


Plant Diversity in the Diet of Costa Rican Primates in Contrasting Habitats: A Meta-Analysis

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Region
2.2. Literature Review and Data Collection
2.3. Description of Field Data Collection Methods in the Literature Reviewed
2.4. Statistical Analysis
2.4.1. Plant α-Diversity of Diet
2.4.2. Plant β-Diversity in Diet
2.4.3. Drivers of the Plant Assemblage in the Diet
3. Results
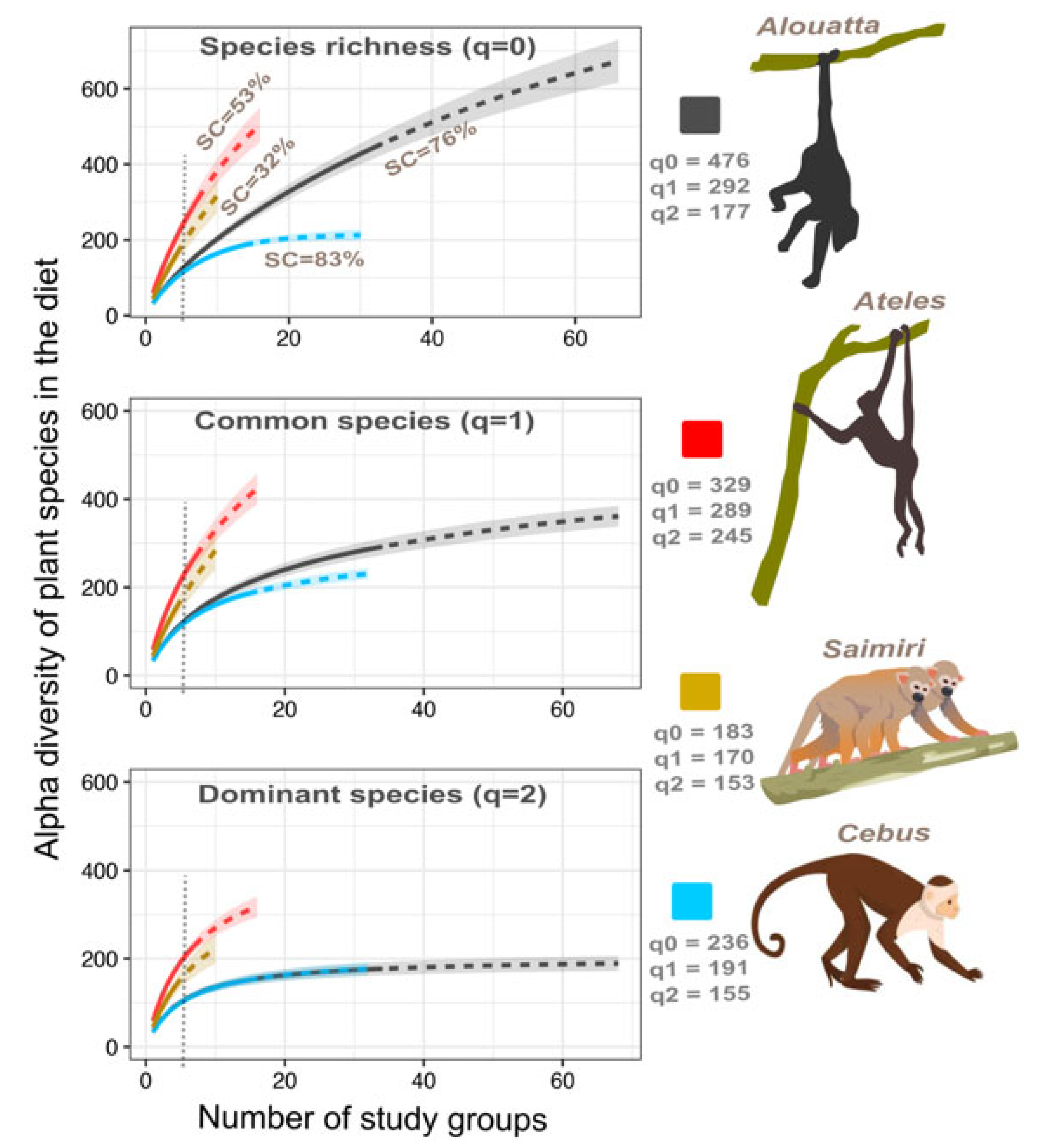
3.1. Alpha and Gamma Plant Species Diversity in the Diet of CR-NHP
3.2. Most Important Dietary Plant Species
3.3. Plant Species Assemblage Dissimilarity and β-Diversity Components
3.4. Factors Affecting Plant Assemblages in Diet
4. Discussion
4.1. Observed and Expected Diversity of Plants in Diet
4.2. Most Important Plant Species in Diet
4.3. Plant β-Diversity and Factors Affecting the Plant Composition in Diet
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kyriazakis, I.; Tolkamp, B.J.; Emmans, G. Diet Selection and Animal State: An Integrative Framework. Proc. Nutr. Soc. 1999, 58, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Jones, C. Behavioral Flexibility in Primates: Causes and Consequences; Springer: Berlin/Heidelberg, Germany, 2006; 136p, ISBN 0-387-23297-4. [Google Scholar]
- Ben-Dor, M.; Sirtoli, R.; Barkai, R. The Evolution of the Human Trophic Level during the Pleistocene. Am. J. Phys. Anthropol. 2021, 175, 27–56. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Rodríguez, V.; Dias, P.A.D. Effects of Habitat Fragmentation and Disturbance on Howler Monkeys: A review. Am. J. Primatol. 2010, 72, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Rodríguez, V.; Rös, M.; Escobar, F.; Melo, F.P.L.; Santos, B.A.; Tabarelli, M.; Chazdon, R. Plant β-Diversity in Fragmented Rain Forests: Testing Floristic Homogenization and Differentiation Hypotheses. J. Ecol. 2013, 101, 1449–1458. [Google Scholar] [CrossRef]
- Lowry, H.; Lill, A.; Wong, B.B. Behavioural Responses of Wildlife to Urban Environments. Biol. Rev. 2013, 88, 537–549. [Google Scholar] [CrossRef]
- Gruber, T.; Luncz, L.; Mörchen, J.; Schuppli, C.; Kendal, R.L.; Hockings, K. Cultural Change in Animals: A Flexible Behavioural Adaptation to Human Disturbance. Palgrave Commun. 2019, 5, 64. [Google Scholar] [CrossRef]
- Hindmarch, S.; Elliott, J.E. A Specialist in the City: The Diet of Barn Owls along a Rural to Urban Gradient. Urban Ecosyst. 2015, 18, 477–488. [Google Scholar] [CrossRef]
- Chaves, Ó.M.; Bicca-Marques, J.C. Crop Feeding by Brown Howlers (Alouatta guariba clamitans) in Forest Fragments: The Conservation Value of Cultivated Species. Int. J. Primatol. 2017, 38, 263–281. [Google Scholar] [CrossRef]
- Fuh, T.; Todd, A.; Feistner, A.; Donati, G.; Masi, S. Group Differences in Feeding and Diet Composition of Wild Western Gorillas. Sci. Rep. 2022, 12, 9569. [Google Scholar] [CrossRef]
- Smith, R.L.; Rebergen, K.; Payne, C.; Megapanos, E.; Lusseau, D. Dietary Plasticity of an Understudied Primate (Sapajus cay) in a Biodiversity Hotspot: Applying Ecological Traits to Habitat Conservation in the Upper Paraná Atlantic Forest. Folia Primatol. 2022, 93, 53–68. [Google Scholar] [CrossRef]
- Chaves, Ó.M.; Bicca-Marques, J.C. Dietary Flexibility of the Brown Howler Monkey Throughout Its Geographic Distribution. Am. J. Primatol. 2013, 75, 16–29. [Google Scholar] [PubMed]
- Bicca-Marques, J.C. How Do Howler Monkeys Cope with Habitat Fragmentation? In Primates in Fragments: Ecology and Conservation; Marsh, L.K., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003; pp. 283–303. [Google Scholar]
- Peres, C.A. Primate Responses to Phenological Changes in an Amazonian Terra Firme Forest. Biotropica 1994, 26, 98–112. [Google Scholar] [CrossRef]
