
Identification of Pseudo-nitzschia Cryptic Species Collected in the Gulf of Naples Using Whole-Cell Fluorescent In Situ Hybridization: From Cultured Sample to Field Test

 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Pseudo-nitzschia Cultivation
2.2. Whole-Cell Fluorescent In Situ Hybridization (WC-FISH) on Pseudo-nitzschia Species Monocultures—Probes Testing and Cross-Reactivity Assays
2.3. WC-FISH on Artificial Samples
2.4. WC-FISH on Environmental Samples
3. Results
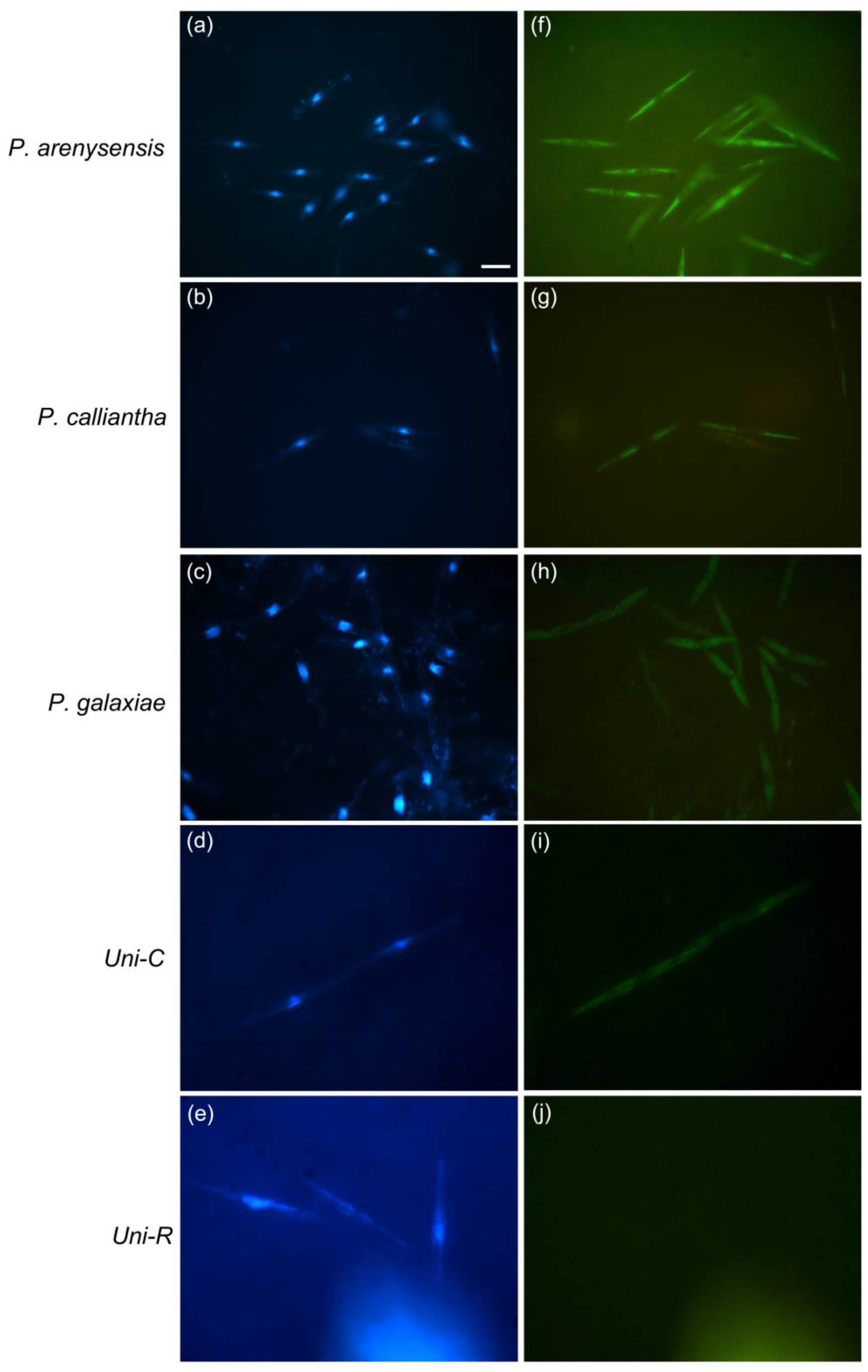
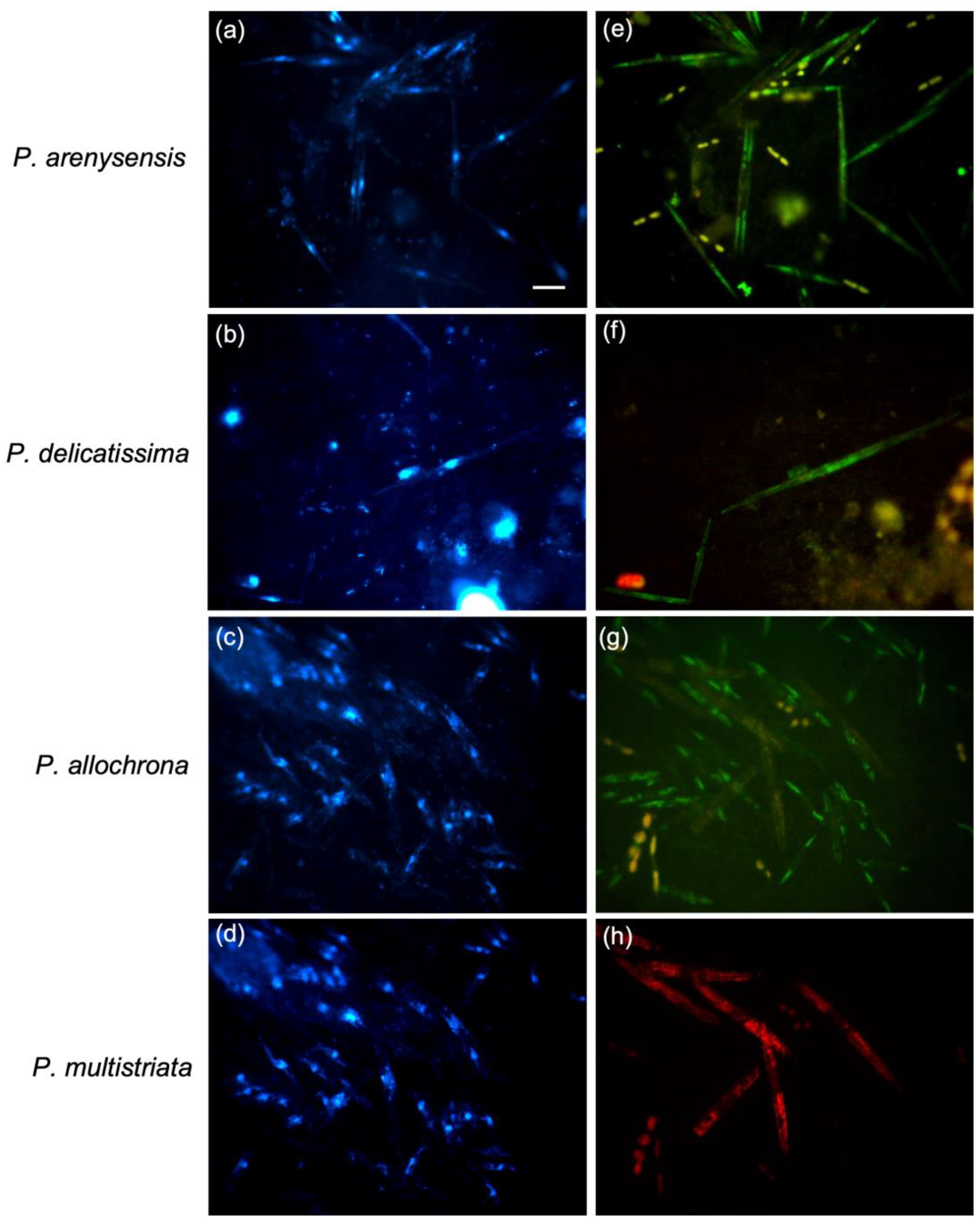
3.1. Probes Testing and Cross-Reactivity Trials
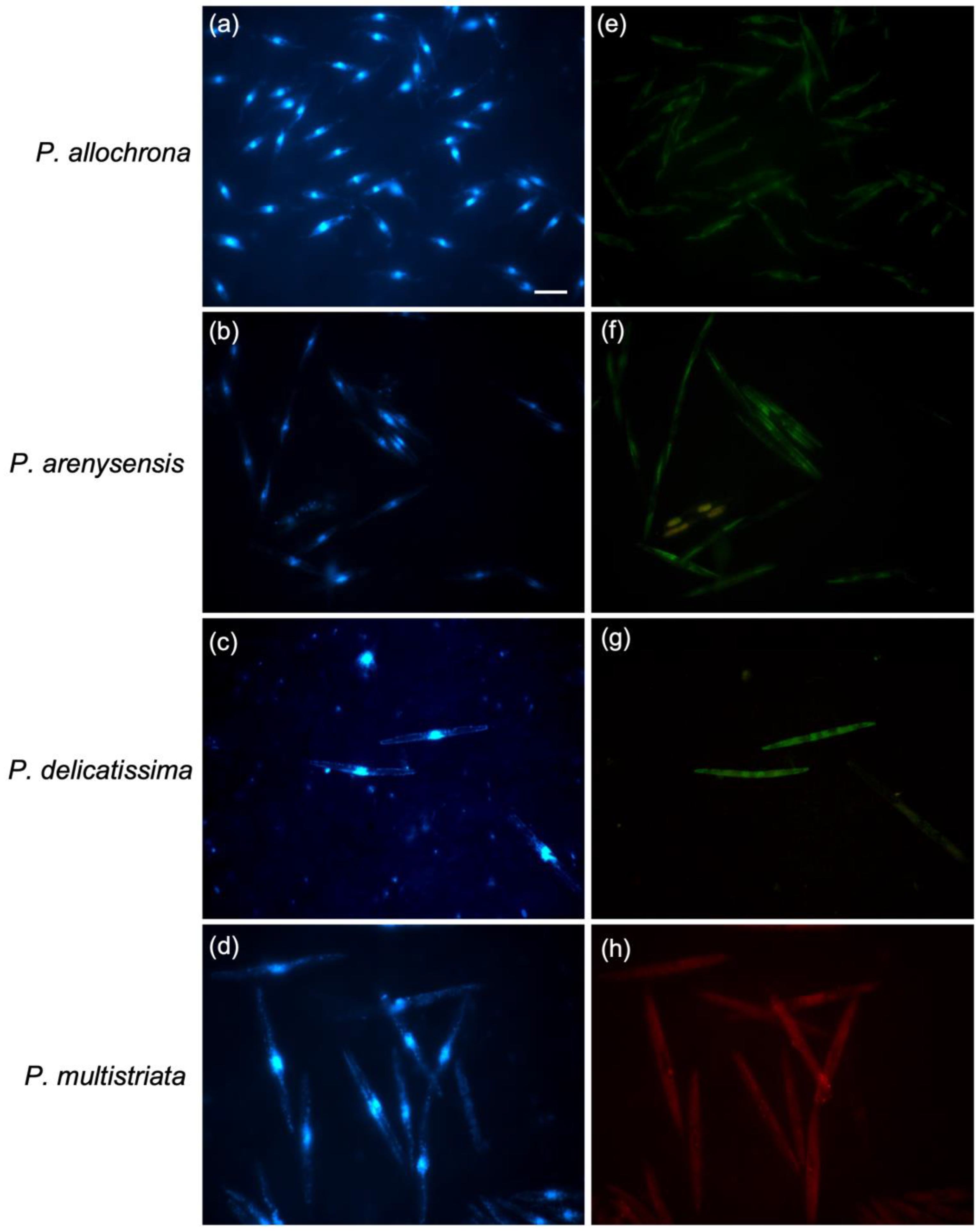
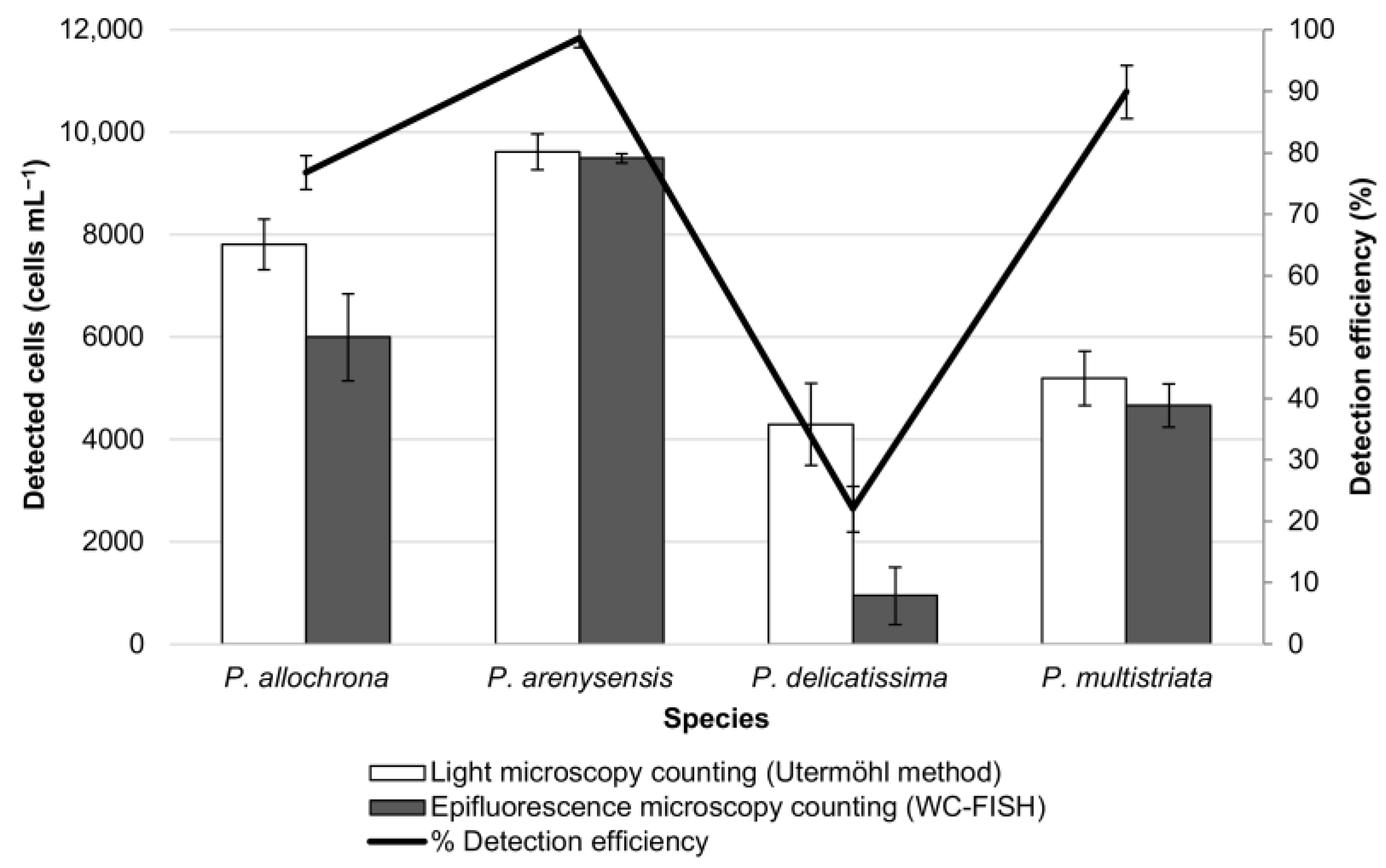
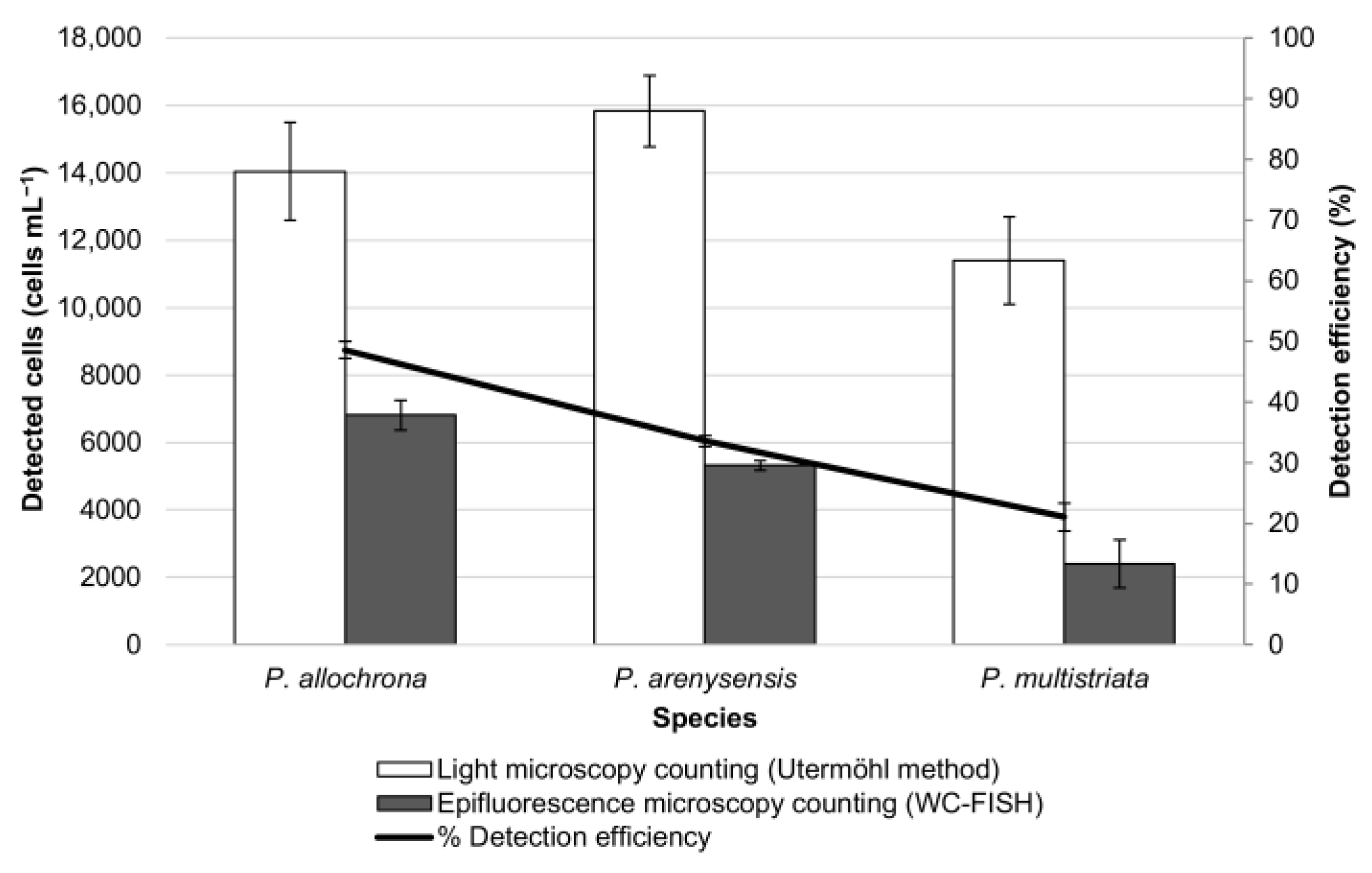
3.2. WC-FISH on Artificial Samples
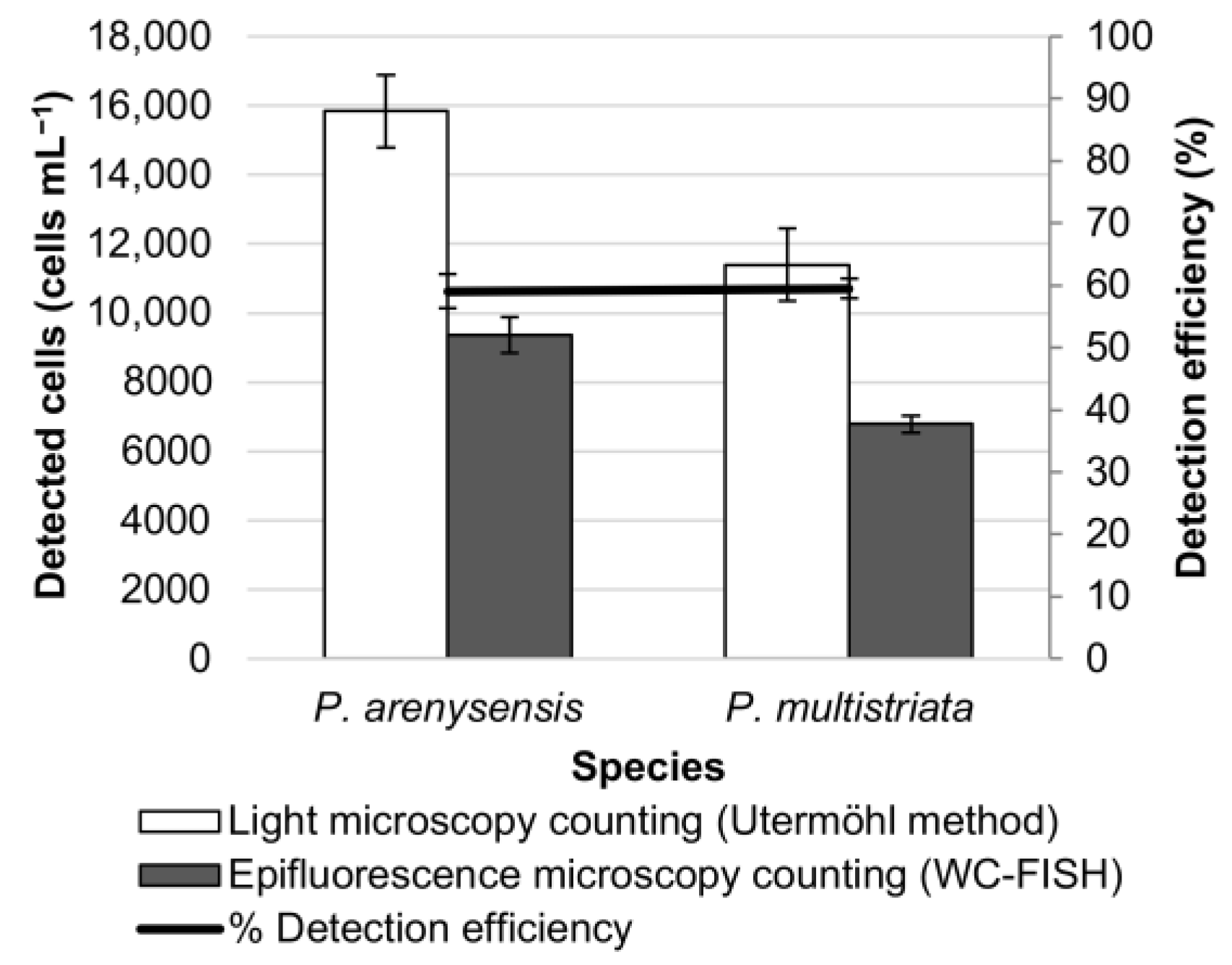
3.3. WC-FISH on Environmental Samples
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Moreno, A.R.; Anderson, C.; Kudela, R.M.; Sutula, M.; Edwards, C.; Bianchi, D. Development, Calibration, and Evaluation of a Model of Pseudo-nitzschia and Domoic Acid Production for Regional Ocean Modeling