

Detection of a Diverse Endophyte Assemblage within Fungal Communities Associated with the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae)

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Communities Associated with L. donacis
2.2. Fungal Communities Isolated from Leaf Sheath of Giant Reed in Texas
2.3. Fungal Isolation
2.4. Conidia Associated with the Ovipositor and the Ovarioles of L. donacis Females
2.5. DNA Extraction, PCR, and Sequencing
2.6. Sequence Data Analysis
3. Results
3.1. Assemblage of Fungal Endophytes Inhabiting the Fungal Communities Associated with L. donacis and Leaf Sheath of Giant Reed
3.2. Lasioptera Donacis Is Carrying Endophyte Conidiae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, G.P. Ecology and management of Arundo donax, and approaches to riparian habitat restoration in southern California. In Plant Invasions: Studies from North America and Europe; Brock, J.H., Wade, M., Pysek, P., Green, D., Eds.; Blackhuys Publishers: Leiden, The Netherlands, 1997; pp. 103–113. [Google Scholar]
- Racelis, A.; Rubio, A.; Vaughan, T.; Goolsby, J. Passive restoration potential of areas invaded with giant reed in south Texas. Ecol. Restor. 2012, 30, 112–116. [Google Scholar] [CrossRef]
- Watts, D.A.; Moore, G.W. Water use dynamics of an invasive reed, Arundo donax, from leaf to stand. Wetlands 2011, 31, 725–734. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, J.; Hardion, L.; Monte, J.; Vila, B.; Santín-Montanyá, M. Monographs on invasive plants in Europe N° 4: Arundo donax L. Bot. Lett. 2021, 168, 131–151. [Google Scholar] [CrossRef]
- Goolsby, J.A.; Moran, P.J. Host range of Tetramesa romana Walker (Hymenoptera: Eurytomidae), a potential biological control ofgiant reed, Arundo donax L. in North America. Biol. Control 2009, 49, 160–168. [Google Scholar] [CrossRef]
- Goolsby, J.; Vacek, A.T.; Salinas, C.; Racelis, A.E.; Moran, P.J.; Kirk, A. Host range of the European leaf sheath mining midge, Lasioptera donacis Coutin, a biological control of giant reed, Arundo donax. Biocontrol Sci. Technol. 2017, 27, 781–795. [Google Scholar] [CrossRef]
- Marshall, M.; Goolsby, J.A.; Vacek, A.T.; Mastoras, A.; Kashefi, J.; Chaskopoulou, A.; Smith, L.; Badillo, I.; Reilly, F.J.; Perez de Leon, A.A.; et al. Biotic and abiotic factors influencing infestation levels of the arundo leafminer, Lasioptera donacis, in its native range in Mediterranean Europe. Subtrop. Agric. Environ. 2018, 69, 8–18. [Google Scholar]
- Bissett, J.; Borkent, A. galls: The significance of fungal nutrition in the evolution of the Cecidomyiidae (Diptera). In Coevolution of Fungi with Plants and Animals; Pirozynski, K.A., Hawksworth, D.L., Eds.; Academic Press: London, UK, 1988; pp. 203–205. [Google Scholar]
- Stireman, J.O.; Devlin, H.; Carr, T.G.; Abbot, P. Evolutionary diversification of the gall midge genus Asteromyia (Cecidomyiidae) in a multitrophic ecological context. Mol. Phylogenetic Evol. 2010, 54, 194–210. [Google Scholar] [CrossRef]
- Gagné, R.J.; Jaschhof, M. A Catalog of the Cecidomyiidae (Diptera) of the World, 5th ed.; Digital; Systematic Entomology Laboratory, Agricultural Research Service, U.S. Department of Agriculture: Washington, DC, USA, 2021; 816p. Available online: https://www.ars.usda.gov/ARSUserFiles/80420580/Gagne_Jaschhof_2021_World_Cat_5th_Ed.pdf (accessed on 12 December 2022).
