
Diversity of DNA Sequences from Pathogenic and Potentially Pathogenic Eukaryotic Microorganisms in Protected Granite Mountain Rocks

 , , , and
, , , and
Abstract

1. Introduction
2. Material and Methods
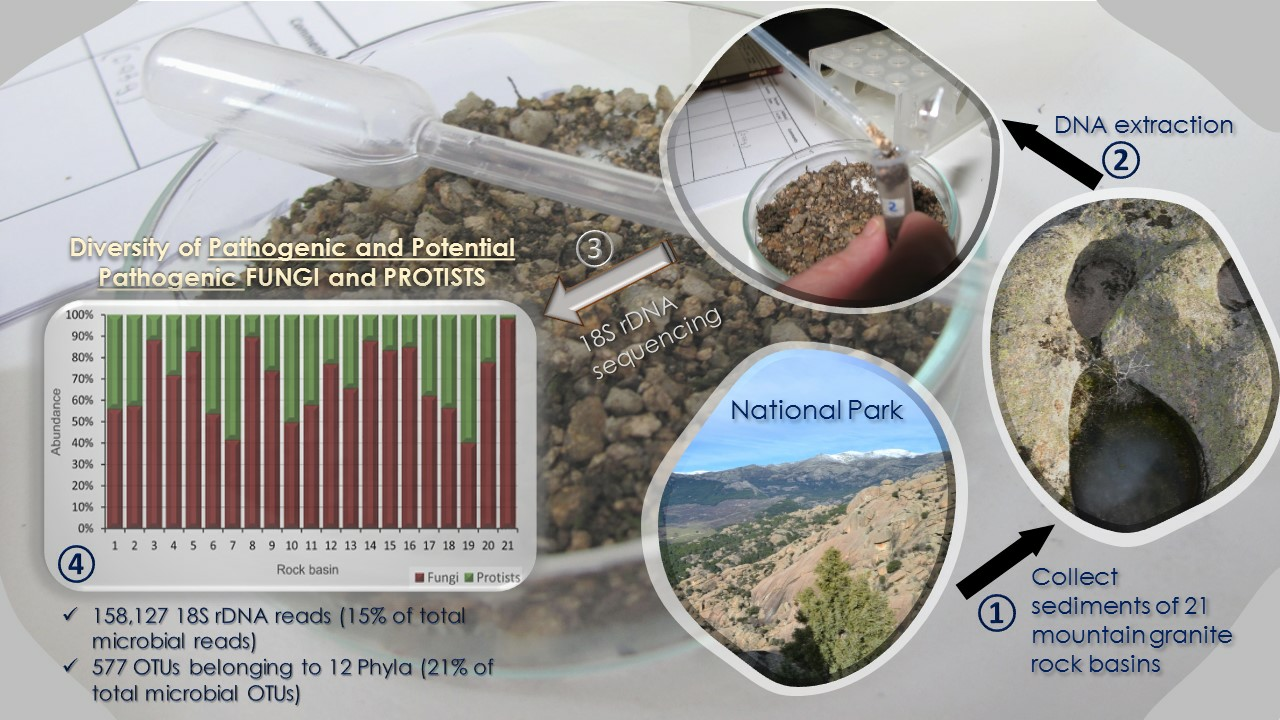
2.1. Habitats Studied and Sample Collection
2.2. Chemical Analyses
2.3. DNA Extraction and Sequencing
2.4. Bioinformatic Analyses
2.5. Taxonomic Classification of Pathogen and Potentially Pathogenic Microorganisms
2.6. Statistical Analyses
3. Results
3.1. Chemical Characterisation of Rock Basins
3.2. Community Structure of Pathogenic and Potentially Pathogenic OTUs
3.3. Taxonomic Affiliation of OTUs and Abundance and Richness of Phyla
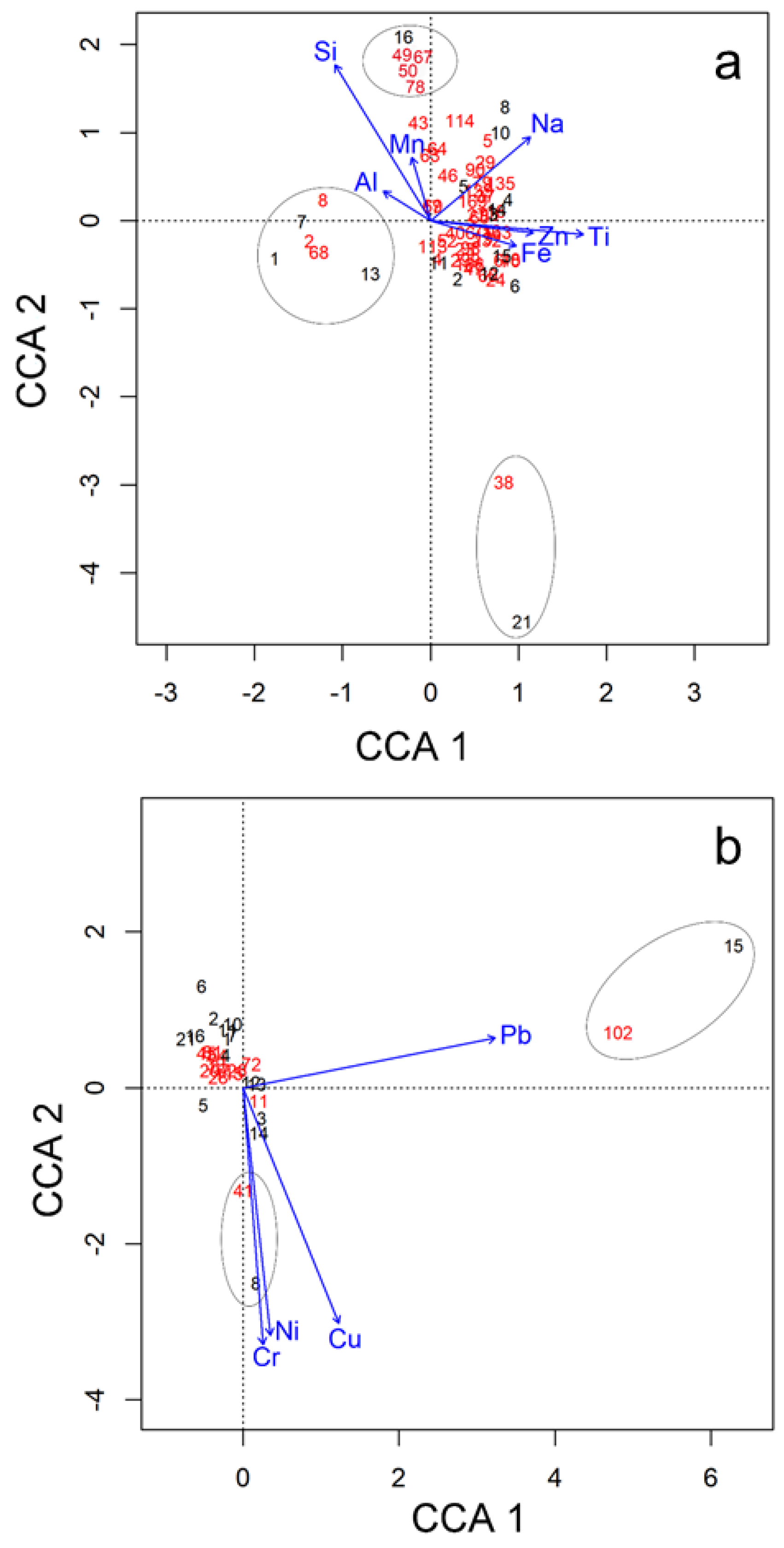
3.4. Relationship of OTUs with Basin Morphometry and Chemical Parameters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frenken, T.; Alacid, E.; Berger, S.A.; Bourne, E.C.; Gerphagnon, M.; Grossart, H.P.; Gsell, A.S.; Ibelings, B.W.; Kagami, M.; Küpper, F.C.; et al. Integrating chytrid fungal parasites into plankton ecology: Research gaps and needs. Environ. Microbiol. 2017, 19, 3802–3822. [Google Scholar] [CrossRef]
- Geisen, S.; Mitchell, E.A.D.; Adl, S.; Bonkowski, M.; Dunthorn, M.; Ekelund, F.; Fernández, L.D.; Jousset, A.; Krashevska, V.; Singer, D.; et al. Soil protists: A fertile frontier in soil biology research. FEMS Microbiol. Rev. 2018, 42, 293–323. [Google Scholar] [CrossRef] [PubMed]
- Selbach, C.; Soldánová, M.; Feld, C.K.; Kostadinova, A.; Sures, B. Hidden parasite diversity in a European freshwater system. Sci. Rep. 2020, 10, 2694. [Google Scholar] [CrossRef]
- Evison, S.E.; Fazio, G.; Chappell, P.; Foley, K.; Jensen, A.B.; Hughes, W.O. Host–parasite genotypic interactions in the honey bee: The dynamics of diversity. Ecol. Evol. 2013, 3, 2214–2222. [Google Scholar] [CrossRef]
- Hamilton, W.D. Pathogens as causes of genetic diversity in their host populations. In Population Biology of Infectious Diseases; Anderson, R.M., May, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; pp. 269–296. [Google Scholar]
- Ibelings, B.W.; De Bruin, A.; Kagami, M.; Rijkeboer, M.; Brehm, M.; Donk, E.V. Host parasite interactions between freshwater phytoplankton and chytrid fungi (Chytridiomycota). J. Phycol. 2004, 40, 437–453. [Google Scholar] [CrossRef]
- Mahé, F.; de Vargas, C.; Bass, D.; Czech, L.; Stamatakis, A.; Lara, E.; Singer, D.; Mayor, J.; Bunge, J.; Sernaker, S.; et al. Parasites dominate hyperdiverse soil protist communities in Neotropical rainforests. Nat. Ecol. Evol. 2017, 1, 0091. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Rassoulzadegan, F. Are viruses driving microbial diversification and diversity? Environ. Microbiol. 2004, 6, 1–11. [Google Scholar] [CrossRef]
- Amundsen, P.-A.; Lafferty, K.D.; Knudsen, R.; Primicerio, R.; Klemetsen, A.; Kuris, A.M. Food web topology and parasites in the pelagic zone of a subarctic lake. J. Anim. Ecol. 2009, 78, 563–572. [Google Scholar] [CrossRef]
- Sime-Ngando, T. Phytoplankton chytridiomycosis: Fungal parasites of phytoplankton and their imprints on the food web dynamics. Front. Microbiol. 2012, 3, 361. [Google Scholar] [CrossRef]