- Chapman, C.A.; Bonnell, T.R.; Gogarten, J.F.; Lambert, J.E.; Omeja, P.A.; Twinomugisha, D.; Wasserman, M.D.; Rothman, J.M. Are Primates Ecosystem Engineers? Int. J. Primatol. 2013, 34, 1–14. [Google Scholar]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. Taxon. 1972, 21, 213–251. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity. Curr. Biol. 2021, 31, 1174–1177. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the Turnover and Nestedness Components of Beta Diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Ruhí, A.; Datry, T.; Sabo, J.L. Interpreting Beta-Diversity Components over Time to Conserve Metacommunities in Highly Dynamic Ecosystems. Conserv. Biol. 2017, 31, 1459–1468. [Google Scholar] [CrossRef]
- Milton, K. Distribution Patterns of Tropical Plant Foods as an Evolutionary Stimulus to Primate Mental Development. Am. Anthropol. 1981, 83, 534–548. [Google Scholar]
- Chapman, C.A. Foraging Strategies, Patch Use, and Constraints on Group Size in Three Species of Costa Rican Primates. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 1986. [Google Scholar]
- Chapman, C.A. Ecological Constraints on Group Size in Three Species of Neotropical Primates. Folia Primatol. 1990, 55, 1–9. [Google Scholar] [CrossRef]
- Milton, K. Food Choice and Digestive Strategies of Two Sympatric Primate Species. Am. Nat. 1981, 117, 496–505. [Google Scholar] [CrossRef]
- Di Fiore, A.; Link, A.; Dew, J.L. Diets of Wild Spider Monkeys. In Spider monkeys: Behavior, Ecology and Evolution of the Genus Ateles; Campbell, C.J., Ed.; Cambridge University Press: New York, NY, USA, 2008; pp. 81–137. [Google Scholar]
- Aureli, F.; Schaffner, C.M. Social Interactions, Social Relationships and the Social System of Spider Monkeys. In Spider Monkeys: Behavior, Ecology and Evolution of the Genus Ateles; Campbell, C.J., Ed.; Cambridge University Press: New York, NY, USA, 2008; pp. 236–265. [Google Scholar]
- Chaves, Ó.M.; Stoner, K.E.; Arroyo-Rodríguez, V. Differences in Diet Between Spider Monkey Groups Living in Forest Fragments and Continuous Forest in Mexico: Diet of Spider Monkeys in Lacandona. Biotropica 2012, 44, 105–113. [Google Scholar]
- Zaldívar, M.E.; Rocha, O.; Glander, K.E.; Aguilar, G.; Huertas, A.S.; Sánchez, R.; Wong, G. Distribution, Ecology, Life History, Genetic Variation, and Risk of Extinction of Nonhuman Primates from Costa Rica. Rev. Biol. Trop. 2004, 52, 679–693. [Google Scholar] [PubMed]
- Rylands, A.B.; Groves, C.P.; Mittermeier, R.A.; Cortés-Ortiz, L.; Hines, J.J.H. Taxonomy and Distributions of Mesoamerican Primates. In New Perspectives in the Study of Mesoamerican Primates: Distribution, Ecology, Behavior, and Conservation; Estrada, A., Garber, P.A., Pavelka, M.S.M., Luecke, L., Eds.; Springer: New York, NY, USA, 2006; pp. 29–79. [Google Scholar]
- Melin, A.D.; Hogan, J.D.; Campos, F.A.; Wikberg, E.; King-Bailey, G.; Webb, S.E.; Kalbitzer, U.; Asensio, N.; Murillo-Chacon, E.; Cheves Hernandez, S.; et al. Primate Life History, Social Dynamics, Ecology, and Conservation: Contributions from Long-Term Research in the Área de Conservación Guanacaste, Costa Rica. Biotropica 2020, 52, 1041–1064. [Google Scholar]
- Solano-Rojas, D. Importancia de los bosques secundarios para el mono tití centroamericano (Saimiri oerstedii oerstedii) en la Península de Osa, Costa Rica. In La Primatología en Latinoamérica. Tomo II Costa Rica-Venezuela; Urbani, B., Kowalewski, M., Cunha, R.G.T., de la Torre, S., Cortés-Ortiz, L., Eds.; Ediciones IVIC: Caracas, Spain, 2018; pp. 385–395. [Google Scholar]
- McKinney, T. Species-Specific Responses to Tourist Interactions by White-Faced Capuchins (Cebus imitator) and Mantled Howlers (Alouatta palliata) in a Costa Rican Wildlife Refuge. Int. J. Primatol. 2014, 35, 573–589. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species 2022. Available online: https://www.iucnredlist.org (accessed on 28 December 2022).