- Skuhravá, M.; Skuhravy, V. Species richness of gall midges (Diptera, Cecidomyiidae) in Europe (West Palaeartic): Biogeography and coevolution with host plants. Acta Soc. Zool. Bohem. 2010, 73, 87–156. [Google Scholar]
- Doğanlar, M.; Sertkaya, E.; Skuhravá, M. Pest status of olive leaf gall midge Dasineura oleae (Angelini, 1831), description of Lasioptera oleicola Skuhravá sp. new (Diptera: Cecidomyiidae) and effectiveness of parasitoids on their populations in Hatay Turkey. Türkiye Entomoloji Derg. 2011, 35, 265–284. [Google Scholar]
- Meyer, J. Cecidogenese de la galle de Lasioptera rubi Reeger et role nourricier d’un mycelium symbiotique. Comptes Rendus Des Séances L’académie Sci. 1952, 234, 2556–2558. [Google Scholar]
- Rohfritsch, O. A fungus associated gall midge, Lasioptera arundinis (Schiner), on Phragmites australis (Cav.) Trin. Bull. De La Société Bot. Fr. 1992, 139, 45–59. [Google Scholar] [CrossRef]
- Rohfritsch, O. Morphological and behavioural adaptations of the gall midge Lasioptera arundinis (Schiner) (Diptera, Cecidomyiidae) to collect and transport conidia of its fungal symbiont. Tijdschr. Voor Entomol. 1997, 140, 59–66. [Google Scholar]
- Rohfritsch, O. Plants, gall midges, and fungi: A three-component system. Entomol. Exp. Appl. 2008, 128, 208–216. [Google Scholar] [CrossRef]
- Hulcr, J.; Stelinski, L.L. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef]
- Borkent, A.; Bisset, J. Gall Midges (Diptera: Cecidomyiidae) are vectors for their fungal symbionts. Symbiosis 1985, 1, 185–194. [Google Scholar]
- Coutin, R. Une nouvelle Cécidomyie des gaines foliaires de la Canne de Provence, Lasioptera donacis n. sp. (Diptera, Cecidomyiidae). Bull. Société Entomol. Fr. 2001, 106, 105–108. [Google Scholar] [CrossRef]
- Thomas, D.; Goolsby, J. Morphology of the Preimaginal Stages of Lasioptera donacis Coutin (Diptera: Cecidomyiidae), a Candidate Biocontrol Agent for Giant Arundo Cane. Psyche J. Entomol. 2015, 2015, 262678. [Google Scholar] [CrossRef]
- Careddu, G.; Botti, M.; Cristofaro, M.; Sporta Caputi, S.; Calizza, E.; Rossi, L.; Costantini, M.L. The Feeding Behaviour of Gall MidgeLarvae and Its Implications forBiocontrol of the Giant Reed: Insightsfrom Stable Isotope Analysis. Biology 2022, 11, 1805. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Ryberg, M.; Abarenkov, K.; Sjokvist, E.; Kristiansson, E. The ITS region as a target for characterization of fungal communities using emerging sequencing technologies. FEMS Microbiol. Lett. 2010, 296, 97–101. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium; Fungal Barcoding Consortium Author List; Bolchacova, E.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.H.; Kjøller, R.; Larsson, E.; Pennanen , T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics In PCR Protocols. A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenetics Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef]
- Hibbett, D.; Glotzer, D. Where are all the undocumented fungal species? A study of Mortierella demonstrates the need for sequence-based classification. New Phytol. 2011, 191, 592–596. [Google Scholar] [CrossRef]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Abarenkov, K.; Tedersoo, L.; Nilsson, R.H.; Vellak, K.; Saar, I.; Veldre, V.; Parmasto, E.; Prous, M.; Aan, A.; Ots, M.; et al. PlutoF —A Web Based Workbench for Ecological and Taxonomic Research, with an Online Implementation for Fungal ITS Sequences. Evol. Bioinform. 2010, 6, 189–196. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41–D47. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. New York In Mammalian Protein Metabolism III; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Pintos, Á.; Alvarado, P.; Planas, J.; Jarling, R. Six new species of Arthrinium from Europe and notes about A. caricicola and other species found in Carex spp. hosts. MycoKeys 2019, 49, 15–48. [Google Scholar] [CrossRef]
- Pintos, Á.; Alvarado, P. Phylogenetic delimitation of Apiospora and Arthrinium. Fungal Syst. Evol. 2021, 7, 197–221. [Google Scholar] [CrossRef]
- Bon, M.C.; Guermache, F.; de Simone, D.; Cristofaro, M.; Vacek, A.; Goolsby, J. Insights into the Microbes and Nematodes Hosted by Pupae of the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae). Fla. Entomol. 2018, 101, 505–507. [Google Scholar] [CrossRef]
- Hibbett, D.; Binder, M.; Bischoff, J.; Blackwell, M.; Cannon, P.; Eriksson, O.; Huhndorf, S.M.; James, T.; Kirk, P.; Lücking, R. A higher-level phylogenetic classification of the Fungi. Mycol Res 2006, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 6 May 2022).