- Agha, R.; Saebelfeld, M.; Manthey, C.; Rohrlack, T.; Wolinska, J. Chytrid parasitism facilitates trophic transfer between bloom-forming cyanobacteria and zooplankton (Daphnia). Sci. Rep. 2016, 6, 35039. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Dobson, A.; Lafferty, K.D.; Marcogliese, D.J.; Memmott, J.; Orlofske, S.A.; Poulin, R.; Thieltges, D.W. When parasites become prey: Ecological and epidemiological significance of eating parasites. Trends Ecol. Evol. 2010, 25, 362–371. [Google Scholar] [CrossRef]
- De S Araújo, G.R.; de Souza, W.; Frases, S. The hidden pathogenic potential of environmental fungi. Fut. Microbiol. 2017, 12, 1533–1540. [Google Scholar] [CrossRef]
- Boisard, J.; Florent, I. Why the -omic future of Apicomplexa should include Gregarines. Biol. Cell 2020, 112, 173–185. [Google Scholar] [CrossRef]
- Comeau, A.M.; Vincent, W.F.; Bernier, L.; Lovejoy, C. Novel chytrid lineages dominate fungal sequences in diverse marine and freshwater habitats. Sci. Rep. 2016, 6, 30120. [Google Scholar] [CrossRef]
- Lefèvre, E.; Roussel, B.; Amblard, C.; Sime-Ngando, T. The molecular diversity of freshwater picoeukaryotes reveals high occurrence of putative parasitoids in the plankton. PLoS ONE 2008, 3, e2324. [Google Scholar] [CrossRef]
- de Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef]
- Morand, S. (macro-) Evolutionary ecology of parasite diversity: From determinants of parasite species richness to host diversification. Int. J. Parasitol. Parasites Wildl. 2015, 4, 80–87. [Google Scholar] [CrossRef]
- Corsaro, D.; Walochnik, J.; Venditti, D.; Hauröder, B.; Michel, R. Solving an old enigma: Morellospora saccamoebae gen. nov., sp. nov. (Rozellomycota), a Sphaerita-like parasite of free-living amoebae. Parasitol. Res. 2020, 119, 925–934. [Google Scholar] [CrossRef]
- Gleason, F.H.; Kagami, M.; Lefevre, E.; Sime-Ngando, T. The ecology of chytrids in aquatic ecosystems: Roles in food web dynamics. Fungal Biol. Rev. 2008, 22, 17–25. [Google Scholar] [CrossRef]
- Reynolds, H.; Raudabaugh, D.; Lilje, O.; Allender, M.; Miller, A.; Gleason, F. Emerging mycoses and fungus-like diseases of vertebrate wildlife. In The Fungal Community: Its Organization and Role in the Ecosystem, 4th ed.; Dighton, J., White, J.F., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 385–404. [Google Scholar]
- Lefranc, M.; Thénot, A.; Lepère, C.; Debroas, D. Genetic diversity of small eukaryotes in lakes differing by their trophic status. Appl. Environ. Microbiol. 2005, 71, 5935–5942. [Google Scholar] [CrossRef] [PubMed]
- López-García, P.; Rodríguez-Valera, F.; Pedrós-Alió, C.; Moreira, D. Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton. Nature 2001, 409, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Singer, D.; Lara, E.; Steciow, M.M.; Seppey, C.V.W.; Paredes, N.; Pillonel, A.; Oszako, T.; Belbahri, L. High-throughput sequencing reveals diverse oomycete communities in oligotrophic peat bog micro-habitat. Fungal Ecol. 2016, 23, 42–47. [Google Scholar] [CrossRef]