- SINAC. Resolución R-SINAC-CONAC-092-2017. Sistema Nacional de Áreas de Conservación. 2017. 13p. Available online: https://www.conagebio.go.cr/Conagebio/public/documentos/legislacion/Directrices/Resolucion92.pdf (accessed on 15 November 2022).
- Azofeifa-Rojas, I. Evaluación del Hábitat, Comportamiento y Riesgo de las Tropas de Monos Congo (Alouatta palliata). En Búsqueda de la Sostenibilidad con Fines Turísticos, en Playa Hermosa, Guanacaste. Master’s Thesis, Universidad de Costa Rica, San José, Costa Rica, 2022; 237p. [Google Scholar]
- Jones-Román, G.; Suárez, C.V.; Odio, R.M.M. Amenazas que Enfrentan los Monos Congo (Alouatta palliata) en Costa Rica e Iniciativas de Conservación Para el Bienestar y una Coexistencia Sana con la Especie. Biocenosis 2021, 32, 5–14. [Google Scholar]
- Hammel, B.; Grayum, M.; Herrera, C.; Zamora, N. Manual de Plantas de Costa Rica. Missouri Botanical Garden Press: St. Louis, MO, USA, 2004; Volume 1, 324p. [Google Scholar]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center Press: San Jose, Costa Rica, 1967; pp. 156–159. [Google Scholar]
- Pro, G.F.W.; Watcher, F.; Atlases, F. Monitoring Designed for Action Washington. Available online: https://www.wri.org/initiatives/global-forest-watch,2022, (accessed on 12 January 2023).
- Tosi, J.A. Mapa Ecológico, República de Costa Rica: Según la Clasificación de Zonas de Vida del Mundo de L.R. Holdridge; Tropical Science Center Press: San Jose, Costa Rica, 1969. [Google Scholar]
- Jiménez, Q.; Carrillo, E.; Kappelle, M. The Northern Pacific Lowland Seasonal Dry Forest of Guanacaste and the Nicoya Peninsula. In Costa Rican Ecosystems; Kappelle, M., Ed.; The University of Chicago Press: Chicago, CL, USA, 2015; pp. 247–289. [Google Scholar]
- Azofeifa-Rojas, I.; Sánchez-Porras, R.; Daniele, S. Mortalidad por Electrocución de Monos Congo (Alouatta palliata) Debido a Líneas Eléctricas en Guanacaste, Costa Rica. Mesoamericana 2021, 25, 15–21. [Google Scholar]
- Bolt, L.M.; Russell, D.G.; Schreier, A.L. Anthropogenic Edges Impact Howler Monkey (Alouatta palliata) Feeding Behaviour in a Costa Rican Rainforest. Primates 2021, 62, 647–657. [Google Scholar]
- Chapman, C. Flexibility in Diets of Three Species of Costa Rican Primates. Folia. Primatol. 1987, 49, 90–105. [Google Scholar]
- Chaves, Ó.M.; Morales-Cerdas, V.; Calderón-Quirós, J.; Retana, M. Data on the Diet Flexibility of Howler Monkeys (Alouatta palliata) in Contrasting Tropical Dry Forest Fragments. in Santa Cruz, Guanacaste, Costa Rica. 2023; unpublished data. [Google Scholar]
- Glander, K. Habitat Descriptions and Resource Utilization: A Preliminary Report on Mantled Howling Monkey Ecology. Socioecology and Psychology of Primates. In Socioecology and Psychology of Primates; Tuttle, R.H., Ed.; The Gruyter Mounton: Berlin, Germany, 1975; pp. 37–57. [Google Scholar]
- Glander, K.E. Howling Monkey Feeding Behavior and Plant Secondary Compounds: A Study of Strategies. In The Ecology of Arboreal Folivores; Montgomery, G.G., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1979; pp. 561–573. ISBN 0-19-513139-8. [Google Scholar]
- Larose, F. Foraging Strategies, Group Size, and Food Competition in the Mantled Howler Monkey, Alouatta palliata. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 1996; p. 168. [Google Scholar]
- Martínez, L. Comparación De Hábitos Alimentarios y su Relación con las Infecciones Parasíticas en los Monos Congo (Alouatta palliata) de Chomes y Palo Verde, Costa Rica. Undergraduate Thesis, Universidad de Costa Rica, San José, Costa Rica, 2010; p. 149. [Google Scholar]
- Mckinney, T. Diet, Ranging, and Activity Budget of White-Faced Capuchins (Cebus capucinus) in an Anthropogenic Habitat. Am. J. Phys. Anthropol. 2010, 167–168. [Google Scholar]
- Melin, A.D.; Khetpal, V.; Matsushita, Y.; Zhou, K.; Campos, F.A.; Welker, B.; Kawamura, S. Howler Monkey Foraging Ecology Suggests Convergent Evolution of Routine Trichromacy as an Adaptation for Folivory. Ecol. Evol. 2017, 7, 1421–1434. [Google Scholar] [CrossRef]
- Morera, R. Uso de Hábitat y Plantas Importantes en La Alimentación de Los Monos Congos (Alouatta palliata) y Carablancas (Cebus capucinus) en El Bosque Tropical Seco, Costa Rica. Ph.D. Thesis, Universidad Nacional, Heredia, Costa Rica, 1996; p. 144. [Google Scholar]
- Riba-Hernández, P.; Stoner, K.E. Data on Diet of Howler Monkeys in Punta Rio Claro, Costa Rica. 2002; unpublished data. [Google Scholar]
- Rockwood, L.L.; Glander, K.E. Howling Monkeys and Leaf-Cutting Ants: Comparative Foraging in a Tropical Deciduous Forest. Biotropica 1979, 11, 1–10. [Google Scholar] [CrossRef]
- Sánchez-Porras, R. Utilización Del Hábitat, Comportamiento Y Dieta Del Mono Congo (Alouatta palliata) en Un Bosque Premontano Húmedo, Costa Rica. Master’s Tesis, Universidad de Costa Rica, San José, Costa Rica, 1991; p. 123. [Google Scholar]