- Crous, P.W.; Groenewald, J.Z. A phylogenetic re-evaluation of Arthrinium. IMA Fungus 2013, 4, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tan, X.M.; Liu, F.; Cai, L. Eight new Arthrinium species from China. Mycokeys 2018, 34, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Clay, K. Fungal Endophytes of Grasses. Annu. Rev. Ecol. Syst. 1990, 21, 275–297. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C.L. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef]
- Sanchez Marquez, S.; Bills, G.F.; Dominguez Acuna, L.; Zabalgogeazcoa, I. Endophytic mycobiota of leaves and roots of the grass Holcus Lanatus. Fungal Divers. 2010, 41, 115–123. [Google Scholar] [CrossRef]
- Kuo, H.-C.; Hui, S.; Choi, J.; Asiegbu, F.O.; Valkonen, J.P.T.; Lee, Y.-H. Secret lifestyles of Neurospora crassa. Sci. Rep. 2014, 4, 5135. [Google Scholar] [CrossRef]
- Zou, J.; Li, X.; Huang, P.W.; Dai, C.C. Endophytism or saprophytism: Decoding the lifestyle transition of the generalist fungus Phomopsis Liq. Microbiol. Res. 2018, 206, 99–112. [Google Scholar] [CrossRef]
- Thangam, D.S.; Selvakumar, G.; Verghese, A.; Jayanthi, P.D.K. Natural mycosis of mango leafhoppers (Cicadellidae: Hemiptera) by Fusarium sp. Biocontrol Sci. Technol. 2014, 24, 229–232. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.R.G.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef]
- Tosi, L.; Beccari, G.; Rondoni, G.; Covarelli, L.; Ricci, C. Natural occurrence of Fusarium proliferatum on chestnut in Italy and its potential entomopathogenicity against the Asian chestnut gall wasp Dryocosmus kuriphilus. J. Pest Sci. 2014, 88, 369–381. [Google Scholar] [CrossRef]
- Dyko, B.J.; Sutton, B.C. New and interesting dematiaceous hyphomycetes from Florida. Mycotaxon 1979, 8, 119–124. [Google Scholar]
- Martínez-Cano, C.; Grey, W.E.; Sands, D.C. First report of Arthrinium arundinis causing kernel blight on barley. Plant Dis. 1992, 76, 1077. [Google Scholar] [CrossRef]
- Ramos, H.P.; Braun, G.H.; Pupo, M.T.; Said, S. Antimicrobial activity from endophytic fungi Arthrinium state of Apiospora montagnei Sacc. and Papulaspora immersa. Braz. Arch. Biol. Technol. 2010, 53, 629–632. [Google Scholar] [CrossRef]