- Ortiz-Álvarez, R.; Triadó-Margarit, X.; Camarero, L.; Casamayor, E.O.; Catalan, J. High planktonic diversity in mountain lakes contains similar contributions of autotrophic, heterotrophic and parasitic eukaryotic life forms. Sci. Rep. 2018, 8, 4457. [Google Scholar] [CrossRef]
- Singer, D.; Duckert, C.; Heděnec, P.; Lara, E.; Hiltbrunner, E.; Mitchell, E.A.D. High-throughput sequencing of litter and moss eDNA reveals a positive correlation between the diversity of Apicomplexa and their invertebrate hosts across alpine habitats. Soil Biol. Biochem. 2020, 147, 107837. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the classification, nomenclature, and diversity of Eukaryotes. J. Eukaryotic Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed]
- Bråte, J.; Logares, R.; Berney, C.; Ree, D.K.; Klaveness, D.; Jakobsen, K.S.; Shalchian-Tabrizi, K. Freshwater Perkinsea and marine-freshwater colonizations revealed by pyrosequencing and phylogeny of environmental rDNA. ISME J. 2010, 4, 1144–1153. [Google Scholar] [CrossRef]
- Käse, L.; Metfies, K.; Neuhaus, S.; Boersma, M.; Wiltshire, K.H.; Kraberg, A.C. Host-parasitoid associations in marine planktonic time series: Can metabarcoding help reveal them? PLoS ONE 2021, 16, e0244817. [Google Scholar] [CrossRef]
- Kagami, M.; Miki, T.; Takimoto, G. Mycoloop: Chytrids in aquatic food webs. Front. Microbiol. 2014, 5, 166. [Google Scholar] [CrossRef] [PubMed]
- Schmeller, D.S.; Loyau, A.; Bao, K.; Brack, W.; Chatzinotas, A.; De Vleeschouwer, F.; Friesen, J.; Gandois, L.; Hansson, S.V.; Haver, M.; et al. People, pollution and pathogens—global change impacts in mountain freshwater ecosystems. Sci. Total Environ. 2018, 622, 756–763. [Google Scholar] [CrossRef]
- Jocque, M.; Vanschoenwinkel, B.; Brendonck, L. Freshwater rock pools: A review of habitat characteristics, faunal diversity and conservation value. Freshwater Biol. 2010, 55, 1587–1602. [Google Scholar] [CrossRef]
- Pérez Uz, M.B.; Velasco González, I.; Murciano, A.; Sánchez Jiménez, A.; García-Rodríguez, M.; Centeno Carrillo, J.d.D.; Montero González, E.; Muñoz Araújo, B.; Olmedo Salinas, C.; Quintela Alonso, P.; et al. Rain-fed granite rock pools in a national park: Extreme niches for protists. Limnetica 2021, 40, 1–18. [Google Scholar] [CrossRef]
- Birck, C.; Epaillard, I.; Leccia, F.; Crassous, C.; Morand, A.; Miaud, C.; Bertrand, C.; Cavalli, L.; Jacquet, S.; Moullec, P. Sentinel lakes: A network for the study and management of mountain lakes in the French Alps and in Corsica. Eco. Mont. 2013, 5, 63–69. [Google Scholar] [CrossRef]
- Oertli, B.; Biggs, J.; Céréghino, R.; Grillas, P.; Joly, P.; Lachavanne, J.-B. Conservation and monitoring of pond biodiversity: Introduction. Aquat. Conserv. Mar. Freshwater Ecosyst. 2005, 15, 535–540. [Google Scholar] [CrossRef]
- Velasco-González, I.; Sanchez-Jimenez, A.; Singer, D.; Murciano, A.; Díez-Hermano, S.; Lara, E.; Martín-Cereceda, M. Rain-fed granite rock basins accumulate a high diversity of dormant microbial eukaryotes. Microb. Ecol. 2020, 79, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Custer, G.F.; Bresciani, L.; Dini-Andreote, F. Ecological and evolutionary implications of microbial dispersal. Front. Microbiol. 2022, 13, 855859. [Google Scholar] [CrossRef]