- Stoner, K.E.; Riba-Hernández, P. Data on Diet of Howler Monkeys in Sector Murciélago, Guanacaste, Costa Rica. 2002; unpublished data. [Google Scholar]
- Stoner, K.E. Habitat Selection and Seasonal Patterns of Activity and Foraging of Mantled Howling Monkeys (Alouatta palliata) in Northeastern Costa Rica. Int. J. Primatol. 1996, 17, 1–30. [Google Scholar] [CrossRef]
- Wehncke, E.V.; Valdez, C.N.; Dominguez, C.A. Seed Dispersal and Defecation Patterns of Cebus capucinus and Alouatta palliata: Consequences for Seed Dispersal Effectiveness. J. Trop. Ecol. 2004, 20, 535–543. [Google Scholar] [CrossRef]
- Perdomo, L. Aspectos Demográficos, Dieta, Anestesia y Evaluación Clínica de Monos Congos (Alouatta palliata) en el Parque Nacional Cahuita, Costa Rica. Master’s Thesis, Universidad Nacional: Heredia, Costa Rica, 2004; p. 100. [Google Scholar]
- Hiramatsu, C.; Melin, A.D.; Aureli, F.; Schaffner, C.M.; Vorobyev, M.; Matsumoto, Y.; Kawamura, S. Importance of Achromatic Contrast in Short-Range Fruit Foraging of Primates. PLoS ONE 2008, 3, e3356. [Google Scholar]
- Riba-Hernández, P. Efecto de la Vista a Color Polimórfica y los Carbohidratos Solubles en la Selección de Frutos por el Mono Araña (Ateles geoffroyi). Master’s Thesis, Universidad de Costa Rica, San José, Costa Rica, 2003. [Google Scholar]
- Riba-Hernández, P.; Stoner, K.E.; Lucas, P.W. The Sugar Composition of Fruits in the Diet of Spider Monkeys (Ateles geoffroyi) in Tropical Humid Forest in Costa Rica. J. Trop. Ecol. 2003, 19, 709–716. [Google Scholar] [CrossRef]
- Weghorst, J.A. Behavioral Ecology and Fission-Fusion Dynamics of Spider Monkeys (Ateles geoffroyi) in Lowland, Wet Forest. Ph.D. Thesis, Washington University, St. Louis, MO, USA, 2007. [Google Scholar]
- Whitworth, A.; Whittaker, L.; Pillco Huarcaya, R.; Flatt, E.; Morales, M.L.; Connor, D.; Priego, M.G.; Forsyth, A.; Beirne, C. Spider Monkeys Rule the Roost: Ateline Sleeping Sites Influence Rainforest Heterogeneity. Animals 2019, 9, 1052. [Google Scholar] [CrossRef]
- Chapman, C.A.; Fedigan, L.M. Dietary Differences between Neighboring Cebus capucinus Groups: Local Traditions, Food Availability or Responses to Food Profitability. Folia Primatol. 1990, 54, 177–186. [Google Scholar] [CrossRef]
- Chapman, C. Patterns of Foraging and Range Use by Three Species of Neotropical Primates. Primates 1988, 29, 177–194. [Google Scholar] [CrossRef]
- Chapman, C.A. Primate Seed Dispersal: The Fate of Dispersed Seeds. Biotropica 1989, 21, 148–154. [Google Scholar] [CrossRef]
- Hogan, J.D.; Melin, A.D.; Mosdossy, K.N.; Fedigan, L.M. Seasonal Importance of Flowers to Costa Rican Capuchins (Cebus capucinus imitator): Implications for Plant and Primate. Am. J. Phys. Anthropol. 2016, 161, 591–602. [Google Scholar] [CrossRef]
- McCabe, G.M. Diet and Nutrition in White-Faced Capuchins (Cebus capucinus): Effects of Group, Sex and Reproductive State. Master’s Thesis, University of Calgary, Calgary, AB, Canada, 2005; p. 120. [Google Scholar]
- McKinney, T. The Effects of Provisioning and Crop-Raiding on the Diet and Foraging Activities of Human-Commensal White-Faced Capuchins (Cebus capucinus). Am. J. Primatol. 2011, 73, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Melin, A.D.; Hiramatsu, C.; Parr, N.A.; Matsushita, Y.; Kawamura, S.; Fedigan, L.M. The Behavioral Ecology of Color Vision: Considering Fruit Conspicuity, Detection Distance and Dietary Importance. Int. J. Primatol. 2014, 35, 258–287. [Google Scholar] [CrossRef]
- Mosdossy, K.N.; Melin, A.D.; Fedigan, L.M. Quantifying Seasonal Fallback on Invertebrates, Pith, and Bromeliad Leaves by White-Faced Capuchin Monkeys (Cebus capucinus) in a Tropical Dry Forest. Am. J. Phys. Anthropol. 2015, 158, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Mallott, E.K.; Garber, P.A.; Malhi, R.S. TrnL Outperforms RbcL as a DNA Metabarcoding Marker When Compared with the Observed Plant Component of the Diet of Wild White-Faced Capuchins (Cebus capucinus). PLoS ONE 2018, 13, e0199556. [Google Scholar] [CrossRef] [PubMed]
- Boinski, S. The Ecology of Squirrel Monkeys in Costa Rica. Ph.D. Thesis, The University of Texas, Austin, TX, USA, 1986; p. 232. [Google Scholar]
- Solano-Rojas, D. Data on The Feeding Behavior of Saimiri oerstedii oerstedii. 2019; unpublished data. [Google Scholar]
- Wong, G. Uso del Hábitat, Estimación de la Composición y Densidad Poblacional del Mono Tití (Saimiri oerstedii citrinellus) en la Zona de Manuel Antonio, Quepos, Costa Rica. Master’s Thesis, Universidad Nacional, Heredia, Costa Rica, 1990. [Google Scholar]
- Chaves, Ó.M.; Morales-Cerdas, V.; Calderón-Quirós, J.; Rojas-Azofeifa, I.; Riba-Hernández, P.; Solano-Rojas, D.; Chacón-Madrigal, E.; Chaves, C.; Melin, A.D. Data and R-Scripts from: Plant Diversity in Diet of Costa Rican Primates in Contrasting Habitats: A Meta-Analysis. Figshare Dataset. 2023. Available online: https://doi.org/10.6084/m9.figshare.21785588.v1 (accessed on 15 January 2022).