- Cristofaro, M.; Goolsby, J.; Thomas, D.; Vacek, A.; Salinas, C.; Perez de Leon, A.; Summy, R.; Racelis, A.; Kirk, A.; Mercadier, G.; et al. Biological studies and field observations in Europe of Lasioptera donacis, a potential biological control giant reed, Arundo donax, an invasive weed of the Rio Grande Basin of Texas and Mexico. In Proceedings of the XIV International Symposium on the Biological Control of Weeds, Cape Town, South Africa, 2–7 March 2014; p. 15. [Google Scholar]
- Yeh, Y.H.; Kirschner, R. Sarocladium spinificis, a new endophytic species from the coastal grass Spinifex littoreus in Taiwan. Bot. Stud. 2014, 56, 55. [Google Scholar] [CrossRef]
- Maia, N.C.; Souza, P.N.C.; Godinho, B.T.V.; Moreira, S.I.; Abreu, L.M.; Jank, L.; Cardoso, P.G. Fungal endophytes of Panicum maximum and Pennisetum purpureum: Isolation, identification, and determination of antifungal potential. Rev. Bras. Zootech. 2018, 47, e20170183. [Google Scholar] [CrossRef]
- Teasdale, S.E.; Caradus, J.R.; Johnson, L.J. Fungal endophyte diversity from tropical forage grass Brachiaria. Plant Ecol. Divers. 2018, 11, 611–624. [Google Scholar] [CrossRef]
- Pelaez, F.; Collado, J.; Arenal, F.; Basilio, A.; Cabello, A.; Diez Matas, M.T.; Garcia, J.B.; Gonzalez Del Val, A.; Gonzalez, V.; Gorrochategui, J.; et al. Endophytic fungi from plants living on gypsum soils as a source of secondary metabolites with antimicrobial activity. Mycol. Res. 1998, 102, 755–761. [Google Scholar] [CrossRef]
- Busby, P.E.; Ridout, M.; Newcombe, G. Fungal endophytes: Modifiers of plant disease. Plant Mol. Biol. 2016, 90, 645–655. [Google Scholar] [CrossRef]
- Currie, A.F.; Gange, A.C.; Ab Razak, N.; Ellison, C.A.; Maczey, N.; Wood, S.V. Endophytic fungi in the invasive weed Impatiens glandulifera: A barrier to classical biological control? Weed Res. 2020, 60, 50–59. [Google Scholar] [CrossRef]