Studies. Harmful Algae 2022, 118, 102296. [Google Scholar] [CrossRef] [PubMed]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; DeLong, R.; De Vogelaere, A.; Harvey, J.; et al. Mortality of Sea Lions along the Central California Coast Linked to a Toxic Diatom Bloom. Nature 2000, 403, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Guillou, L.; Nézan, E.; Cueff, V.; Erard-Le Denn, E.; Cambon-Bonavita, M.-A.; Gentien, P.; Barbier, G. Genetic Diversity and Molecular Detection of Three Toxic Dinoflagellate Genera (Alexandrium, Dinophysis, and Karenia) from French Coasts. Protist 2002, 153, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Zabaglo, K.; Chrapusta, E.; Bober, B.; Kaminski, A.; Adamski, M.; Bialczyk, J. Environmental Roles and Biological Activity of Domoic Acid: A Review. Algal Res. 2016, 13, 94–101. [Google Scholar] [CrossRef]
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and Domoic Acid: New Research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef]
- Smodlaka Tanković, M.; Baričević, A.; Gerić, M.; Domijan, A.-M.; Pfannkuchen, D.M.; Kužat, N.; Ujević, I.; Kuralić, M.; Rožman, M.; Matković, K.; et al. Characterisation and Toxicological Activity of Three Different Pseudo-nitzschia Species from the Northern Adriatic Sea (Croatia). Environ. Res. 2022, 214, 114108. [Google Scholar] [CrossRef]
- Trainer, V.L.; Hickey, B.M.; Horner, R.A. Biological and Physical Dynamics of Domoic Acid Production off the Washington Coast. Limnol. Oceanogr. 2002, 47, 1438–1446. [Google Scholar] [CrossRef]
- Orsini, L.; Procaccini, G.; Sarno, D.; Montresor, M. Multiple RDNA ITS-Types within the Diatom Pseudo-nitzschia Delicatissima (Bacillariophyceae) and Their Relative Abundances across a Spring Bloom in the Gulf of Naples. Mar. Ecol. Prog. Ser. 2004, 271, 87–98. [Google Scholar] [CrossRef]
- Sahraoui, I.; Bates, S.S.; Bouchouicha, D.; Mabrouk, H.H.; Hlaili, A.S. Toxicity of Pseudo-nitzschia Populations from Bizerte Lagoon, Tunisia, Southwest Mediterranean, and First Report of Domoic Acid Production by P. Brasiliana. Diatom Res. 2011, 26, 293–303. [Google Scholar] [CrossRef]