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef]
- Jost, L. Partitioning Diversity into Independent Alpha and Beta Components. Ecology. 2007, 88, 2427–2439. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R Package for Rarefaction and Extrapolation of Species Diversity (Hill Numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Estimating Species Richness. In Biological Diversity; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: New York, NY, USA, 2011; pp. 39–50. [Google Scholar]
- Gotelli, N.J.; Colwell, R.K. Quantifying Biodiversity: Procedures and Pitfalls in the Measurement and Comparison of Species Richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-Based Rarefaction and Extrapolation: Standardizing Samples by Completeness Rather than Size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis; Pearson Prentice-Hall: New York, NY, USA, 2010. [Google Scholar]
- Baselga, A.; Orme, C.D.L. Betapart: An R Package for the Study of Beta Diversity. Methods Ecol. Evol. 2012, 3, 808812. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; López, M. Betapart: Partitioning Beta Diversity into Turnover and Nestedness Components, Version 1.5.6; CRAN R: Vienna, Austria, 2022. [Google Scholar]
- Staudhammer, C.L.; Escobedo, F.J.; Blood, A. Assessing Methods for Comparing Species Diversity from Disparate Data Sources: The Case of Urban and Peri-urban Forests. Ecosphere 2018, 9, 1–21. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Dias, P.A.; Rangel-Negrín, A. Diets of Howler Monkeys. In Howler Monkeys, Developments in Primatology: Progress and Prospects; Kowalewski, M.M., et al., Eds.; Springer: New York, NY, USA, 2015; pp. 21–56. [Google Scholar]
- De Freitas, C.H.; Setz, E.Z.F.; Araujo, A.R.B.; Gobbi, N. Agricultural Crops in the Diet of Bearded Capuchin Monkeys, Cebus libidinosus Spix (Primates: Cebidae), in Forest Fragments in Southeast Brazil. Rev. Bras. Zool. 2008, 25, 32–39. [Google Scholar] [CrossRef]
- Gómez-Posada, C. Diet and Feeding Behavior of a Group of Brown Capuchin Monkeys Cebus apella According to Fruits and Arthropods Availability, in the Colombian Amazon. Acta Amaz. 2012, 42, 363–372. [Google Scholar] [CrossRef]
- Lima, E.M.; Ferrari, S.F. Diet of a Free-Ranging Group of Squirrel Monkeys (Saimiri sciureus) in Eastern Brazilian Amazonia. Folia Primatol. 2003, 74, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Paim, F.P.; Chapman, C.A.; de Queiroz, H.L.; Paglia, A.P. Does Resource Availability Affect the Diet and Behavior of the Vulnerable Squirrel Monkey, Saimiri vanzolinii. Int. J. Primatol. 2017, 38, 572–587. [Google Scholar] [CrossRef]
- De Gonçalves, B.A.; Lima, L.C.P.; Aguiar, L.M. Diet Diversity and Seasonality of Robust Capuchins (Sapajus sp.) in a Tiny Urban Forest. Am. J. Primatol. 2022, 84, e23396. [Google Scholar]
- Lambert, J.E. Primate Nutritional Ecology: Feeding Biology and Diet at Ecological and Evolutionary Scales Oxford University Press. In Primates in Perspective, Campbell, C.J., Fuentes, A., Mackinnon, K.C., Panger, M., Bearder, S.J., Eds.; Oxford University Press: Oxford, UK, 2007; pp. 482–495. [Google Scholar]
- Felton, A.M.; Felton, A.; Lindenmayer, D.B.; Foley, W.J. Nutritional Goals of Wild Primates. Funct. Ecol. 2009, 23, 70–78. [Google Scholar] [CrossRef]
- Iason, G. The Role of Plant Secondary Metabolites in Mammalian Herbivory: Ecological Perspectives. Proc. Nutr. Soc. 2005, 64, 123–131. [Google Scholar]
- Freeland, W.J.; Janzen, D.H. Strategies in Herbivory by Mammals: The Role of Plant Secondary Compounds. Am. Nat. 1974, 108, 269–289. [Google Scholar]
- Coley, P.D.; Barone, J.A. Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Syst. 1996, 27, 305–335. [Google Scholar] [CrossRef]
- Windley, H.R.; Starrs, D.; Stalenberg, E.; Rothman, J.M.; Ganzhorn, J.U.; Foley, W.J. Plant Secondary Metabolites and Primate Food Choices: A Meta-Analysis and Future Directions. Am. J. Primatol. 2022, 84, e23397. [Google Scholar] [CrossRef] [PubMed]
- Janzen, D.H.; Waterman, P.G. A Seasonal Census of Phenolics, Fiber and Alkaloids in Foliage of Forest Trees in Costa Rica: Some Factors Influencing Their Distribution and Relation to Host Selection by Sphingidae and Saturniidae. Biol. J. Linn. Soc. 1984, 21, 439–454. [Google Scholar] [CrossRef]
- Kohl, K.D.; Weiss, R.B.; Cox, J.; Dale, C.; Dearing, M.D. Gut Microbes of Mammalian Herbivores Facilitate Intake of Plant Toxins. Ecol. Lett. 2014, 17, 1238–1246. [Google Scholar] [CrossRef]
- Lambert, J.E.; Rothman, J.M. Fallback Foods, Optimal Diets, and Nutritional Targets: Primate Responses to Varying Food Availability and Quality. Annu. Rev. Anthropol. 2015, 44, 493–512. [Google Scholar] [CrossRef]
