


Constitutive Innate Immunity of Migrant and Resident Long-Nosed Bats (Leptonycteris yerbabuenae) in the Drylands of Mexico


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Sample Collection
2.2. Measurement of Bacteria-Killing Ability
2.3. Data Analyses
3. Results
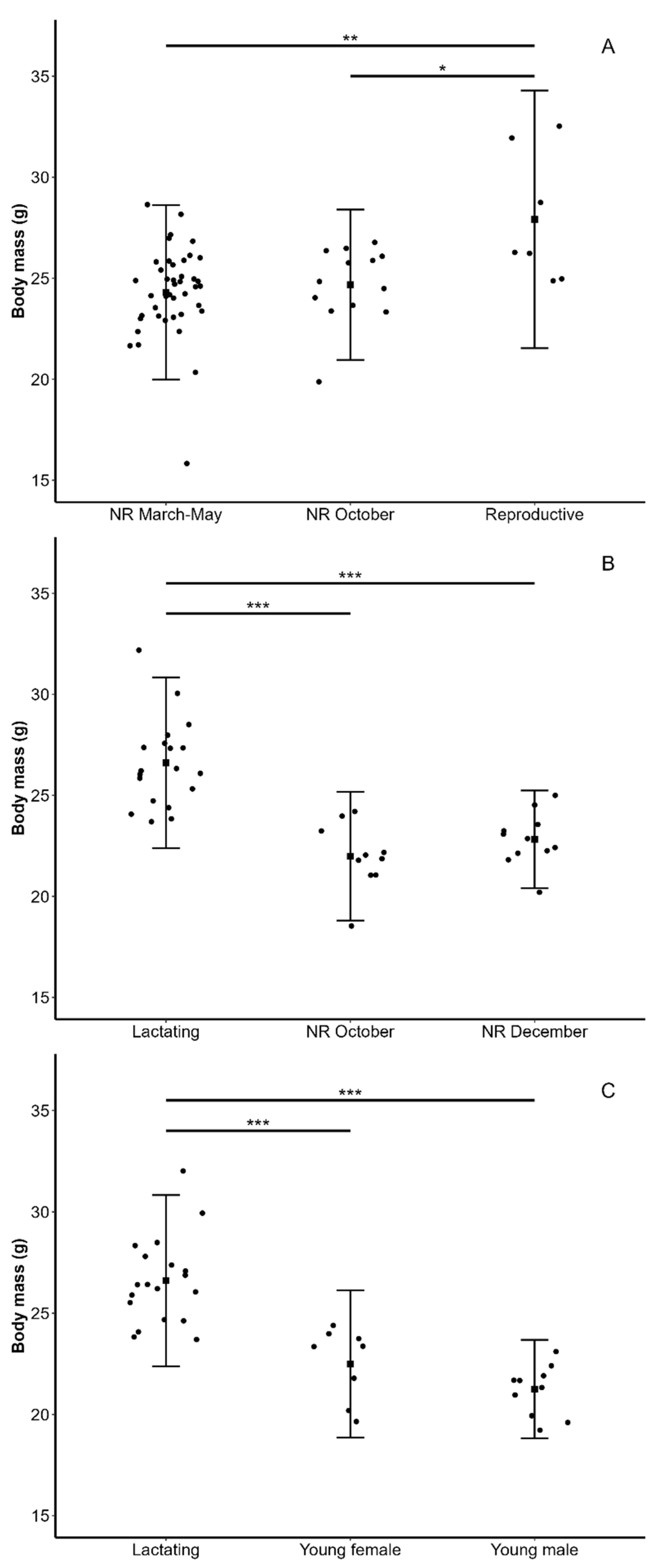
3.1. Reproductive Status and Body Mass
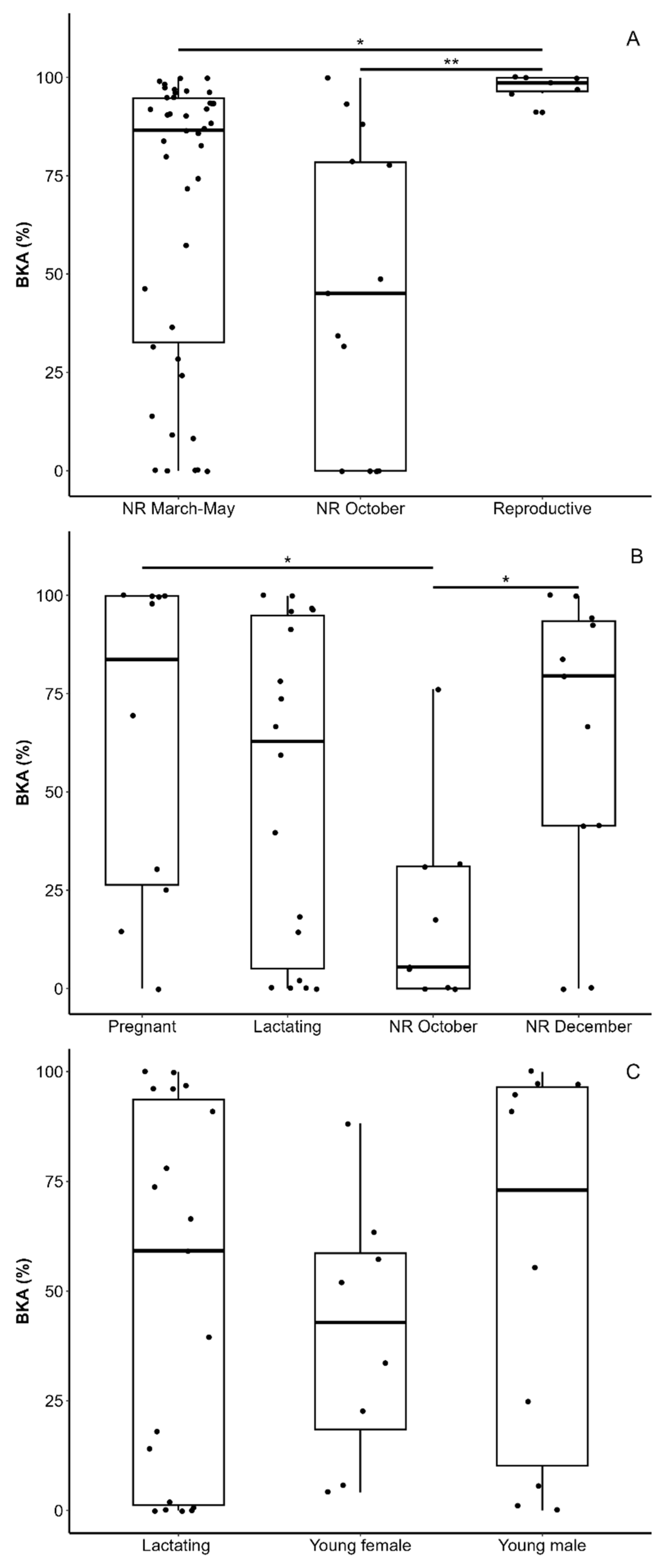
3.2. Bacteria-Killing Ability
4. Discussion
4.1. Male BKA
4.2. Female BKA
4.3. BKA of Young and Lactating Females
4.4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neuweiler, G. Ecology. In Biology of Bats; Oxford University Press: New York, NY, USA, 2000; ISBN 0195099508. [Google Scholar]
- Altizer, S.; Bartel, R.; Han, B.A. Animal migration and infectious disease risk. Science 2011, 331, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Binning, S.A. Migratory recovery from infection as a selective pressure for the evolution of migration. Am. Nat. 2016, 187, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Binning, S.A. Recovery from infection is more likely to favour the evolution of migration than social escape from infection. J. Anim. Ecol. 2020, 89, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R.; de Angeli Dutra, D. Animal migrations and parasitism: Reciprocal effects within a unified framework. Biol. Rev. 2021, 96, 1331–1348. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.J.; Altizer, S.; Peacock, S.J.; Shaw, A.K. Animal migration and infection dynamics: Recent advances and future frontiers. In Animal Behavior and Parasitism; Ezenwa, V., Altizer, S.M., Hall, R., Eds.; Oxford University Press: Oxford, UK, 2022; pp. 111–132. ISBN 9780192895561. [Google Scholar]
- Dingle, H.; Drake, A. What is migration? Bioscience 2007, 57, 113–121. [Google Scholar] [CrossRef]
- Kelly, T.R.; MacGillivray, H.L.; Hobson, K.A.; MacDougall-Shackleton, S.A.; MacDougall-Shackleton, E.A. Immune profiles vary seasonally, but are not significantly related to migration distance or natal dispersal, in a migratory songbird. J. Exp. Zool. 2017, 327, 284–292. [Google Scholar] [CrossRef]