- Lundholm, N.; Bates, S.S.; Baugh, K.A.; Bill, B.D.; Connell, L.B.; Léger, C.; Trainer, V.L. Cryptic and pseudo-cryptic diversity in diatoms—with descriptions of Pseudo-nitzschia hasleana sp. nov. and P. fryxelliana sp. nov. J. Phycol. 2012, 48, 436–454. [Google Scholar] [CrossRef]
- Lundholm, N.; Moestrup, Ø.; Kotaki, Y.; Hoef-Emden, K.; Scholin, C.; Miller, P. Inter-and intraspecific variation of the Pseudo-nitzschia delicatissima complex (Bacillariophyceae) illustrated by rRNA probes, morphological data and phylogenetic analyses. J. Phycol. 2006, 42, 464–481. [Google Scholar] [CrossRef]
- Ruggiero, M.V.; Sarno, D.; Barra, L.; Kooistra, W.H.C.F.; Montresor, M.; Zingone, A. Diversity and Temporal Pattern of Pseudo-nitzschia Species (Bacillariophyceae) through the Molecular Lens. Harmful Algae 2015, 42, 15–24. [Google Scholar] [CrossRef]
- Lim, H.C.; Teng, S.T.; Leaw, C.P.; Lim, P.T. Three Novel Species in the Pseudo-nitzschia Pseudodelicatissima Complex: P. Batesiana Sp. Nov., P. Lundholmiae Sp. Nov., and P. Fukuyoi Sp. Nov. (Bacillariophyceae) from the Strait of Malacca, Malaysia. J. Phycol. 2013, 49, 902–916. [Google Scholar] [CrossRef]