- Narvaez-Rivera, G.M. The Human-Alloprimate Interface in Gandoca, Costa Rica: An Ethnoprimatological Approach to Assess Conflict between Residents and Three Neotropical Primates. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2017; p. 65. [Google Scholar]
- González-Zamora, A.; Arroyo-Rodríguez, V.; Chaves, Ó.M.; Sánchez-López, S.; Stoner, K.E.; Riba-Hernández, P. Diet of Spider Monkeys Ateles geoffroyi in Mesoamerica: Current Knowledge and Future Directions. Am. J. Primatol. 2009, 71, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Galán-Acedo, C.; Arroyo-Rodriguez, V.; Chapman, C.A. Beyond Patch Size: The Impact of Regional Context and Habitat Quality on Three Endangered Primates. Perspect. Ecol. Conserv. 2021, 19, 207–215. [Google Scholar] [CrossRef]
- Hopkins, M.E. Spatial Foraging Patterns and Ranging Behavior of Mantled Howler Monkeys Alouatta palliata, Barro Colorado Island, Panama. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2008; 130p. [Google Scholar]
- Solano-Rojas, D. Evaluación del hábitat, paisaje y la población del mono tití (Cebidae, Platyrrhini: Saimiri oerstedii oerstedii) en la península de Osa, Costa Rica. Master’s Thesis, Universidad de Costa Rica, San José, Costa Rica, 2007; 87p. [Google Scholar]
- Milton, K. Macronutrient Patterns of 19 Species of Panamanian Fruits from Barro Colorado Island. Neotrop. Prim. 2008, 15, 1–7. [Google Scholar] [CrossRef]
- Azofeifa Rojas, I.; Gregory, T. Canopy Bridges: Preventing and Mitigating Anthropogenic Impacts on Mantled Howler Monkeys (Alouatta palliata palliata) in Costa Rica. Folia Primatol. 2022, 93, 383–395. [Google Scholar] [CrossRef]
- Frankel, J.S.; Mallott, E.K.; Hopper, L.M.; Ross, S.R.; Amato, K.R. The effect of captivity on the primate gut microbiome varies with host dietary niche. Am. J. Primatol. 2010, 81, e23061. [Google Scholar] [CrossRef]
- Condit, R.; Pitman, N.; Leigh, E.G.; Chave, J.; Terborgh, J.; Foster, R.B.; Núñez, P.; Aguilar, S.; Valencia, R.; Villa, G.; et al. Beta-Diversity in Tropical Forest Trees. Science 2002, 295, 666–669. [Google Scholar] [CrossRef]
- Estrada, A.; Garber, P.A.; Rylands, A.B.; Roos, C.; Fernandez-Duque, E.; Di Fiore, A.; Nekaris, K.A.-I.; Nijman, V.; Heymann, E.W.; Lambert, J.E. Impending Extinction Crisis of the World’s Primates: Why Primates Matter. Sci. Adv. 2017, 3, 1–16. [Google Scholar]
- Estrada, A.; Garber, P.A.; Mittermeier, R.A.; Wich, S.; Gouveia, S.; Dobrovolski, R.; Nekaris, K.A.-I.; Nijman, V.; Rylands, A.B.; Maisels, F. Primates in Peril: The Significance of Brazil, Madagascar, Indonesia and the Democratic Republic of the Congo for Global Primate Conservation. PeerJ 2018, 6, e4869. [Google Scholar] [PubMed]
- Morera-Beita, A.; Sánchez, D.; Wanek, W.; Hofhansl, F.; Werner, H.; Chacón-Madrigal, E.; Montero-Muñoz, J.L.; Silla, F. Beta Diversity and Oligarchic Dominance in the Tropical Forests of Southern Costa Rica. Biotropica 2019, 51, 117–128. [Google Scholar]
- Chaves, Ó.M.; Bicca-Marques, J.C. Feeding Strategies of Brown Howler Monkeys in Response to Variations in Food Availability. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos-Barnett, T.C.; Cavalcante, T.; Boyle, S.A.; Matte, A.L.; Bezerra, B.M.; de Oliveira, T.G.; Barnett, A.A. Pulp Fiction: Why Some Populations of Ripe-Fruit Specialists Ateles chamek and A. marginatus Prefer Insect-Infested Foods. Int. J. Primatol. 2022, 43, 1–25. [Google Scholar]
- Pickett, S.B.; Bergey, C.M.; Di Fiore, A. A Metagenomic Study of Primate Insect Diet Diversity. Am. J. Primatol. 2012, 74, 622–631. [Google Scholar] [PubMed]
- Nishikawa, M.; Ferrero, N.; Cheves, S.; Lopez, R.; Kawamura, S.; Fedigan, L.M.; Melin, A.D.; Jack, K.M. Infant cannibalism in wild white-faced capuchin monkeys. Ecol. Evol. 2020, 23, 12679–12684. [Google Scholar]
- Fedigan, L.M. Vertebrate predation in Cebus capucinus: Meat eating in a neotropical monkey. Folia Primatol. 1990, 54, 196–205. [Google Scholar] [CrossRef]
- Boinski, S.; Tim, R.M. Predation by squirrel monkeys and double-toothed kites on tent-making bats. Am. J. Primatol. 1985, 9, 121–127. [Google Scholar] [CrossRef]
- Melin, A.D. Data on The Feeding Behavior of Saimiri oerstedii. 2015; unpublished data. [Google Scholar]
- Freese, C.H. Predation on swollen-thorn Acacia ants by white-faced monkeys, Cebus capucinus. Biotropica 1976, 8, 278–281. [Google Scholar] [CrossRef]