- Baumgardner, D.J. Soil-related bacterial and fungal infections. J. Am. Board Family Med. 2012, 25, 734–744. [Google Scholar] [CrossRef]
- Rokas, A. Evolution of the human pathogenic lifestyle in fungi. Nat. Microbiol. 2022, 7, 607–619. [Google Scholar] [CrossRef]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.M.; Breiner, H.-W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef]
- Schiaffino, M.R.; Lara, E.; Fernández, L.D.; Balagué, V.; Singer, D.; Seppey, C.C.W.; Massana, R.; Izaguirre, I. Microbial eukaryote communities exhibit robust biogeographical patterns along a gradient of Patagonian and Antarctic lakes. Environ. Microbiol. 2016, 18, 5249–5264. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef]
- Pearson, W.R. BLAST and FASTA similarity searching for multiple sequence alignment. In Multiple Sequence Alignment Methods; Russell, D.J., Ed.; Humana Press: Totowa, NJ, USA, 2014; pp. 75–101. [Google Scholar]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference database (PR2): A catalog of unicellular eukaryote small sub-unit rRNA sequences with curated taxonomy. Nucleic Acids Res. 2012, 41, D597–D604. [Google Scholar] [CrossRef]
- Scheele, B.C.; Pasmans, F.; Skerratt, L.F.; Berger, L.; Martel, A.; Beukema, W.; Acevedo, A.A.; Burrowes, P.A.; Carvalho, T.; Catenazzi, A.; et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 2019, 363, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A. Separating the two components of abundance-based dissimilarity: Balanced changes in abundance vs. abundance gradients. Methods Ecol. Evol. 2013, 4, 552–557. [Google Scholar] [CrossRef]
- Gómez-Rodríguez, C.; Baselga, A. Variation among European beetle taxa in patterns of distance decay of similarity suggests a major role of dispersal processes. Ecography 2018, 41, 1825–1834. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ebert, D. Distributions and impacts of microparasites on Daphnia in a rockpool metapopulation. Oecologia 1998, 115, 213–221. [Google Scholar] [CrossRef]
- Therriault, T.W.; Kolasa, J. Desiccation frequency reduces species diversity and predictability of community structure in coastal rock pools. Israel J. Zool. 2001, 47, 477–489. [Google Scholar] [CrossRef]
- De Miguel, E. Determinación de Niveles de Fondo y Niveles de Referencia de Metales Pesados y Otros Elementos Traza en Suelos de la Comunidad de Madrid; Instituto Geológico y Minero de España: Madrid, Spain, 2002. [Google Scholar]
- Logares, R.; Mangot, J.-F.; Massana, R. Rarity in aquatic microbes: Placing protists on the map. Res. Microbiol. 2015, 166, 831–841. [Google Scholar] [CrossRef]
- Lepère, C.; Domaizon, I.; Debroas, D. Unexpected importance of potential parasites in the composition of the freshwater small-eukaryote community. Appl. Environ. Microbiol. 2008, 74, 2940–2949. [Google Scholar] [CrossRef]