- Percopo, I.; Ruggiero, M.V.; Balzano, S.; Gourvil, P.; Lundholm, N.; Siano, R.; Tammilehto, A.; Vaulot, D.; Sarno, D. Pseudo-nitzschia Arctica Sp. Nov., a New Cold-Water Cryptic Pseudo-nitzschia Species within the P. pseudodelicatissima Complex. J. Phycol. 2016, 52, 184–199. [Google Scholar] [CrossRef]
- Quijano-Scheggia, S.I.; Garcés, E.; Lundholm, N.; Moestrup, Ø.; Andree, K.; Camp, J. Morphology, Physiology, Molecular Phylogeny and Sexual Compatibility of the Cryptic Pseudo-nitzschia Delicatissima Complex (Bacillariophyta), Including the Description of P. Arenysensis Sp. Nov. Phycologia 2009, 48, 492–509. [Google Scholar] [CrossRef]
- Ruggiero, M.V.; Kooistra, W.H.C.F.; Piredda, R.; Sarno, D.; Zampicinini, G.; Zingone, A.; Montresor, M. Temporal Changes of Genetic Structure and Diversity in a Marine Diatom Genus Discovered via Metabarcoding. Environ. DNA 2022, 4, 763–775. [Google Scholar] [CrossRef]
- Percopo, I.; Ruggiero, M.V.; Sarno, D.; Longobardi, L.; Rossi, R.; Piredda, R.; Zingone, A. Phenological Segregation Suggests Speciation by Time in the Planktonic Diatom Pseudo-nitzschia Allochrona Sp. Nov. Ecol. Evol. 2022, 12, e9155. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, K.A.; Rocap, G.; Armbrust, E.V. Inter-and intraspecific community structure within the diatom genus Pseudo-nitzschia (Bacillariophyceae). J. Phycol. 2008, 44, 637–649. [Google Scholar] [CrossRef]
- Galluzzi, L.; Cegna, A.; Casabianca, S.; Penna, A.; Saunders, N.; Magnani, M. Development of an Oligonucleotide Microarray for the Detection and Monitoring of Marine Dinoflagellates. J. Microbiol. Methods 2011, 84, 234–242. [Google Scholar] [CrossRef]
- Andree, K.B.; Fernández-Tejedor, M.; Elandaloussi, L.M.; Quijano-Scheggia, S.; Sampedro, N.; Garcés, E.; Camp, J.; Diogène, J. Quantitative PCR Coupled with Melt Curve Analysis for Detection of Selected Pseudo-nitzschia Spp. (Bacillariophyceae) from the Northwestern Mediterranean Sea. Appl. Environ. Microbiol. 2011, 77, 1651–1659. [Google Scholar] [CrossRef]
- Barra, L.; Ruggiero, M.V.; Sarno, D.; Montresor, M.; Kooistra, W.H.C.F. Strengths and Weaknesses of Microarray Approaches to Detect Pseudo-nitzschia Species in the Field. Environ. Sci. Pollut. Res. 2013, 20, 6705–6718. [Google Scholar] [CrossRef] [PubMed]