- Melin, A.D.; Fedigan, L.M.; Young, H.C.; Kawamura, S. Can color vision variation explain sex differences in invertebrate foraging by capuchin monkeys? Curr. Zool. 2010, 56, 300–312. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Species 2 | Sobs3 | Site 4 | HLZ 5 | Habitat 6 | Suc. 7 | PS 8 | #G 9 | GS 10 | SM 11 | SE 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 (T) | Ap | 37 | 1 (G) | TDF | mF | S | U | Pop | 13–24 | 1, 2 | 2 (9) |
| 2 (P) | Ap | 21 | 2 (L) | TLRF | C | S | P | 2 | 3/20 | 2 | 5 (10) |
| 3 (P) | Ap | 9 | 3 (G) | TDF | C | S | P | 40 | 40 | 1, 2 | 2 (17) |
| 4 (u) | Ap | 75 | 4, 5 (G) | TDF | sF, LF | S | P | 3 | 6/36/43 | 1, 2 | 2 (16) |
| 5 (b) | Ap | 47 | 6 (G) | TDF | sF | S | P | 1 | 13 | 2 | 2 (7) |
| 6 (b) | Ap | 61 | 6 (G) | TDF | C | S | P | 2 | 8/27 | 2 | 2 (14) |
| 7 (T*) | Ap | 57 | 3 (G) | TDF | C | S | P | 4 | 6/24 | 2 | 1 (11) |
| 8 (t) | Ap | 27 | 7 (G) | TDF | C | S | P | 1 | 38 | 1,2 | 1 (6) |
| 8 (t) | Ap | 22 | 8 (P) | TDF | sF | S | U | 1 | 8 | 1, 2 | 1 (6) |
| 9 (T*) | Ap | 52 | 9 (P) | TRF | C | S | P | 2 | 26/27 | 2 | 2 (24) |
| 10 (P) | Ap | 36 | 3 (G) | TDF | C | S | P | 2 | 10/7 | 3 | 3 (12) |
| 11 (T) | Ap | 40 | 3 (G) | TDF | C | M | P | 1 | 30 | 2 | 2 (12) |
| 12 (u) | Ap | 67 | 10 (P) | TRWF | LF | M | P | 2 | 20 | 2 | 2 (11) |
| 12 (u) | Ap | 62 | 10 (P) | TRF | LF | M | P | 1 | 18 | 1 | 2 (12) |
| 13 (P) | Ap | 22 | 6 (G) | TDF | sF | M | P | 1 | –– | 2 | 2 (10) |
| 14 (T) | Ap | 30 | 11 (A) | HPF | sF | S | U | 4 | 11 | 1, 2 | 2 (8) |
| 14 (T) | Ap | 12 | 12 (A) | HPF | sF | S | U | 4 | 15 | 1, 2 | 2 (7) |
| 15 (u) | Ap | 27 | 12 (G) | TDF | LF | S | P | 1 | –– | 2 | 3 (24) |
| 16 (P) | Ap | 64 | 13 (H) | TRF | C | M | P | 2 | 12/23 | 2 | 2 (15) |
| 16 (P) | Ap | 94 | 13 (H) | TRF | C | M | P | 2 | 12/23 | 2 | 2 (15) |
| 17 (P) | Ap | 9 | 7 (G) | TDF | C | S | P | 2 | 8/10 | –– | 2 (7) |
| 18 (P) | Ap | 72 | 3 (G) | TDF | C | S | P | Pop | –– | 1, 2 | 37 (–) |
| 19 (T) | Ap | 30 | 14 (L) | TRF | C | S | P | Pop | 2–29 | 4 | 2 (8) |
| 3 (P) | Ag | 12 | 3 (G) | TDF | C | S | P | 1 | 42 | 1, 2 | 3 (24) |
| 12 (u) | Ag | 18 | 12 (G) | TDF | LF | S | P | 1 | –– | 2 | 3 (24) |
| 18 (P) | Ag | 89 | 3 (G) | TDF | C | S | P | Pop | –– | 1, 2 | 37 (–) |
| 20 (P) | Ag | 33 | 3 (G) | TDF | C | M | P | 1 | 20 | 2 | 2 (8) |
| 21 (T) | Ag | 86 | 10 (P) | TRF | LF | M | P | 1 | 30 | 2 | 2 (12) |
| 22 (P) | Ag | 27 | 10 (P) | TRF | LF | M | P | 1 | 31 | 2 | 2 (12) |
| 23 (T*) | Ag | 98 | 15 (P) | TRF | C | M | P | 1 | 85 | 2, 4 | 2(17) |
| 24 (P) | Ag | 111 | 16 (P) | TRF | C | M | P | Pop | –– | 4, 5 | 2 (6) |
| 3 (P) | Ci | 6 | 3 (G) | TDF | C | S | P | 1 | 26 | 1, 2 | 2 (17) |
| 11 (T) | Ci | 56 | 3 (G) | TDF | C | M | P | 1 | 2 | 2 (12) | |
| 17 (P) | Ci | 30 | 7 (G) | TDF | C | S | P | 2 | 16/26 | –– | 2 (7) |
| 18 (P) | Ci | 142 | 3 (G) | TDF | C | S | P | Pop | –– | 1, 2 | 37 (–) |
| 25 (P) | Ci | 15 | 3 (G) | TDF | C | M, S | P | 3 | 26 | 2 | 3 (20) |
| 25 (P) | Ci | 13 | 3 (G) | TDF | C | M, S | P | 3 | 16 | 2 | 3 (20) |
| 25 (P) | Ci | 13 | 3 (G) | TDF | C | M, S | P | 3 | 26 | 2 | 3 (20) |
| 26 (P) | Ci | 5 | 3 (G) | TDF | C | M | P | 1 | 24 | 2 | 4 (24) |
| 27 (P) | Ci | 13 | 3 (G) | TDF | C | M | P | Pop | –– | 2 | 3 (26) |
| 28 (P) | Ci | 15 | 3 (G) | TDF | C | M, S | P | 6 | 8–35 | 1, 2, 7 | 8 (22) |
| 29 (T) | Ci | 35 | 3 (G) | TDF | C | M | P | 2 | 18–25 | 2 | 2 (7) |
| 30 (P) | Ci | 50 | 9 (P) | TRF | LF | S | P | 2 | 20/22 | 1, 2 | 2 (24) |
| 31 (P) | Ci | 112 | 3 (G) | TDF | C | M | P | 4 | –– | 1, 2 | 2 (12) |
| 32 (P) | Ci | 9 | 3 (G) | TDF | C | M | P | 4 | 72 | 2 | 1 (7) |
| 33 (P) | Ci | 59 | 2 (L) | TLRF | C | S | P | 1 | 22 | 2 | 1 (12) |
| 34 (T*) | So | 67 | 15 (P) | TRF | LF | M | P | 1 | 45 | 1, 2 | 2 (11) |
| 35 (b) | So | 25 | 17 (P) | TRF | C | S | P, U | 22 | 9–28 | 4, 6 | 2 (9) |
| 36 (u) | So | 92 | 18 (P) | TRF | LF | M | P, U | Pop | 28–99 | 2 | 2 (8) |
| 37 (T) | So | 33 | 19 (P) | TWF | LF | M | P, U | 14 | 22–66 | 2 | 2 (10) |
| Primate Species | Effort 1 | Observed Diversity (Hill Numbers) 2 | #Gen. | #Fam. | #E 3 | Expected Diversity (Hill Numbers) 4 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 0 | 1 | 2 | |||||
| Alouatta palliata | 34 (9000) | 476 (451–501) | 292 | 177 | 247 | 72 | 16 | 854 (748–959) | 431 (400–462) | 203 (186–220) |
| Ateles geoffroyi | 8 (5190) | 329 (306–352) | 289 | 245 | 172 | 60 | 4 | 739 (627–850) | 625 (553–696) | 450 (395–505) |
| Cebus imitator | 16 (5679) | 236 (221–251) | 191 | 155 | 170 | 67 | 17 | 302 (267–336) | 258 (242–274) | 206 (189–223) |
| Saimiri oerstedii | 5 (806) | 183 (161–205) | 170 | 153 | 96 | 48 | 10 | 763 (501–1024) | 645 (465–824) | 408 (322–494) |
| All | 61 (20,675) | 868 (829–907) * | 520 | 306 | 344 | 87 | 29 | 1740 (1561–1918) | 774 (726–823) | 348 (322–374) |
| Primate Species | Plant Species 1 | Family | GF 2 | Food Items 3 | Groups 4 |