- Yi, Z.; Berney, C.; Hartikainen, H.; Mahamdallie, S.; Gardner, M.; Boenigk, J.; Cavalier-Smith, T.; Bass, D. High-throughput sequencing of microbial eukaryotes in Lake Baikal reveals ecologically differentiated communities and novel evolutionary radiations. FEMS Microbiol. Ecol. 2017, 93, fix073. [Google Scholar] [CrossRef]
- de Sousa, J.R.; Goncalves, V.N.; de Holanda, R.A.; Santos, D.A.; Bueloni, C.F.; Costa, A.O.; Petry, M.V.; Rosa, C.A.; Rosa, L.H. Pathogenic potential of environmental resident fungi from ornithogenic soils of Antarctica. Fungal Biol. 2017, 121, 991–1000. [Google Scholar] [CrossRef]
- Geisen, S.; Mitchell, E.A.D.; Wilkinson, D.M.; Adl, S.; Bonkowski, M.; Brown, M.W.; Fiore-Donno, A.M.; Heger, T.J.; Jassey, V.E.J.; Krashevska, V.; et al. Soil protistology rebooted: 30 fundamental questions to start with. Soil Biol. Biochem. 2017, 111, 94–103. [Google Scholar] [CrossRef]
- Blaalid, R.; Khomich, M. Current knowledge of Chytridiomycota diversity in Northern Europe and future research needs. Fungal Biol. Rev. 2021, 36, 42–51. [Google Scholar] [CrossRef]
- Barr, D. Phylum Chytridiomycota. In Handbook of Protoctista; Margulis, L., Corliss, J., Melkonian, M., Chapman, D.J., Eds.; Jones & Barlett: Burlington, MA, USA, 1990; pp. 454–466. [Google Scholar]
- Gerphagnon, M.; Colombet, J.; Latour, D.; Sime-Ngando, T. Spatial and temporal changes of parasitic chytrids of cyanobacteria. Sci. Rep. 2017, 7, 6056. [Google Scholar] [CrossRef]
- Van den Wyngaert, S.; Rojas-Jimenez, K.; Seto, K.; Kagami, M.; Grossart, H.-P. Diversity and hidden host specificity of chytrids infecting colonial volvocacean algae. J. Eukaryotic Microbiol. 2018, 65, 870–881. [Google Scholar] [CrossRef]
- Bosch, J.; Martínez-Solano, I.; García-París, M. Evidence of a chytrid fungus infection involved in the decline of the common midwife toad (Alytes obstetricans) in protected areas of central Spain. Biol. Conserv. 2001, 97, 331–337. [Google Scholar] [CrossRef]
- Fernández-Beaskoetxea, S.; Bosch, J.; Bielby, J. Infection and transmission heterogeneity of a multi-host pathogen (Batrachochytrium dendrobatidis) within an amphibian community. Dise. Aquat. Organ. 2016, 118, 11–20. [Google Scholar] [CrossRef]
- Freeman, K.R.; Martin, A.P.; Karki, D.; Lynch, R.C.; Mitter, M.S.; Meyer, A.F.; Longcore, J.E.; Simmons, D.R.; Schmidt, S.K. Evidence that chytrids dominate fungal communities in high-elevation soils. Proc. Natl. Acad. Sci. USA 2009, 106, 18315–18320. [Google Scholar] [CrossRef]
- Hendrix, F.F.; Campbell, W.A. Pythium as plant pathogens. Ann. Rev. Phytopathol. 1973, 11, 77–98. [Google Scholar] [CrossRef]
- Levesque, C.A.; de Cock, A.W. Molecular phylogeny and taxonomy of the genus Pythium. Mycol. Res. 2004, 108, 1363–1383. [Google Scholar] [CrossRef]
- Wolinska, J.; Giessler, S.; Koerner, H. Molecular identification and hidden diversity of novel Daphnia parasites from European lakes. Appl. Environ. Microbiol. 2009, 75, 7051–7059. [Google Scholar] [CrossRef] [PubMed]