- Avgar, T.; Street, G.; Fryxell, J.M. On the adaptive benefits of mammal migration. Can. J. Zool. 2014, 92, 481–490. [Google Scholar] [CrossRef]
- Fleming, T.H.; Eby, P. Ecology of Bat Migration. In Bat Ecology; Kunz, T.H., Fenton, B.M., Eds.; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 156–197. ISBN 0-226-46206-4. [Google Scholar]
- Popa-Lisseanu, A.G.; Voigt, C.C. Bats on the move. J. Mammal. 2009, 90, 1283–1289. [Google Scholar] [CrossRef]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef]
- Mühldorfer, K. Bats and bacterial pathogens: A review. Zoonoses Public Health 2013, 60, 93–103. [Google Scholar] [CrossRef]
- Paweska, J.T.; Storm, N.; Grobbelaar, A.A.; Markotter, W.; Kemp, A.; van Vuren, P.J. Experimental inoculation of Egyptian fruit bats (Rousettus aegyptiacus) with ebola virus. Viruses 2016, 8, 29. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Schwarz, T.M.; Ilinykh, P.A.; Jordan, I.; Ksiazek, T.G.; Sachidanandam, R.; Basler, C.F.; Bukreyev, A. Innate immune responses of bat and human cells to filoviruses: Commonalities and distinctions. J. Virol. 2017, 91, e02471-16. [Google Scholar] [CrossRef]
- Hegemann, A.; Fudickar, A.M.; Nilsson, J.Å. A physiological perspective on the ecology and evolution of partial migration. J. Ornithol. 2019, 160, 893–905. [Google Scholar] [CrossRef]
- de Angeli Dutra, D.; Fecchio, A.; Martins Braga, É.; Poulin, R. Migratory birds have higher prevalence and richness of avian haemosporidian parasites than residents. Int. J. Parasitol. 2021, 51, 877–882. [Google Scholar] [CrossRef]
- Owen, J.C.; Moore, F.R. Swainson’s thrushes in migratory disposition exhibit reduced immune function. J. Ethol. 2008, 26, 383–388. [Google Scholar] [CrossRef]
- Eikenaar, C.; Hegemann, A. Migratory common blackbirds have lower innate immune function during autumn migration than resident conspecifics. Biol. Lett. 2016, 12, 78–81. [Google Scholar] [CrossRef]