|---|---|---|---|---|---|
| Alouatta palliata (Ap) | Samanea saman | Fabaceae | tree | L, l, lb, F, f, fl | 23 |
| Enterolobium cyclocarpum | Fabaceae | tree | L, l, F, f, fl | 21 | |
| Andira inermis | Fabaceae | tree | L, I, lb, fl | 19 | |
| Spondias mombin | Anacardiaceae | tree | L, l, lb, F, f, fl | 18 | |
| Bursera simaruba | Burseraceae | tree | L, l, lb, F, f | 17 | |
| Astronium graveolens | Anacardiaceae | tree | L, l, lb, F*, fl, p | 16 | |
| Brosimum alicastrum | Moraceae | tree | L, l, lb, F, f, fl | 16 | |
| Maclura tinctoria | Moraceae | tree | L, l, lb, F, f, fl | 16 | |
| Guazuma ulmifolia | Malvaceae | tree | L, l, F, f, | 14 | |
| Handroanthus ochraceus | Bignoniaceae | tree | L, l, fl | 14 | |
| Ateles geoffroyi (Ag) | Spondias mombin | Anacardiaceae | tree | F, fl | 7 |
| Dilodendron costaricense | Sapindaceae | tree | F* | 6 | |
| Brosimum alicastrum | Moraceae | tree | L*, F* | 5 | |
| B. costaricanum | Moraceae | tree | F, f | 4 | |
| Bursera simaruba | Burseraceae | tree | l, F* | 4 | |
| Cecropia peltata | Urticaceae | tree | L, l, F*, fl | 4 | |
| Garcinia madruno | Clusiaceae | tree | F | 4 | |
| Manilkara chicle | Sapotaceae | tree | f | 4 | |
| Pouteria torta | Sapotaceae | tree | l, F | 4 | |
| Sideroxylon capiri | Sapotaceae | tree | F, f | 4 | |
| Cebus imitator (Ci) | Aralia excelsa | Araliaceae | tree | L*, F* | 8 |
| Bursera simaruba | Burseraceae | tree | F* | 8 | |
| Muntingia calabura | Muntingiaceae | tree | F* | 8 | |
| Sloanea terniflora | Elaeocarpaceae | tree | F* | 8 | |
| Spondias mombin | Anacardiaceae | tree | F* | 8 | |
| Vachellia collinsii | Fabaceae | shrub | F* | 8 | |
| Genipa americana | Rubiaceae | tree | F* | 7 | |
| Luehea candida | Malvaceae | tree | F*, fl | 7 | |
| Luehea speciosa | Malvaceae | tree | F*, fl | 7 | |
| Ficus sp. SR | Moraceae | tree | L, l, F* | 7 | |
| Saimiri oerstedii (So) | Psidium guajava | Myrtaceae | tree | F, f | 4 |
| Anacardium excelsum | Anacardiaceae | tree | F* | 3 | |
| Cecropia obtusifolia | Urticaceae | tree | F* | 3 | |
| Conostegia schlimii | Melastomataceae | tree | F* | 3 | |
| Miconia argentea | Melastomataceae | tree | F* | 3 | |
| Nephelium lappaceum | Sapindaceae | tree | F, f | 3 | |
| Ochroma pyramidale | Malvaceae | tree | F*, n | 3 | |
| Passiflora vitifolia | Passifloraceae | vine | F*, n | 3 | |
| Symphonia globulifera | Clusiaceae | tree | F*, n | 3 | |
| Vitex cooperi | Lamiaceae | tree | F* | 3 | |
| All primate species | Spondias mombin | Anacardiaceae | tree | L, L*, l, F, f, fl, o | 35 |
| Bursera simaruba | Burseraceae | tree | L, L*, l, F*, fl, o | 29 | |
| Samanea saman | Fabaceae | tree | L, L*, l, F*,F, f, fl, s, o | 27 | |
| Brosimum alicastrum | Moraceae | tree | L, L*, l, F*,F, f, fl, o | 24 | |
| Enterolobium cyclocarpum | Fabaceae | tree | L, L*, l, F*,F, f, fl, s, o | 24 | |
| Maclura tinctoria | Moraceae | tree | L, L*, l, lb, F*,F, f, fl, o | 23 | |
| Andira inermis | Fabaceae | tree | L, L*, l, lb, fl, o | 22 | |
| Guazuma ulmifolia | Malvaceae | tree | L, L*, l, F, F*, f | 21 | |
| Muntingia calabura | Muntingiaceae | tree | L, L*, l, F, F*, fl | 21 | |
| Astronium graveolens | Anacardiaceae | tree | L, L*, l, F*, fl, p, o | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves, Ó.M.; Morales-Cerdas, V.; Calderón-Quirós, J.; Azofeifa-Rojas, I.; Riba-Hernández, P.; Solano-Rojas, D.; Chaves-Cordero, C.; Chacón-Madrigal, E.; Melin, A.D. Plant Diversity in the Diet of Costa Rican Primates in Contrasting Habitats: A Meta-Analysis. Diversity 2023, 15, 602. https://doi.org/10.3390/d15050602
Chaves ÓM, Morales-Cerdas V, Calderón-Quirós J, Azofeifa-Rojas I, Riba-Hernández P, Solano-Rojas D, Chaves-Cordero C, Chacón-Madrigal E, Melin AD. Plant Diversity in the Diet of Costa Rican Primates in Contrasting Habitats: A Meta-Analysis. Diversity. 2023; 15(5):602. https://doi.org/10.3390/d15050602
Chicago/Turabian StyleChaves, Óscar M., Vanessa Morales-Cerdas, Jazmín Calderón-Quirós, Inés Azofeifa-Rojas, Pablo Riba-Hernández, Daniela Solano-Rojas, Catalina Chaves-Cordero, Eduardo Chacón-Madrigal, and Amanda D. Melin. 2023. "Plant Diversity in the Diet of Costa Rican Primates in Contrasting Habitats: A Meta-Analysis" Diversity 15, no. 5: 602. https://doi.org/10.3390/d15050602
APA StyleChaves, Ó. M., Morales-Cerdas, V., Calderón-Quirós, J., Azofeifa-Rojas, I., Riba-Hernández, P., Solano-Rojas, D., Chaves-Cordero, C., Chacón-Madrigal, E., & Melin, A. D. (2023). Plant Diversity in the Diet of Costa Rican Primates in Contrasting Habitats: A Meta-Analysis. Diversity, 15(5), 602. https://doi.org/10.3390/d15050602