- Vilela, R.; Taylor, J.W.; Walker, E.D.; Mendoza, L. Lagenidium giganteum pathogenicity in mammals. Emerg. Infect. Dis. 2015, 21, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Martí Carrizosa, M. Candida parapsilosis, C. orthopsilosis y C. metapsilosis: Epidemiología de las Candidemias, Patrones de Sensibilidad y Mecanismos de Resistencia a las Equinocandinas. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2015. [Google Scholar]
- Brieland, J.; McClain, M.; Heath, L.; Chrisp, C.; Huffnagle, G.; LeGendre, M.; Hurley, M.; Fantone, J.; Engleberg, C. Coinoculation with Hartmannella vermiformis enhances replicative Legionella pneumophila lung infection in a murine model of Legionnaires’ disease. Infect. Immun. 1996, 64, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Centeno, M.; Rivera, F.; Cerva, L.; Tsutsumi, V.; Gallegos, E.; Calderón, A.; Ortiz, R.; Bonilla, P.; Ramírez, E.; Suárez, G. Hartmannella vermiformis isolated from the cerebrospinal fluid of a young male patient with meningoencephalitis and bronchopneumonia. Arch. Med. Res. 1996, 27, 579–586. [Google Scholar]
- Poulin, R. The evolution of life history strategies in parasitic animals. Adv. Parasitol. 1996, 37, 107–134. [Google Scholar]
- Altermatt, F.; Pajunen, V.I.; Ebert, D. Desiccation of rock pool habitats and its influence on population persistence in a Daphnia metacommunity. PLoS ONE 2009, 4, e4703. [Google Scholar] [CrossRef]
- Vanschoenwinkel, B.; Gielen, S.; Seaman, M.; Brendonck, L. Wind mediated dispersal of freshwater invertebrates in a rock pool metacommunity: Differences in dispersal capacities and modes. Hydrobiologia 2009, 635, 363–372. [Google Scholar] [CrossRef]
- Bogitsh, B.J.; Carter, C.E.; Oeltmann, T.N. Human Parasitology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Gleason, F.H.; Letcher, P.M.; McGee, P.A. Some Chytridiomycota in soil recover from drying and high temperatures. Mycol. Res. 2004, 108, 583–589. [Google Scholar] [CrossRef]
- Gadd, G.M. Geomycology: Biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 2007, 111, 3–49. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basin | Variable | ASC | CHY | CRY | MUC | HYP | OOM | API | MES | CER | PER | AMO | BIG | FUNGI | PROTISTS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Abun (%) | 1.90 | 53.36 | 0.52 | 10.99 | 0.13 | 1.55 | 0.13 | 0.09 | 31.08 | 0.26 | 55.78 | 44.22 | ||
| OTUs | 19 | 80 | 3 | 7 | 3 | 2 | 1 | 1 | 7 | 2 | 102 | 23 | |||
| [Host] | [3,4,5] | [5] | [4,5] | [4] | [3,5] | [1,2] | [1] | [3] | [0] | [5] | [3,4,5] | [0–5] | |||
| 2 | Abun (%) | 2.29 | 54.67 | 0.12 | 0.21 | 16.34 | 0.82 | 0.82 | 0.04 | 24.69 | 57.29 | 42.71 | |||
| OTUs | 25 | 91 | 6 | 1 | 7 | 13 | 9 | 1 | 9 | 123 | 39 | ||||
| [Host] | [1,3,4] | [5] | [4,5] | [4] | [4] | [3,5] | [1,2] | [3] | [0] | [1,3,4,5] | [0–5] | ||||
| 3 | Abun (%) | 1.41 | 86.59 | 0.08 | 10.70 | 0.32 | 0.60 | 0.01 | 0.01 | 0.10 | 0.17 | 88.09 | 11.91 | ||
| OTUs | 52 | 113 | 4 | 10 | 18 | 6 | 1 | 1 | 1 | 7 | 169 | 44 | |||
| [Host] | [1–5] | [5] | [4,5] | [4] | [1,2,3,5] | [1,2] | [1] | [3] | [5] | [0] | [1–5] | [0–5] | |||
| 4 | Abun (%) | 6.60 | 64.80 | 0.02 | 0.22 | 13.04 | 1.72 | 0.49 | 0.02 | 13.06 | 0.03 | 71.64 | 28.36 | ||