- Eikenaar, C.; Isaksson, C.; Hegemann, A. A hidden cost of migration? innate immune function versus antioxidant defense. Ecol. Evol. 2018, 8, 2721–2728. [Google Scholar] [CrossRef]
- Voigt, C.C.; Fritze, M.; Lindecke, O.; Costantini, D.; Pētersons, G.; Czirják, G. The immune response of bats differs between pre-migration and migration seasons. Sci. Rep. 2020, 10, 17384. [Google Scholar] [CrossRef]
- Rogers, E.J.; McGuire, L.; Longstaffe, F.J.; Clerc, J.; Kunkel, E.; Fraser, E. Relating wing morphology and immune function to patterns of partial and differential bat migration using stable isotopes. J. Anim. Ecol. 2022, 91, 858–869. [Google Scholar] [CrossRef]
- Christe, P.; Arlettaz, R.; Vogel, P. Variation in intensity of a parasitic mite (Spinturnix myoti) in relation to the reproductive cycle and immunocompetence of its bat host (Myotis myotis). Ecol. Lett. 2000, 3, 207–212. [Google Scholar] [CrossRef]
- Otálora-Ardila, A.; Flores-Martínez, J.J.; Rosales, C.; Salame-Méndez, A.; Herrera M., L.G. Physiological and ecological correlates of the cellular and humoral innate immune responses in an insular desert bat: The fish-eating myotis (Myotis vivesi). Diversity 2022, 14, 781. [Google Scholar] [CrossRef]
- Ruoss, S.; Becker, N.I.; Otto, M.S.; Czirják, G.; Encarnação, J.A. Effect of sex and reproductive status on the immunity of the temperate bat Myotis daubentonii. Mamm. Biol. 2019, 94, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.J.; Czirják, G.; Volokhov, D.V.; Bentz, A.B.; Carrera, J.E.; Camus, M.S.; Navara, K.J.; Chizhikov, V.E.; Fenton, M.B.; Simmons, N.B.; et al. Livestock abundance predicts vampire bat demography, immune profiles and bacterial infection risk. Phil. Trans. R. Soc. B 2018, 373, 20170089. [Google Scholar] [CrossRef] [PubMed]
- Medellín, R. Leptonycteris yerbabuenae. The IUCN Red List of Threatened Species. 2016, e.T136659A21988965. Available online: https://www.iucnredlist.org/species/136659/21988965 (accessed on 10 September 2022).