- Gange, A.C.; Koricheva, J.; Currie, A.F.; Jaber, L.R.; Vidal, S. Meta-analysis of the role of entomopathogenic and unspecialized fungal endophytes as plant bodyguards. New Phytol. 2019, 223, 2002–2010. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.; Faeth, S.H. Do Fungal Endophytes Result in Selection for Leaf miner Ovipositional Preference? Ecology 2001, 82, 1097–1111. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.; Fu, C.; Liu, M.; Xing, Q.; et al. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MOTU Number/ Reference UNITE Fungal Species Hypothesis (SH) | Blast Similarity Score % (E-Value = 0) | Fungal Species/Class/Order | Genbank Accession Number |
|---|---|---|---|
| MOTU1/SH1541923.08FU | 99.4–100 | Sarocladium terricola/ Sordariomycetes/ Hypocreales | OP970564 to OP970582 |
| MOTU2/SH1541921.08FU | 99.65 | Sarocladium strictum/ Sordariomycetes/ Hypocreales | OP970583 & OP970584 |
| MOTU3/SH1541924.08FU | 99.65 | Sarocladium spinificis/ Sordariomycetes/ Hypocreales | OP970585 to OP970587 |
| MOTU4/SH1541928.08FU | 99.30 | Nectria sp./ Sordariomycetes/ Hypocreales | OP970588 & OP970589 |
| MOTU5/SH1567483.08FU | 99.62–100 | Fusarium sporotrichioides/ Sordariomycetes/ Hypocreales | OP970590 to OP970596 |
| MOTU6/SH1610159.08FU | 99.63–100 | Fusarium proliferatum/ Sordariomycetes/ Hypocreales | OP970597 to OP970615 |
| MOTU7/SH1610157.08FU | 100 | Fusarium sp./ Sordariomycetes/ Hypocreales | OP970616 |
| MOTU8/SH1505921.08FU | 99.32 | Alternaria sp./ Sordariomycetes/ Hypocreales | OP970617 |
| MOTU9/SH1526398.08FU | 100 | Alternaria alternata/ Sordariomycetes/ Hypocreales | OP970618 to OP970621 |
| MOTU10/SH1507367.08FU | 100 | Botryosphaeria dothidea/ Dothideomycetes/ Botryosphaeriales | OP970622 to OP970625 |
| MOTU11/SH1615599.08FU | 100 | Chaetomium globosum/ Euascomycetes/ Sordariales | OP970626 |
| MOTU12/SH1572816.08FU | 100 | Cladosporium cladosporioides/ Dothideomycetes/ Capnodiales | OP970627 & OP970628 |
| MOTU13/SH1572792.08FU | 100 | Cladosporium velox/ Dothideomycetes/ Capnodiales | OP970629 & OP970630 |
| MOTU14/SH1572792.08FU | 100 | Cladosporium aciculare/ Dothideomycetes/ Capnodiales | OP970631 |
| MOTU15/SH1572820.08FU | 100 | Cladosporium ramotenellum/ Dothideomycetes/ Capnodiales | OP970632 |
| MOTU16/SH1526406.08FU | 100 | Bipolaris sp./ Dothideomycetes/ Pleosporales | OP970633 & OP970634 |
