
- Schiess, H. Schilfbestände als Habitatinselm von Vögeln. Vierteljahrsschr. Der Nat. Ges. Zuerich 1990, 135, 259–265. [Google Scholar]
- Tscharntke, T. Fragmentation of Phragmites habitats, minimum viable population size, habitat suitability, and local extinction of moths, midges, flies, aphids, and birds. Conserv. Biol. 1992, 6, 530–536. [Google Scholar] [CrossRef]
- Poulin, B.; Lefebvre, G.; Mathevet, R. Habitat selection by booming bitterns Botaurus stellaris in French Mediterranean reed-beds. Oryx 2005, 39, 265–274. [Google Scholar] [CrossRef]
- Poulin, B.; Lefebvre, G.; Mauchamp, A. Habitat requirements of passerines and reedbed management in southern France. Biol. Conserv. 2002, 107, 315–325. [Google Scholar] [CrossRef]
- Puglisi, L.; Adamo, M.C.; Baldaccini, N.E. Man-induced habitat changes and sensitive species: A GIS approach to the Eurasian Bittern (Botaurus stellaris) distribution in a Mediterranean wetland. Biodivers. Conserv. 2005, 14, 1909–1922. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Lefebvre, G.; Poulin, B.; Tscharntke, T. Reed cutting affects arthropod communities, potentially reducing food for passerine birds. Biol. Conserv. 2005, 121, 157–166. [Google Scholar] [CrossRef]
- Knights, B. Enhancing stocks of European eel, Anguilla anguilla, to benefit bittern, Botaurus stellaris. In Interactions between Fish and Birds: Implications for Management; Cowx, I.G., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2003; Chapter 22; pp. 298–313. [Google Scholar]
- Noble, R.A.A.; Harvey, J.P.; Cowx, I.G.G. Can management of freshwater fish populations be used to protect and enhance the conservation status of a rare, fish-eating bird, the bittern, Botaurus stellaris, in the UK? Fish. Manag. Ecol. 2004, 11, 291–302. [Google Scholar] [CrossRef]
- Kloskowski, J.; Krogulec, J. Habitat selection of Aquatic Warbler Acrocephalus paludicola in Poland: Consequences for conservation of the breeding areas. Vogelvelt 1999, 120, 113–120. [Google Scholar]
- Tanneberger, F.; Flade, M.; Preiksa, Z.; Schröder, B. Habitat selection of the globally threatened Aquatic Warbler Acrocephalus paludicola at the western margin of its breeding range and implications for management. Ibis 2010, 152, 347–358. [Google Scholar] [CrossRef]
- Dijkema, K.S. Salt and brackish marshes around the Baltic Sea and adjacent parts of the North Sea: Their vegetation and management. Biol. Conserv. 1990, 51, 191–210. [Google Scholar] [CrossRef]
- Hellings, S.E.; Gallagher, J.L. The effects of salinity and flooding on Phragmites australis. J. Appl. Ecol. 1992, 29, 41–49. [Google Scholar] [CrossRef]
- Haslam, S.M.; Klötzli, F.; Sukopp, H.; Szczepański, A. The management of wetlands. In The Production Ecology of Wetlands; Westlake, D.F., Květ, J., Szczepański, A., Eds.; Cambridge University Press: Cambridge, UK, 1998; pp. 405–464. [Google Scholar]
- Wichtmann, W.; Schröder, C.; Joosten, H. Paludiculture—Productive Use of Wet Peatlands. Climate Protection—Biodiversity—Regional Economic Benefits; Schweizerbart Science Publishers: Stuttgart, Germany, 2016. [Google Scholar]
- Ziegler, R.; Wichtmann, W.; Abel, S.; Kemp, R.; Simard, M.; Joosten, H. Wet peatland utilisation for climate protection–An international survey of paludiculture innovation. Clean. Eng. Technol. 2021, 5, 100305. [Google Scholar] [CrossRef]
- Lahtinen, L.; Mattila, T.; Myllyviita, T.; Seppälä, J.; Vasander, H. Effects of paludiculture products on reducing greenhouse gas emissions from agricultural peatlands. Ecol. Eng. 2022, 175, 106502. [Google Scholar] [CrossRef]