- Morales-Garza, M.R.; del C. Arizmendi, M.; Campos, J.E.; Martínez-Garcia, M.; Valiente-Banuet, A. Evidences on the migratory movements of the nectar-feeding bat Leptonycteris curasoae in Mexico using random amplified polymorphic DNA (RAPD). J. Arid Environ. 2007, 68, 248–259. [Google Scholar] [CrossRef]
- Wilkinson, G.S.; Fleming, T.H. Migration and evolution of lesser longnosed bats Leptonycteris curasoae, inferred from mitochondrial DNA. Mol. Ecol. 1996, 5, 329–339. [Google Scholar] [CrossRef]
- Menchaca, A.; Arteaga, M.C.; Medellin, R.A.; Jones, G. Conservation units and historical matrilineal structure in the tequila bat (Leptonycteris yerbabuenae). Glob. Ecol. Conserv. 2020, 23, e01164. [Google Scholar] [CrossRef]
- Trejo-Salazar, R.E.; Castellanos-Morales, G.; Hernández-Rosales, D.C.; Gámez, N.; Gasca-Pineda, J.; Morales Garza, M.R.; Medellin, R.; Eguiarte, L.E. Discordance in maternal and paternal genetic markers in lesser long-nosed bat Leptonycteris yerbabuenae, a migratory bat: Recent expansion to the north and male phylopatry. PeerJ 2021, 9, e12168. [Google Scholar] [CrossRef]
- Stoner, K.E.; Karla, K.A.; Roxana, R.C.; Quesada, M. Population dynamics, reproduction, and diet of the lesser long-nosed bat (Leptonycteris curasoae) in Jalisco, Mexico: Implications for conservation. Biodivers. Conserv. 2003, 12, 357–373. [Google Scholar] [CrossRef]
- Millet, S.; Bennett, J.; Lee, K.A.; Hau, M.; Klasing, K.C. Quantifying and comparing constitutive immunity across avian species. Dev. Comp. Immunol. 2007, 31, 188–201. [Google Scholar] [CrossRef]
- Tieleman, B.I.; Williams, J.B.; Ricklefs, R.E.; Klasing, K.C. Constitutive innate immunity is a component of the pace-of-life syndrome in tropical birds. Proc. R. Soc. B 2005, 272, 1715–1720. [Google Scholar] [CrossRef]
- Moore, M.S.; Reichard, J.D.; Murtha, T.D.; Zahedi, B.; Fallier, R.M.; Thomas, H. Specific alterations in complement protein activity of little brown myotis (Myotis lucifugus) hibernating in white-nose syndrome affected sites. PLoS ONE 2011, 6, e27430. [Google Scholar] [CrossRef]
- Merchant, M.E.; Roche, C.; Elsey, R.M.; Prudhomme, J. Antibacterial properties of serum from the american alligator (Alligator mississippiensis). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 505–513. [Google Scholar] [CrossRef]
- Foo, Y.Z.; Nakagawa, S.; Rhodes, G.; Simmons, L.W. The effects of sex hormones on immune function: A meta-analysis. Biol. Rev. 2016, 92, 551–571. [Google Scholar] [CrossRef]
- Ruiz, M.; French, S.S.; Demas, G.E.; Martins, E.P. Food supplementation and testosterone interact to influence reproductive behavior and immune function in Sceloporus graciosus. Horm. Behav. 2010, 57, 134–139. [Google Scholar] [CrossRef]
- Weintraub, R.M.; Churchill, W.H.; Crisler, C.; Rapp, H.J.; Borsos, T. Mouse complement: Influence of sex hormones on its activity. Science 1966, 152, 783–785. [Google Scholar] [CrossRef]
- Derzsy, Z.; Prohászka, Z.; Rigó, J.; Füst, G.; Molvarec, A. Activation of the complement system in normal pregnancy and preeclampsia. Mol. Immunol. 2010, 47, 1500–1506. [Google Scholar] [CrossRef]
- Richani, K.; Soto, E.; Romero, R.; Espinoza, J.; Chaiworapongsa, T.; Nien, J.K.; Edwin, S.; Kim, Y.M.; Hong, J.S.; Mazor, M. Normal pregnancy is characterized by systemic activation of the complement system. J. Matern. Neonatal Med. 2005, 17, 239–245. [Google Scholar] [CrossRef]
- Lochmiller, R.L.; Deerenberg, C. Trade-offs in evolutionary immunology: Just what is the cost of immunity? Oikos 2000, 88, 87–98. [Google Scholar] [CrossRef]