| MOTU17/SH1524421.08FU | 99.82 | Lecanicillium lecanii/ Sordariomycetes/ Hypocreales | OP970635 & OP970636 |
| MOTU18/SH1547057.08FU | 100 | Epicoccum italicum/ Dothideomycetes/ Pleosporales | OP970637 |
| MOTU19/SH1516144.08FU | 99.63–100 | Penicilium funiculosus syn: Talaromyces funiculosus/ Eurotiomycetes/Eurotiales | OP970638 to OP970644 |
| MOTU20/SH1529984.08FU | 100 | Penicilium rubens/ /Eurotiomycetes/Eurotiales | OP970645 & OP970646 |
| MOTU21/SH1530001.08FU | 100 | Penicilium terrigenum/ /Eurotiomycetes/Eurotiales | OP970647 |
| MOTU22/SH1547057.08FU | 100 | Phoma sp./ Dothideomycetes/Paleopoles | OP970648 |
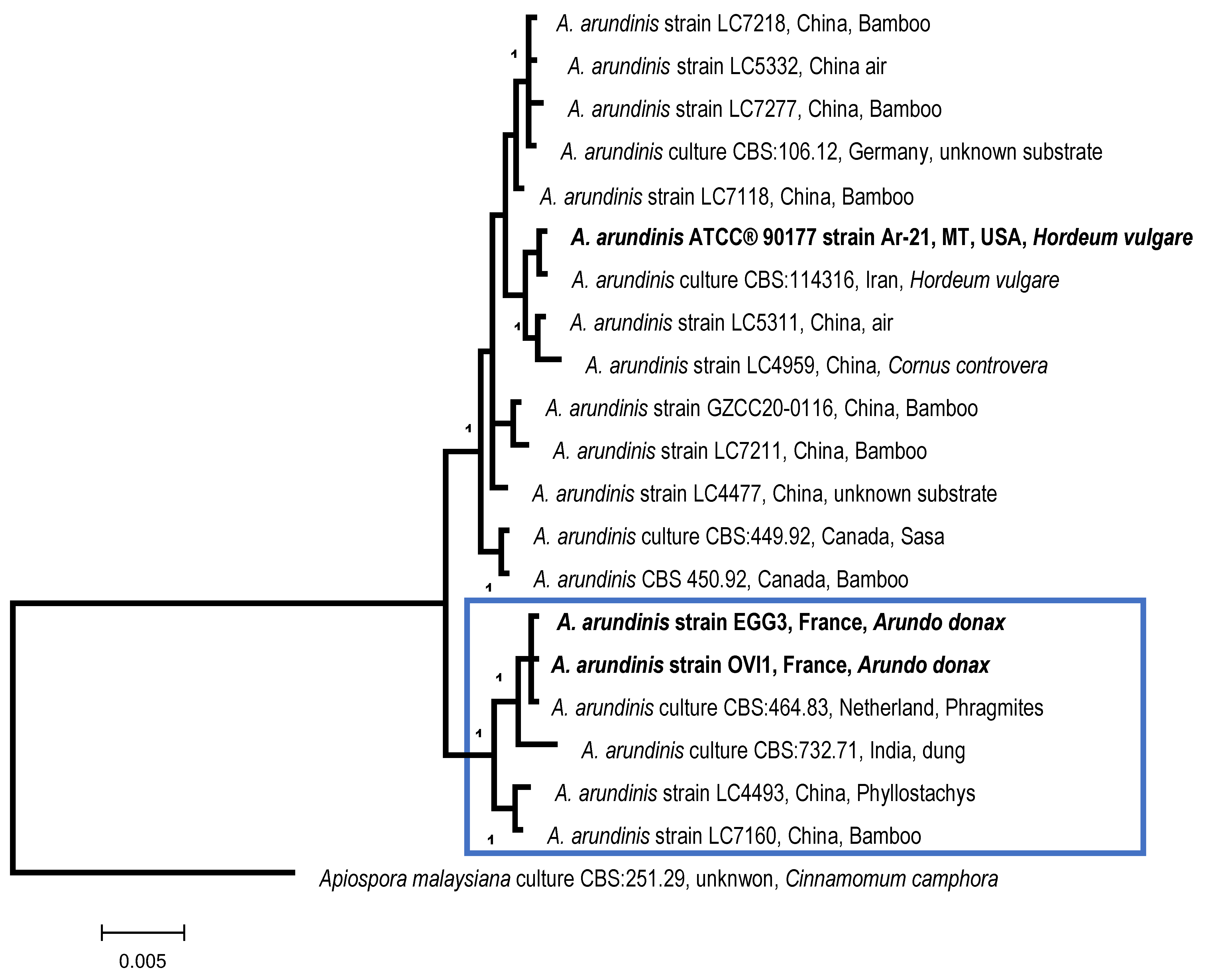
| MOTU23/SH1540045.08FU | 100 | Apiospora arundinis syn:
Arthrinium arundinis/ Sordariomycetes/Xylariales | OP970649 to OP970653 MF627422 [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bon, M.-C.; Goolsby, J.A.; Mercadier, G.; Guermache, F.; Kashefi, J.; Cristofaro, M.; Vacek, A.T.; Kirk, A. Detection of a Diverse Endophyte Assemblage within Fungal Communities Associated with the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae). Diversity 2023, 15, 571. https://doi.org/10.3390/d15040571
Bon M-C, Goolsby JA, Mercadier G, Guermache F, Kashefi J, Cristofaro M, Vacek AT, Kirk A. Detection of a Diverse Endophyte Assemblage within Fungal Communities Associated with the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae). Diversity. 2023; 15(4):571. https://doi.org/10.3390/d15040571
Chicago/Turabian StyleBon, Marie-Claude, John A. Goolsby, Guy Mercadier, Fatiha Guermache, Javid Kashefi, Massimo Cristofaro, Ann T. Vacek, and Alan Kirk. 2023. "Detection of a Diverse Endophyte Assemblage within Fungal Communities Associated with the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae)" Diversity 15, no. 4: 571. https://doi.org/10.3390/d15040571
APA StyleBon, M.-C., Goolsby, J. A., Mercadier, G., Guermache, F., Kashefi, J., Cristofaro, M., Vacek, A. T., & Kirk, A. (2023). Detection of a Diverse Endophyte Assemblage within Fungal Communities Associated with the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae). Diversity, 15(4), 571. https://doi.org/10.3390/d15040571