- Wichmann, S.; Köbbing, J.F. Common reed for thatching—A first review of the European market. Ind. Crops Prod. 2015, 77, 1063–1073. [Google Scholar] [CrossRef]
- Becker, L.; Wichmann, S.; Beckmann, V. Common Reed for Thatching in Northern Germany: Estimating the Market Potential of Reed of Regional Origin. Resources 2020, 9, 146. [Google Scholar] [CrossRef]
- Mesléard, F.; Perennou, C. Aquatic Emergent Vegetation, Ecology and Management; Conservation of Mediterranean Wetlands, No. 6; Tour du Valat: Arles, France, 1996. [Google Scholar]
- Wichtmann, W.; Wichmann, S. Environmental, social and economic aspects of a sustainable biomass production. J. Sustain. Energy Environ. Spec. Issue 2011, 1, 77–81. [Google Scholar]
- Dragoni, F.; Giannini, V.; Ragaglini, G.; Bonari, E.; Silvestri, N. Effect of harvest time and frequency on biomass quality and biomethane potential of common reed (Phragmites australis) under paludiculture conditions. Bioenergy Res. 2017, 10, 1066–1078. [Google Scholar] [CrossRef]
- Eller, F.; Ehde, P.M.; Oehmke, C.; Ren, L.; Brix, H.; Sorrell, B.K.; Weisner, S.E. Biomethane Yield from different European Phragmites australis genotypes, compared with other herbaceous wetland species grown at different fertilization regimes. Resources 2020, 9, 57. [Google Scholar] [CrossRef]
- Granéli, W. Reed (Phragmites australis (Cav.) Trin. ex Steudel) as an energy source in Sweden. Biomass 1984, 4, 183–208. [Google Scholar] [CrossRef]
- Kitzler, H.; Pfeifer, C.; Hofbauer, H. Combustion of reeds in a 3 MW district heating plant. Int. J. Environ. Sci. Dev. 2012, 3, 407–411. [Google Scholar] [CrossRef]
- Lica, D.; Coşereanu, C.; Budău, G.; Lunguleasa, A. Characteristics of reed briquettes--biomass renewable resource of the Danube delta. Pro Ligno 2012, 8, 44–51. [Google Scholar]
- Patuzzi, F.; Mimmo, T.; Cesco, S.; Gasparella, A.; Baratieri, M. Common reeds (Phragmites australis) as sustainable energy source: Experimental and modelling analysis of torrefaction and pyrolysis processes. GCB Bioenergy 2012, 5, 367–374. [Google Scholar] [CrossRef]
- Ren, L.; Eller, F.; Lambertini, C.; Guo, W.Y.; Brix, H.; Sorrell, B.K. Assessing nutrient responses and biomass quality for selection of appropriate paludiculture crops. Sci. Total Environ. 2019, 664, 1150–1161. [Google Scholar] [CrossRef]
- Kuptz, D.; Kuchler, C.; Rist, E.; Eickenscheidt, T.; Mack, R.; Schön, C.; Hartmann, H. Combustion behaviour slagging tendencies of pure blended kaolin additivated biomass pellets from fen paludicultures in two small-scale boilers < 30 kW. Biomass Bioenergy 2022, 164, 106532. [Google Scholar]
- Hartung, C.; Andrade, D.; Dandikas, V.; Eickenscheidt, T.; Drösler, M.; Zollfrank, C.; Heuwinkel, H. Suitability of paludiculture biomass as biogas substrate− biogas yield and long-term effects on anaerobic digestion. Renew. Energy 2020, 159, 64–71. [Google Scholar] [CrossRef]
- Wichtmann, W.; Tanneberg, F. Land use options for rewetted peatlands. In Carbon Credits from Peastland Rewetting. Climate–Biodiversity–Land Use; Tanneberg, F., Wichtmann, W., Eds.; Schweitzerbart Science Publishers: Stuttgart, Austria, 2011; pp. 107–132. [Google Scholar]
- Wichmann, S. Commercial viability of paludiculture: A comparison of harvesting reeds for biogas production, direct combustion, and thatching. Ecol. Eng. 2017, 103, 497–505. [Google Scholar] [CrossRef]