- Demas, G.E.; Greives, T.; Chester, E.; French, S. The Energetics of Immunity. In Ecoimmunology; Oxford University Press: New York, NY, USA, 2012; pp. 259–296. ISBN 9780199737345. [Google Scholar]
- Buehler, D.M.; Tieleman, B.I.; Piersma, T. How do migratory species stay healthy over the annual cycle? A conceptual model for immune function and for resistance to disease. Integr. Comp. Biol. 2010, 50, 346–357. [Google Scholar] [CrossRef]
- Simon, A.K.; Hollander, G.A.; Mcmichael, A. Evolution of the immune system in humans from infancy to old age. Proc. R. Soc. B 2015, 282, 20143085. [Google Scholar] [CrossRef]
- Sikes, R.S.; Gannon, W.L. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- Ceballos, G.; Fleming, T.H.; Chavez, C.; Nassar, J. Population dynamics of Leptonycteris curasoae (Chiroptera: Phyllostomidae) in Jalisco, Mexico. J. Mammal. 1997, 78, 1220–1230. [Google Scholar] [CrossRef]
- Anthony, E.L.P. Age Determination in Bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Ed.; Smithsonian Institution Press: Washington, DC, USA; London, UK, 1988; pp. 47–58. [Google Scholar]
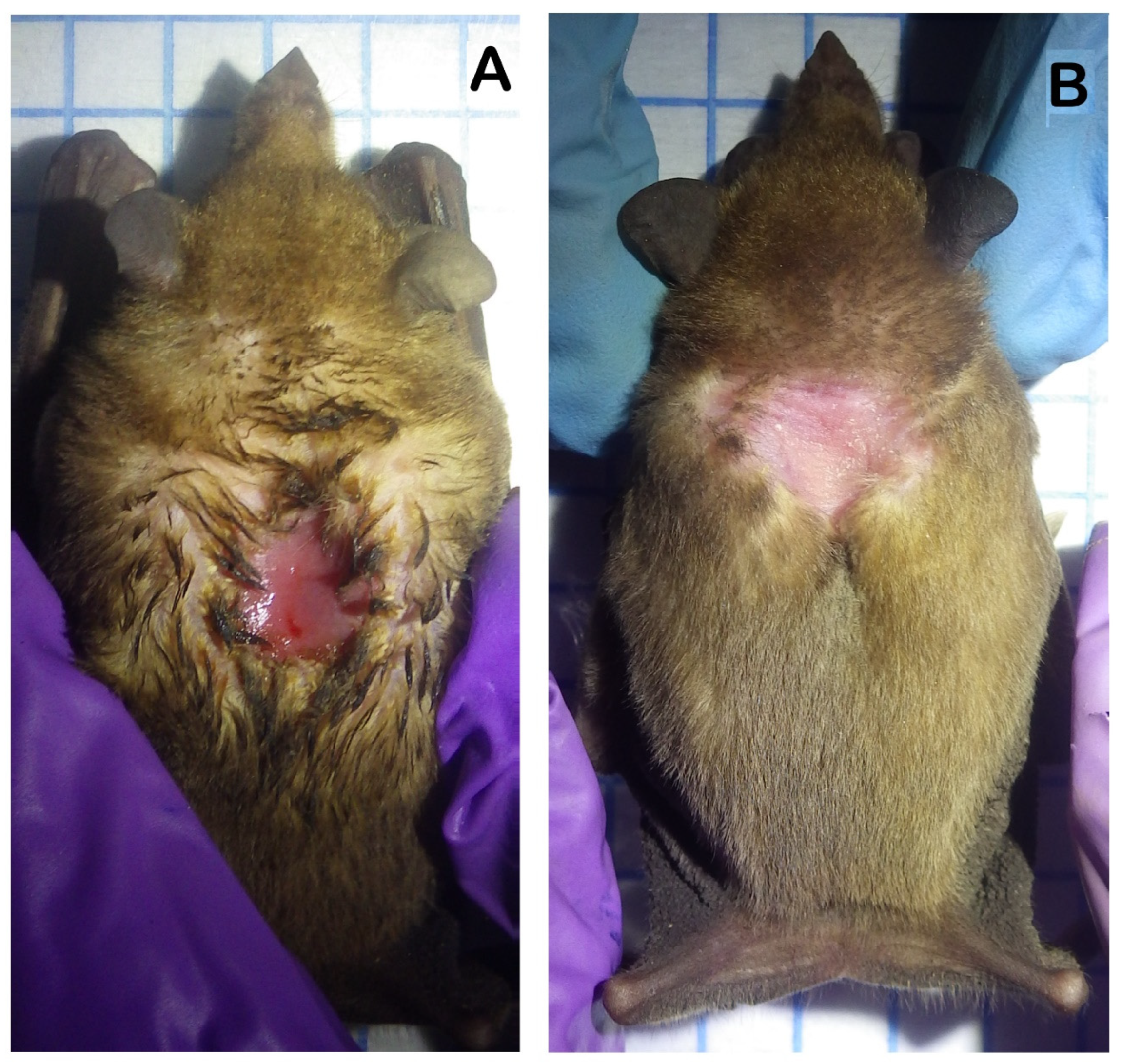
- Nassar, J.M.; Salazar, M.V.; Quintero, A.; Stoner, K.E.; Gómez, M.; Cabrera, A.; Jaffé, K. Seasonal sebaceous patch in the nectar-feeding bats Leptonycteris curasoae and L. yerbabuenae (Phyllostomidae: Glossophaginae): Phenological, histological, and preliminary chemical characterization. Zoology 2008, 111, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-romo, A.M.; Burgos, J.F.; Kunz, T.H. Smearing behaviour of male Leptonycteris curasoae (Chiroptera) and female responses to the odour of dorsal patches. Behaviour 2011, 148, 461–483. [Google Scholar]
- Rincón-Vargas, F.; Stoner, K.E.; Vigueras-Villaseñor, R.M.; Nassar, J.M.; Chaves, Ó.M.; Hudson, R. Internal and external indicators of male reproduction in the lesser long-nosed bat Leptonycteris yerbabuenae. J. Mammal. 2013, 94, 488–496. [Google Scholar] [CrossRef]
- Strobel, S.N.; Becker, J.A. No short-term effect of handling and capture stress on immune responses on bats assessed by bacterial killing assay. Mamm. Biol. 2015, 80, 312–315. [Google Scholar] [CrossRef]
- Becker, D.J.; Czirják, G.Á.; Rynda-apple, A.; Plowright, R.K. Handling stress and sample storage are associated with weaker complement-mediated bactericidal ability in birds but not bats. Physiol. Biochem. Zool. 2019, 92, 37–48. [Google Scholar] [CrossRef]
- Liebl, A.L.; Ii, L.B.M. Simple quantification of blood and plasma antimicrobial capacity using spectrophotometry. Funct. Ecol. 2009, 23, 1091–1096. [Google Scholar] [CrossRef]
- French, S.S.; Neuman-lee, L.A. Improved ex vivo method for microbiocidal activity across vertebrate species. Biol. Open 2012, 1, 482–487. [Google Scholar] [CrossRef]
- Schneeberger, K.; Courtiol, A.; Czirja Gabor, A.; Voigt, C.C. Immune profile predicts survival and reflects senescence in a small, long-lived mammal, the greater sac-winged bat (Saccopteryx bilineata). PLoS ONE 2014, 9, e108268. [Google Scholar] [CrossRef]
- Hernández-Arciga, U.; Herrera M., L.G.; Ibañez-Contreras, A.; Miranda-Labra, R.U.; Flores-Martínez, J.J.; Königsberg, M. Baseline and post-stress seasonal changes in immunocompetence and redox state maintenance in the fishing bat Myotis vivesi. PLoS ONE 2018, 13, e0190047. [Google Scholar] [CrossRef]