- Edvardsen, B.; Dittami, S.M.; Groben, R.; Brubak, S.; Escalera, L.; Rodríguez, F.; Reguera, B.; Chen, J.; Medlin, L.K. Molecular Probes and Microarrays for the Detection of Toxic Algae in the Genera Dinophysis and Phalacroma (Dinophyta). Environ. Sci. Pollut. Res. 2013, 20, 6733–6750. [Google Scholar] [CrossRef]
- Kegel, J.U.; Del Amo, Y.; Medlin, L.K. Introduction to Project MIDTAL: Its Methods and Samples from Arcachon Bay, France. Environ. Sci. Pollut. Res. 2013, 20, 6690–6704. [Google Scholar] [CrossRef]
- McCoy, G.R.; Touzet, N.; Fleming, G.T.; Raine, R. An Evaluation of the Applicability of Microarrays for Monitoring Toxic Algae in Irish Coastal Waters. Environ. Sci. Pollut. Res. 2013, 20, 6751–6764. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.E.; Scholin, C.A. Identification and enumeration of cultured and wild Pseudo-nitzschia (Bacillariophyceae) using species-specific LSU rRNA-targeted fluorescent probes and filter-based whole cell hybridization. J. Phycol. 1998, 34, 371–382. [Google Scholar] [CrossRef]