- Toorn, J. van der Variability of Phragmites australis (Cav.) Trin. Ex Steudel in relation to the environment. Van Zee Tot Land 1972, 48, 1–122. [Google Scholar]
- Mook, J.H.; van der Toorn, J.W. Experiment on the development of reed vegetation in the Zuidflevoland polder. In Progress Report, 1974; Institute of Ecological Research: Heteren, The Netherlands, 1974; pp. 11–15. [Google Scholar]
- Lutz, M. Les étangs de pisciculture en Europe centrale. Typologie des systèmes d’exploitation et impacts des modalités de gestion sur l’avifaune. Ph.D. Thesis, Université Louis Pasteur, Strasbourg, France, 2001. [Google Scholar]
- Dyrcz, A. Breeding ecology of the great Reed Warlber (Acrocephalus arundinaceus) and Reed Warbler (A. scirpaceus) at fish ponds in SW-Poland and lakes in NW-Switzerland. Acta Ornithol. Warszawa 1981, 18, 307–334. [Google Scholar]
- Broyer, J.; Varagnat, P.; Constant, G.; Caron, P. Habitat du Héron pourpré Ardea purpurea sur les étangs de pisciculture en France. Alauda 1998, 66, 221–228. [Google Scholar]
- Brochet, A.L.; Gauthier-Clerc, M.; Mathevet, R.; Béchet, A.; Mondain-Monval, J.Y.; Tamisier, A. Marsh management, reserve creation, hunting periods and carrying capacity for wintering ducks and coots. Biodivers. Conserv. 2009, 18, 1879–1894. [Google Scholar] [CrossRef]
- Prokešová, J.; Kocian, L. Habitat selection of two Acrocephalus warblers breeding in reed beds near Malacky (Western Slovakia). Biologia 2004, 59, 637–644. [Google Scholar]
- Grim, T.; Honza, M. Effect of habitat on the diet of reed warbler (Acrocephalus scirpaceus) nestlings. Folia Zool. 1996, 45, 31–34. [Google Scholar]
- Tamisier, A.; Grillas, P. A review of habitat changes in the Camargue: An assessment of the effects of the loss of biological diversity on the wintering waterfowl community. Biol. Conserv. 1994, 70, 39–47. [Google Scholar] [CrossRef]
- Wheeler, B.D. Integrating wildlife with commercial uses. In Reedbeds for Wildlife; Ward, D., Ed.; The Royal Society for the Protection of Birds: Oxford, UK; University of Bristol Information Press: Oxford, UK, 1992; pp. 79–89. [Google Scholar]
- van Deursen, E.J.M.; Drost, H.J. Defoliation and treading by cattle of reed Phragmites australis. J. Appl. Ecol. 1990, 27, 284–297. [Google Scholar] [CrossRef]
- Poulin, B.; Duborper, E.; Lefebvre, G. Spring stopover of the globally threatened aquatic warbler Acrocephalus paludicola in Mediterranean France. Ardeola 2010, 57, 167–173. [Google Scholar]
- Hejný, S.; Husák, Š. Effect of fishpond management on the littoral communities. Exploitation of reed. In Pond Littoral Ecosystems. Structure and Functioning; Dykyjová, D., Květ, J., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1978; pp. 397–415. [Google Scholar]
- Kubů, F.; Květ, J.; Hejný, S. Fishpond management (Czechoslovakia). In Wetlands and Shallow Continental Water Bodies; Patten, B.C., Jorgeusen, S.E., Dumont, H.J., Gopal, B., Koryavov, P., Kvet, J., Loftier, H., Sverizhev, Y., Tundisi, J.G., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1994; Volume 2, pp. 391–404. [Google Scholar]
- Bernard, C. L’étang, l’homme et l’oiseau Incidences des modes de gestion des étangs piscicoles sur les ceintures de végétation et l’avifaune nicheuse en Sologne, Brenne, Bresse, Territoire de Belfort et Champagne humide. Ph.D. Thesis, Ecole Normale Supérieure des Lettres et Sciences Humaines de Lyon, Lyon, France, 2018. [Google Scholar]
- Broyer, J.; Curtet, L. The influence of fish farming intensification on taxonomic richness and biomass density of macrophyte-dwelling invertebrates in French fishponds. Knowl. Manag. Aquat. Ecosyst. 2011, 400, 10. [Google Scholar] [CrossRef]