- Becker, D.J.; Chumchal, M.M.; Bentz, A.B.; Platt, S.G.; Czirják, G.; Rainwater, T.R.; Altizer, S.; Streicker, D.G. Predictors and immunological correlates of sublethal mercury exposure in vampire bats. R. Soc. Open Sci. 2017, 4, 170073. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; ISBN 9781544336466. [Google Scholar]
- Delignette-Muller, M.L.; Dutang, C. Fitdistrplus: An r package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biometr. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Miller, G.A.; Chapman, J.P. Misunderstanding analysis of covariance. J. Abnorm. Psychol. 2001, 110, 40–48. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R. Package Version 0.4.6. 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 17 January 2023).
- Ezenwa, V.O.; Ekernas, L.S.; Creel, S. Unravelling complex associations between testosterone and parasite infection in the wild. Funct. Ecol. 2012, 26, 123–133. [Google Scholar] [CrossRef]
- Walker, W.H. Testosterone signaling and the regulation of spermatogenesis. Spermatogenesis 2011, 1, 116–120. [Google Scholar] [CrossRef]
- Caren, L.D.; Rosenberg, L.T. Steroids and serum complement in mice: Influence of hydrocortisone, diethylstilbestrol, and testosterone. Science 1966, 152, 782–783. [Google Scholar] [CrossRef]
- Allen, L.C.; Turmelle, A.S.; Mendonça, M.T.; Navara, K.J.; Kunz, T.H.; McCracken, G.F. Roosting ecology and variation in adaptive and innate immune system function in the Brazilian free-tailed bat (Tadarida brasiliensis). J. Comp. Physiol. B 2009, 179, 315–323. [Google Scholar] [CrossRef]
- Nunn, C.L.; Gittleman, J.L.; Antonovics, J. Promiscuity and the primate immune system. Science 2000, 290, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Gaona, O.; Cerqueda-García, D.; Falcón, L.I.; Vázquez-Domínguez, G.; Valdespino-Castillo, P.M.; Neri-Barrios, C.X. Microbiota composition of the dorsal patch of reproductive male Leptonycteris yerbabuenae. PLoS ONE 2019, 14, e0226239. [Google Scholar] [CrossRef] [PubMed]
- Heesterbeek, D.A.C.; Angelier, M.L.; Harrison, R.A.; Rooijakkers, S.H.M. Complement and bacterial infections: From molecular mechanisms to therapeutic applications. J. Innate Immun. 2018, 10, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Bagherimoghaddam, A.; Rafatpanah, H.; Mansouritorghabeh, H. Elevated levels of C3, C4, and CH50 of the complement system in ICU and non-ICU patients with COVID-19. Health Sci. Rep. 2022, 5, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Romo, M.; Kunz, T.H. Dorsal patch and chemical signaling in males of the long-nosed bat, Leptonycteris curasoae (Chiroptera: Phyllostomidae). J. Mammal. 2009, 90, 1139–1147. [Google Scholar] [CrossRef]
- Muñoz-Romo, M.; Burgos, J.F.; Kunz, T.H. The dorsal patch of males of the Curaçaoan long-nosed bat, Leptonycteris curasoae (Phyllostomidae: Glossophaginae) as a visual signal. Acta Chiropt. 2011, 13, 207–215. [Google Scholar] [CrossRef]
- Nassar, J.M.; Galicia, R.; Ibarra, A.; Medellin, R.A. Tracking the origin of the smearing behavior in long-nosed bats (Leptonycteris spp.). Mamm. Biol. 2016, 81, 623–627. [Google Scholar] [CrossRef]
- Abbassy, M.; Shaheen, H.; Dykstra, E.; Beavers, G.; Hoel, D.; Hanafi, H.; Afifi, M.; Ibrahim, M.; Karim, K. Identification of Culex pipiens Linnaeus (Diptera: Culicidae) immunogens recognized by host humoral immunity and their impact on survival and fecundity. Egypt J. Immunol. 2007, 14, 43–54. [Google Scholar]
- Herrera M., L.G.; Hernández-Arciga, U.; González-Carcacía, J.A.; Nassar, J.M. Olfactory cues of mate quality in mammals: Inflammatory response is higher in male long-nosed bats with odorous dorsal patch. Mammal Res. in press.