| OTUs | 60 | 98 | 2 | 1 | 8 | 10 | 3 | 1 | 6 | 2 | 161 | 30 | |||
| [Host] | [1,3,4,5] | [5] | [4,5] | [4] | [4] | [2,3,5] | [1] | [5] | [0] | [5] | [1,3,4,5] | [0–5] | |||
| 5 | Abun (%) | 13.79 | 68.82 | 0.05 | 12.54 | 0.62 | 0.20 | 0.01 | 0.02 | 3.95 | 82.66 | 17.34 | |||
| OTUs | 46 | 74 | 3 | 8 | 5 | 3 | 1 | 1 | 5 | 123 | 23 | ||||
| [Host] | [3,4,5] | [5] | [4,5] | [4] | [3,5] | [1,2] | [3] | [5] | [0] | [3,4,5] | [0–5] | ||||
| 6 | Abun (%) | 2.51 | 51.04 | 0.02 | 42.53 | 0.99 | 0.02 | 0.24 | 0.01 | 2.66 | 53.56 | 46.44 | |||
| OTUs | 36 | 79 | 2 | 15 | 8 | 2 | 2 | 1 | 5 | 117 | 33 | ||||
| [Host] | [1,3,4,5] | [5] | [4,5] | [4] | [3,5] | [1,2] | [1] | [3] | [0] | [1,3,4,5] | [0–5] | ||||
| 7 | Abun (%) | 0.56 | 40.68 | 0.10 | 55.33 | 0.03 | 1.40 | 1.88 | 0.02 | 41.34 | 58.66 | ||||
| OTUs | 10 | 43 | 1 | 8 | 2 | 6 | 3 | 1 | 54 | 20 | |||||
| [Host] | [3,5] | [5] | [4,5] | [4] | [3,5] | [1,2] | [0] | [5] | [3,5] | [0–5] | |||||
| 8 | Abun (%) | 2.83 | 86.76 | 9.73 | 0.03 | 0.12 | 0.03 | 0.47 | 89.59 | 10.41 | |||||
| OTUs | 42 | 102 | 9 | 4 | 3 | 2 | 5 | 144 | 23 | ||||||
| [Host] | [1,3,4,5] | [5] | [4] | [3,5] | 1,2] | [5] | [0] | [1,3,4,5] | [0–5] | ||||||
| 9 | Abun (%) | 11.11 | 62.59 | 14.07 | 1.48 | 4.82 | 0.74 | 4.81 | 0.37 | 73.70 | 26.30 | ||||
| OTUs | 8 | 43 | 6 | 4 | 5 | 1 | 5 | 1 | 51 | 22 | |||||
| [Host] | [1,3,5] | [5] | [4] | [3,5] | [1] | [1] | [0] | [5] | [1,3,5] | [0–5] | |||||
| 10 | Abun (%) | 16.22 | 33.25 | 0.05 | 41.41 | 0.86 | 5.59 | 0.10 | 0.05 | 2.47 | 49.52 | 50.48 | |||
| OTUs | 25 | 59 | 1 | 11 | 6 | 3 | 1 | 1 | 5 | 85 | 27 | ||||
| [Host] | [3,4] | [5] | [4,5] | [4] | [3,5] | [1] | [3] | [5] | [0] | [3,4,5] | [0,1,3,4,5] | ||||
| 11 | Abun (%) | 7.74 | 49.69 | 0.16 | 23.88 | 0.45 | 0.08 | 0.04 | 17.95 | 57.60 | 42.40 | ||||
| OTUs | 31 | 71 | 2 | 7 | 7 | 1 | 1 | 6 | 104 | 22 | |||||
| [Host] | [1,3,4,5] | [5] | 4,5] | [4] | [2,3,5] | [1] | [1] | [0] | [1,3,4,5] | [0,1,3,4,5] | |||||
| 12 | Abun (%) | 1.69 | 75.14 | 0.06 | 14.76 | 0.22 | 1.79 | 0.02 | 0.11 | 6.21 | 76.89 | 23.11 | |||
| OTUs | 27 | 79 | 4 | 9 | 10 | 6 | 1 | 2 | 4 | 110 | 32 | ||||
| [Host] | [1,3,4,5] | [5] | [4,5] | [4] | [2,3,5] | [1,2] | [3] | [5] | [0] | [1,3,4,5] | [0–5] | ||||
| 13 | Abun (%) | 7.71 | 57.74 | 21.29 | 1.26 | 5.18 | 6.82 | 65.45 | 34.55 | ||||||
| OTUs | 13 | 29 | 3 | 5 | 4 | 6 | 42 | 18 | |||||||
| [Host] | [1,3,4] | [5] | [4] | [2,3,5] | [1,2] | [0] | [1,3,4,5] | [0–5] | |||||||
| 14 | Abun (%) | 11.55 | 76.14 | 0.10 | 8.42 | 0.14 | 2.37 | 0.76 | 0.52 | 87.79 | 12.21 | ||||
| OTUs | 42 | 82 | 2 | 9 | 5 | 9 | 2 | 5 | 126 | 30 | |||||
| [Host] | [1,3,4,5] | [5] | [4,5] | [4] | [2,3,5] | [1,2] | [5] | [0] | [1,3,4,5] | [0–5] | |||||
| 15 | Abun (%) | 18.68 | 64.55 | 0.03 | 15.26 | 0.19 | 0.06 | 1.24 | 83.26 | 16.74 | |||||
| OTUs | 32 | 54 | 1 | 5 | 4 | 2 | 3 | 87 | 14 | ||||||
| [Host] | [1,3,4] | [5] | [4,5] | [4] | [3,5] | [1] | [0] | [1,3,4,5] | [0,1,3,4,5] | ||||||
| 16 | Abun (%) | 26.40 | 58.17 | 0.23 | 14.05 | 0.42 | 0.11 | 0.01 | 0.02 | 0.03 | 0.56 | 84.80 | 15.20 | ||