- Janda, J.; Květ, J. Measuring ecological change in Czechoslovak wetlands. In Waterfowl and Wetland Conservation in the 1990s: A Global Perspective, Proceedings of an IWRB Symposium, St Petersburg Beach, FL, USA, 12–19 November 1992; Moser, M., Prentice, R.C., van Vessen, J., Eds.; IWRB: Slimbridge, UK, 1993; pp. 77–82. [Google Scholar]
- Joosten, H.; Clarke, D. Wise Use of Mires and Peatlands; International Mire Conservation Group: Kiel, Germany; International Peat Society: Jyväskylä, Finland, 2002; Volume 304. [Google Scholar]
- Pfadenhauer, J.; Sliva, J.; Marzelli, M. Renaturierungvon Landwirtschaftlich Genutzten Niedermooren Undabgetorften Hochmooren; Schriftenreihe Bayerisches Landes-amt für Umweltschutz 148: Augsburg, Germany, 2000. [Google Scholar]
- Hennicke, F. Das Naturschutzgroßprojekt Peenetal-Landschaft. In Landschaftsökologische Moorkunde, 2nd ed.; Succow, M., Joosten, H., Eds.; Schweizerbart: Stuttgart, Germany, 2001; pp. 487–492. [Google Scholar]
- Timmermann, T.; Margóczi, K.; Takács, G.; Vegelin, K. Restoration of peat-forming vegetation by rewetting species-poor fen grasslands. Appl. Veg. Sci. 2006, 9, 241–250. [Google Scholar] [CrossRef]
- Toet, S.; Van Logtestijn, R.S.P.; Schreijer, M.; Kampf, R.; Verhoeven, J.T.A. The functioning of a wetland system used for polishing effluent from a sewage treatment plant. Ecol. Eng. 2005, 25, 101–124. [Google Scholar] [CrossRef]
- Meerburg, B.G.; Vereijken, P.H.; de Visser, W.; Verhagen, J.; Korevaar, H.; Querner, E.P.; de Blaeij, A.T.; van der Werf, A. Surface water sanitation and biomass production in a large constructed wetland in the Netherlands. Wetl. Ecol. Manag. 2010, 18, 463–470. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Pedrocchi, C.; Comín, F.A. Effects of wetland construction on water quality in a semi-arid catchment degraded by intensive agricultural use. Ecol. Eng. 2010, 36, 631–639. [Google Scholar] [CrossRef]
- Vymazal, J.; Brix, H.; Cooper, P.F.; Green, M.B.; Haberl, R. (Eds.) Constructed Wetlands for Wastewater Treatment in Europe; Backhuys Publishers: Leiden, The Netherlands, 1998. [Google Scholar]
- Brix, H. Do macrophytes play a role in constructed treatment wetlands? Water Sci. Technol. 1997, 35, 11–17. [Google Scholar] [CrossRef]
- Cooper, P. What can we learn from old wetlands? Lessons that have been learned and some that may have been forgotten over the past 20 years. Desalination 2009, 24, 1–26. [Google Scholar] [CrossRef]
- Picek, T.; Čížková, H.; Dušek, J. Greenhouse gas emissions from a constructed wetland—Plants as important sources of carbon. Ecol. Eng. 2007, 31, 98–106. [Google Scholar] [CrossRef]
- Zhai, X.; Piwpuan, N.; Arias, C.A.; Headley, T.; Brix, H. Can root exudates from emergent wetland plants fuel denitrification in subsurface flow constructed wetland systems? Ecol. Eng. 2013, 61, 555–563. [Google Scholar] [CrossRef]
- Brix, H.; Schierup, H.-H. Soil oxygenation in constructed reed beds: The role of macrophyte and soil-atmospfere interface oxygen transport. In Constructed Wetlands in Water Pollution Control; Cooper, P.F., Findlater, B.C., Eds.; Pergamon Press: Oxford, UK, 1990; pp. 53–66. [Google Scholar]
- Armstrong, W.; Armstrong, J.; Beckett, P.M. Measurement and modelling of oxygen release from roots of Phragmites australis. In Constructed Wetlands in Water Pollution Control; Cooper, P.F., Findlater, B.C., Eds.; Pergamon Press: Oxford, UK, 1990; pp. 41–51. [Google Scholar]