- He, Y.; Xu, B.; Song, D.; Wang, Y.; Yu, F.; Chen, Q.; Zhao, M. Normal range of complement components during pregnancy: A prospective study. Am. J. Reprod. Immunol. 2020, 83, e13202. [Google Scholar] [CrossRef]
- Hopkinson, N.D.; Powell, R.J. Classical complement activation induced by pregnancy: Implications for management of connective tissue diseases. J. Clin. Pathol. 1992, 45, 66–67. [Google Scholar] [CrossRef]
- Cockrum, E.L. Seasonal distribution of northwestern populations of the long-nosed bats, Leptonycteris sanborni family Phyllostomidae. An. Inst. Biol. Ser. Zool. 1991, 62, 181–202. [Google Scholar]
- Valiente-Banuet, A.; Arizmendi, M.D.C.; Rojas-Martínez, A.; Domínguez-Canseco, L. Ecological relationships between columnar cacti and nectar-feeding bats in Mexico. J. Trop. Ecol. 1996, 12, 103–119. [Google Scholar] [CrossRef]
- Fleming, T.H.; Nuñez, R.A.; Sternberg, L.D.S.L. Seasonal changes in the diets of migrant and non-migrant nectarivorous bats as revealed by carbon stable isotope analysis. Oecologia 1993, 94, 72–75. [Google Scholar] [CrossRef]
- Burke, R.A.; Frey, J.K.; Ganguli, A.; Stoner, K.E. Species distribution modelling supports “nectar corridor” hypothesis for migratory nectarivorous bats and conservation of tropical dry forest. Divers. Distrib. 2019, 25, 1399–1415. [Google Scholar] [CrossRef]
- Gilot-fromont, E.; Jégo, M.; Bonenfant, C.; Gibert, P.; Rannou, B.; Klein, F.; Gaillard, J.-M. Immune phenotype and body condition in roe deer: Individuals with high body condition have different, not stronger immunity. PLoS ONE 2012, 7, e45576. [Google Scholar] [CrossRef]
- Hernández-Castellano, L.E.; Moreno-Indias, I.; Sánchez-Macías, D.; Morales-delaNuez, A.; Torres, A.; Argüello, A.; Castro, N. Sheep and goats raised in mixed flocks have diverse immune status around parturition. J. Dairy Sci. 2019, 102, 8478–8485. [Google Scholar] [CrossRef]
- Rojas-Martinez, A.; Valiente-Banuet, A.; Del Coro Arizmendi, M.; Alcántara-Eguren, A.; Arita, H.T. Seasonal distribution of the long-nosed bat (Leptonycteris curasoae) in North America: Does a generalized migration pattern really exist? J. Biogeogr. 1999, 26, 1065–1077. [Google Scholar] [CrossRef]
- Eikenaar, C.; Hegemann, A.; Packmor, F.; Kleudgen, I.; Isaksson, C. Not just fuel: Energy stores are correlated with immune function and oxidative damage in a long-distance migrant. Curr. Zool. 2020, 66, 21–28. [Google Scholar] [CrossRef]
- Friedrichs, V.; Toussaint, C.; Schäfer, A.; Rissmann, M.; Dietrich, O.; Mettenleiter, T.C.; Pei, G.; Balkema-Buschmann, A.; Saliba, A.E.; Dorhoi, A. Landscape and age dynamics of immune cells in the Egyptian rousette bat. Cell Rep. 2022, 40, 111305. [Google Scholar] [CrossRef]
- Aastrup, C.; Hegemann, A. Jackdaw nestlings rapidly increase innate immune function during the nestling phase but no evidence for a trade-off with growth. Dev. Comp. Immunol. 2021, 117, 103967. [Google Scholar] [CrossRef] [PubMed]
- Mauck, R.A.; Matson, K.D.; Philipsborn, J.; Ricklefs, R.E. Increase in the constitutive innate humoral immune system in leach’s storm-petrel (Oceanodroma leucorhoa) chicks is negatively correlated with growth rate. Funct. Ecol. 2005, 19, 1001–1007. [Google Scholar] [CrossRef]