| OTUs | 64 | 105 | 6 | 15 | 10 | 4 | 1 | 3 | 2 | 4 | 175 | 39 | |||
| [Host] | [1,3,4,5] | [5] | 4,5] | [4] | [1,3,5] | [1,2] | [1] | [3] | [5] | [0] | [1,3,4,5] | [0–5] | |||
| 17 | Abun (%) | 2.70 | 59.24 | 0.06 | 6.36 | 1.72 | 28.69 | 0.23 | 0.06 | 0.11 | 0.84 | 62.00 | 38.00 | ||
| OTUs | 22 | 67 | 1 | 5 | 8 | 5 | 1 | 1 | 2 | 4 | 90 | 26 | |||
| [Host] | [3,4,5] | [5] | [4,5] | [4] | [2,3,5] | [1,2] | [1] | [3] | [5] | [0] | [3,4,5] | [0–5] | |||
| 18 | Abun (%) | 16.08 | 35.67 | 4.53 | 16.23 | 17.25 | 0.73 | 8.63 | 0.88 | 56.29 | 43.71 | ||||
| OTUs | 10 | 43 | 4 | 3 | 7 | 3 | 1 | 2 | 57 | 16 | |||||
| [Host] | [1,3] | [5] | [4,5] | [4] | [2,3,5] | [1,2] | [1] | [0] | [1,3,5] | [0–5] | |||||
| 19 | Abun (%) | 7.46 | 31.54 | 0.98 | 54.16 | 1.83 | 0.37 | 1.59 | 0.12 | 0.37 | 1.59 | 39.98 | 60.02 | ||
| OTUs | 13 | 40 | 5 | 6 | 4 | 3 | 2 | 1 | 1 | 4 | 58 | 21 | |||
| [Host] | [1,3,4,5] | [5] | [4,5] | [4] | [2,3,5] | [1,2] | [1] | [3] | [5] | [0] | [1,3,4,5] | [0–5] | |||
| 20 | Abun (%) | 31.38 | 41.18 | 5.06 | 17.78 | 1.01 | 1.98 | 0.01 | 0.26 | 0.10 | 1.25 | 77.61 | 22.39 | ||
| OTUs | 55 | 86 | 6 | 13 | 16 | 7 | 1 | 3 | 1 | 5 | 147 | 46 | |||
| [Host] | [2,3,4,5] | [5] | [4,5] | [4] | [2,3,5] | [1,2] | [1] | [3] | [5] | [0] | [2,3,4,5] | [0–5] | |||
| 21 | Abun (%) | 3.61 | 94.38 | 0.15 | 0.84 | 0.07 | 0.33 | 0.26 | 0.37 | 98.14 | 1.86 | ||||
| OTUs | 22 | 50 | 2 | 3 | 2 | 3 | 1 | 3 | 74 | 12 | |||||
| [Host] | [1,3,5] | [5] | [4,5] | [4] | [3] | [1] | [5] | [0] | [1,3,5] | [0,1,3,4,5] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velasco-González, I.; Lara, E.; Singer, D.; de Cos-Gandoy, A.; García-Rodríguez, M.; Murciano, A.; Pérez-Uz, B.; Williams, R.; Sanchez-Jimenez, A.; Martín-Cereceda, M. Diversity of DNA Sequences from Pathogenic and Potentially Pathogenic Eukaryotic Microorganisms in Protected Granite Mountain Rocks. Diversity 2023, 15, 594. https://doi.org/10.3390/d15050594
Velasco-González I, Lara E, Singer D, de Cos-Gandoy A, García-Rodríguez M, Murciano A, Pérez-Uz B, Williams R, Sanchez-Jimenez A, Martín-Cereceda M. Diversity of DNA Sequences from Pathogenic and Potentially Pathogenic Eukaryotic Microorganisms in Protected Granite Mountain Rocks. Diversity. 2023; 15(5):594. https://doi.org/10.3390/d15050594
Chicago/Turabian StyleVelasco-González, Ismael, Enrique Lara, David Singer, Amaya de Cos-Gandoy, Manuel García-Rodríguez, Antonio Murciano, Blanca Pérez-Uz, Richard Williams, Abel Sanchez-Jimenez, and Mercedes Martín-Cereceda. 2023. "Diversity of DNA Sequences from Pathogenic and Potentially Pathogenic Eukaryotic Microorganisms in Protected Granite Mountain Rocks" Diversity 15, no. 5: 594. https://doi.org/10.3390/d15050594
APA StyleVelasco-González, I., Lara, E., Singer, D., de Cos-Gandoy, A., García-Rodríguez, M., Murciano, A., Pérez-Uz, B., Williams, R., Sanchez-Jimenez, A., & Martín-Cereceda, M. (2023). Diversity of DNA Sequences from Pathogenic and Potentially Pathogenic Eukaryotic Microorganisms in Protected Granite Mountain Rocks. Diversity, 15(5), 594. https://doi.org/10.3390/d15050594