- Tyroller, L.; Rousseau, D.P.L.; Santa, S.; García, J. Application of the gas tracer method for measuring oxygen transfer rates in subsurface flow constructed wetlands. Water Res. 2010, 44, 4217–4225. [Google Scholar] [CrossRef]
- Cooper, P. A review of the design and performance of vertical-flow and hybrid reed bed treatment systems. Water Sci. Technol. 1999, 40, 1–9. [Google Scholar] [CrossRef]
- Cooper, P. The performance of vertical flow constructed wetland systems with special reference to the significance of oxygen transfer and hydraulic loading rates. Water Sci. Technol. 2005, 51, 81–90. [Google Scholar] [CrossRef]
- Vymazal, J.; Kröpfelová, L. A three-stage experimental constructed wetland for treatment of domestic sewage: First two years of operation. Ecol. Eng. 2011, 37, 90–98. [Google Scholar] [CrossRef]
- Borin, M.; Tocchetto, D. Five year water and nitrogen balance for a constructed surface flow wetland treating agricultural drainage waters. Sci. Total Environ. 2007, 380, 38–47. [Google Scholar] [CrossRef]
- García-Lledó, A.; Ruiz-Rueda, O.; Vilar-Sanz, A.; Sala, L.; Bañeras, L. Nitrogen removal efficiencies in a free water surface constructed wetland in relation to plant coverage. Ecol. Eng. 2011, 37, 678–684. [Google Scholar] [CrossRef]
- Cloern, J.E. Our evolving conceptual model of the coastal eutrophication problem. Mar. Ecol. Prog. Ser. 2001, 210, 223–253. [Google Scholar] [CrossRef]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Dykyjová, D. Nutrient uptake by littoral communities of helophytes. In Pond Littoral Ecosystems. Structure and Functioning; Dykyjová, D., Květ, J., Eds.; Ecological Studies 28; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1978; pp. 257–291. [Google Scholar]
- Tátrai, I.; Mátyás, K.; Korponai, J.; Paulovits, G.; Pomogyi, P. The role of the Kis-Balaton Water Protection System in the control of water quality of Lake Balaton. Ecol. Eng. 2000, 16, 73–78. [Google Scholar] [CrossRef]
- Bousquet, F.; Le Page, C. Multi-agent simulations and ecosystem management: A review. Ecol. Model. 2004, 176, 313–332. [Google Scholar] [CrossRef]
- Mathevet, R.; Bousquet, F. Résilience & Environnement, Penser les Changements Socio-Écologiques; Buchet-Chastel: Paris, France, 2014. [Google Scholar]
- Mathevet, R.; Mauchamp, A.; Lifran, R.; Poulin, B.; Lefebvre, G. ReedSim: Simulating ecological and economical dynamics of Mediterranean reedbeds. In Integrative Modelling of Biophysical, Social and Economic Systems for Resource Management Solution; Post, D., Ed.; Modelling and Simulation Society of Australia and New Zealand: Townsville, Australia, 2003; pp. 1007–1012. [Google Scholar]
- Poulin, B.; Mathevet, R.; Le Page, C.; Etienne, M.; Lefebvre, G.; Poulin, B.; Gigot, G.; Proréol, S.; Mauchamp, A. BUTORSTAR: A role-playing game for collective awareness of wise reedbed use. Simul. Gaming 2007, 38, 233–262. [Google Scholar]
- Čížková, H.; Květ, J.; Comín, F.A.; Laiho, R.; Pokorný, J.; Pithart, D. Actual state of European wetlands and their possible future in the context of global climate change. Aquat. Sci. 2013, 75, 3–26. [Google Scholar] [CrossRef]
- Lefebvre, G.; Redmond, L.; Germain, C.; Palazzi, E.; Terzago, S.; Willm, L.; Poulin, B. Predicting the vulnerability of seasonally-flooded wetlands to climate change across the Mediterranean Basin. Sci. Total Environ. 2019, 692, 546–555. [Google Scholar] [CrossRef]
- Convention on Wetlands. Available online: www.ramsar.org (accessed on 10 April 2023).