- Cacho, N.T.; Lawrence, R.M. Innate immunity and breast milk. Front. Immunol. 2017, 8, 584. [Google Scholar] [CrossRef]
- Ingala, M.R.; Becker, D.J.; Bak Holm, J.; Kristiansen, K.; Simmons, N.B. Habitat fragmentation is associated with dietary shifts and microbiota variability in common vampire bats. Ecol. Evol. 2019, 9, 6508–6523. [Google Scholar] [CrossRef]
- Wilkinson, G.S.; Adams, D.M.; Haghani, A.; Lu, A.T.; Zoller, J.; Breeze, C.E.; Arnold, B.D.; Ball, H.C.; Carter, G.G.; Cooper, L.N.; et al. DNA methylation predicts age and provides insight into exceptional longevity of bats. Nat. Commun. 2021, 12, 1615. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age/Sex | Reproductive Traits | Site | Month/Year | BKA (%) | Body Mass (g) | n |
|---|---|---|---|---|---|---|
| Adult male | Non-reproductive | Jalisco | December 2019 | 16.41, 12.89–77.72 | 27.6 ± 5.7 | 3 |
| Adult female | Lactating | Jalisco | May 2019 | 47.99 | 29.0 | 1 |
| Adult female | Lactating | Jalisco | March 2020 | 24.28 | 25.9 | 1 |
| Adult female | Post-lactating | Jalisco | March 2020 | 12.77 | 23.8 | 1 |
| Adult female | Non-reproductive | Jalisco | May 2019 | 3.19, 99.63 | 24.0, 25.0 | 2 |
| Adult female | Non-reproductive | Jalisco | March 2020 | 94.05, 96.23 | 23.5, 27.0 | 2 |
| Adult female | Non-reproductive | Sonora | May 2019 | 96.66, 23.71–99.90 | 22.5 ± 1.0 | 3 |
| Adult female | Non-reproductive | Sonora | July 2019 | 0 | 27.5 | 1 |
| Adult female | Post-lactating | Sonora | July 2019 | 97.35, 23.71–99.90 | 27.2 ± 2.0 | 4 |
| Subadult male | Non-reproductive | Jalisco | March 2019 | 97.33, 52.23–99.90 | 23.3 ± 2.3 | 3 |
| Subadult male | Non-reproductive | Jalisco | December 2019 | 37.07 | 23.3 | 1 |
| Subadult male | Non-reproductive | Jalisco | March 2020 | 4.54, 0–71.43 | 23.4 ± 1.6 | 4 |
| Subadult female | Non-reproductive | Jalisco | December 2019 | 88.36, 74.32–99.90 | 23.8 ± 1.6 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Ruiz, D.A.; Flores-Martínez, J.J.; Rosales, C.; Herrera Montalvo, L.G. Constitutive Innate Immunity of Migrant and Resident Long-Nosed Bats (Leptonycteris yerbabuenae) in the Drylands of Mexico. Diversity 2023, 15, 530. https://doi.org/10.3390/d15040530
Rivera-Ruiz DA, Flores-Martínez JJ, Rosales C, Herrera Montalvo LG. Constitutive Innate Immunity of Migrant and Resident Long-Nosed Bats (Leptonycteris yerbabuenae) in the Drylands of Mexico. Diversity. 2023; 15(4):530. https://doi.org/10.3390/d15040530
Chicago/Turabian StyleRivera-Ruiz, David A., José Juan Flores-Martínez, Carlos Rosales, and Luis Gerardo Herrera Montalvo. 2023. "Constitutive Innate Immunity of Migrant and Resident Long-Nosed Bats (Leptonycteris yerbabuenae) in the Drylands of Mexico" Diversity 15, no. 4: 530. https://doi.org/10.3390/d15040530
APA StyleRivera-Ruiz, D. A., Flores-Martínez, J. J., Rosales, C., & Herrera Montalvo, L. G. (2023). Constitutive Innate Immunity of Migrant and Resident Long-Nosed Bats (Leptonycteris yerbabuenae) in the Drylands of Mexico. Diversity, 15(4), 530. https://doi.org/10.3390/d15040530