- Tanneberg, F.; Wichtmann, W. (Eds.) Carbon Credits from Peastland Rewetting. Climate–Biodiversity–Land Use; Schweitzerbart Science Publishers: Stuttgart, Austria, 2011. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub-Class | Alliance | No. of Associations | Occurrence of P. australis | No. of Relevés | ||
|---|---|---|---|---|---|---|
| Dominant | Constant | Present | ||||
| Phragmitetalia | Phragmition communis | 19 | 1 | 5 | 18 | 12,690 |
| Bolboschoenetalia | Scirpion maritimi | 7 | 1 | 7 | 7 | 1682 |
| Bolboschoeno maritimi-Schoenoplection tabernaemontani | 6 | 1 | 6 | 3 | 1796 | |
| Magnocaricetalia | Magnocaricion elatae | 17 | 0 | 9 | 17 | 4452 |
| Magnocaricion gracilis | 6 | 0 | 3 | 6 | 5181 | |
| Carici-Rumicion hydrolapathi | 3 | 0 | 3 | 2 | 983 | |
| Nasturtio-Glycerietalia | Glycerio-Sparganion | 9 | 0 | 0 | 0 | 3177 |
| Caricion broterianae | 3 | 0 | 0 | 0 | 367 | |
| Oenanthetalia and Arctophiletalia | Eleocharito palustris-Sagittarion sagittifoliae | 18 | 0 | 1 | 17 | 4956 |
| Alopecuro-Glycerion spicatae | 1 | 0 | 0 | 0 | 30 | |
| Arctophilion fulvae | 1 | 0 | 0 | 0 | 19 | |
| Total | 11 | 90 | 3 | 34 | 70 | 35,333 |
| Region/Country | Freshwater Reed Beds | Brackish Swamps | Tall-Herb Fens and Moist Meadows |
|---|---|---|---|
| N and NW Europe | |||
| Scandinavia [32] | Schoenoplecto-Phragmitetum | Bolboschoenetum maritimi | Magnocaricion |
| Great Britain [22,31] | Phragmites australis comm. | Halo-Scirpion Elymion pycnanthi Ammophilion arenariae | Phragmites australis-Peucedanum palustre comm. Phragmites australis-Eupatorium cannabinum comm. |
| Netherlands [33] | Typho-Phragmitetum | Phragmition | In more communities |
| Central Europe | |||
| Germany [34] | Scirpo-Phragmitetum Phragmiti-Euphorbietum palustris | In more communities | Thelypterido-Phragmitetum Phragmiti-Caricetum lasiocarpae |
| Poland [35] | Phragmitetum australis | Phragmition | Thelypteridi-Phragmitetum |
| Czech Republic [36] | Phragmitetum australis Phragmition australis | Astero pannonici-Bolboschoenetum compacti Schoenoplectetum tabernaemontani | Thelypterido palustris-Phragmitetum australis Magno-Caricion elatae Cladietum marisci |
| Austria [37] | Phragmitetum vulgaris Phragmiti-Euphorbietum palustris | Bolboschoeno-Phragmitetum communis (inland salt marshes) | Caricion lasiocarpae |
| SE Europe | |||
| Hungary [38,39] | Phragmitetum communis Scirpo-Phragmitetum | – | – |
| Romania [20,40] | Scirpo-Phragmitetum | Phragmition | – |
| Croatia [41] | Phragmition | Caricetum vesicariae Phalaridetum arundinaceae | |
| E Europe | |||
| Ukraine [42] | Phragmitetum communis | Phragmiti-Juncetum maritimi | Phragmiteto-Schoenetum ferrugunei [43] |
| Russia [44,45,46] | Phragmition communis | Puccinellio-Phragmition | Phragmiti-Magnocaricion |
| (Volga [29]) | Calystegio-Phragmitetum | Argusio-Phragmitetum | – |
| S and SW Europe | |||
| France [47] | Phragmition (Scirpo-Phragmitetum) | Phragmites communis-Juncus maritimus-Scirpus maritimus comm. | – |
| Italy [48] | Phragmitetum australis | Bolboschoenus maritimus agg. community Schoenoplectetum tabernaemontani | Magno-Caricion elatae |
| Spain [47] | Typho angustifoliae-Phragmitetum australis Scirpo lacustris-Phragmitetum | Scirpo compacti-Phragmitetum australis | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čížková, H.; Kučera, T.; Poulin, B.; Květ, J. Ecological Basis of Ecosystem Services and Management of Wetlands Dominated by Common Reed (Phragmites australis): European Perspective. Diversity 2023, 15, 629. https://doi.org/10.3390/d15050629
Čížková H, Kučera T, Poulin B, Květ J. Ecological Basis of Ecosystem Services and Management of Wetlands Dominated by Common Reed (Phragmites australis): European Perspective. Diversity. 2023; 15(5):629. https://doi.org/10.3390/d15050629
Chicago/Turabian StyleČížková, Hana, Tomáš Kučera, Brigitte Poulin, and Jan Květ. 2023. "Ecological Basis of Ecosystem Services and Management of Wetlands Dominated by Common Reed (Phragmites australis): European Perspective" Diversity 15, no. 5: 629. https://doi.org/10.3390/d15050629
APA StyleČížková, H., Kučera, T., Poulin, B., & Květ, J. (2023). Ecological Basis of Ecosystem Services and Management of Wetlands Dominated by Common Reed (Phragmites australis): European Perspective. Diversity, 15(5), 629. https://doi.org/10.3390/d15050629