- Parsons, M.L.; Scholin, C.A.; Miller, P.E.; Doucette, G.J.; Powell, C.L.; Fryxell, G.A.; Dortch, Q.; Soniat, T.M. Pseudo-nitzschia species (Bacillariophyceae) in Louisiana coastal waters: Molecular probe field trials, genetic variability, and domoic acid analyses. J. Phycol. 1999, 35, 1368–1378. [Google Scholar] [CrossRef]
- Sako, Y.; Hosoi-Tanabe, S.; Uchida, A. fluorescence in situ hybridization using rRNA-targeted probes for simple and rapid identification of the toxic dinoflagellates Alexandrium tamarense and Alexandrium catenella. J. Phycol. 2004, 40, 598–605. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Keafer, B.A.; Gribble, K.E.; Marin, R.; Scholin, C.A. Identification and Enumeration of Alexandrium Spp. from the Gulf of Maine Using Molecular Probes. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 2467–2490. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, C.; Zhang, B.; Wang, G.; Lu, D.; Xu, Z.; Yan, P. Development of a PNA Probe for Fluorescence In Situ Hybridization Detection of Prorocentrum Donghaiense. PLoS ONE 2011, 6, e25527. [Google Scholar] [CrossRef]
- Caracciolo, A.B.; Dejana, L.; Fajardo, C.; Grenni, P.; Martin, M.; Mengs, G.; Sánchez-Fortún, S.; Lettieri, T.; Saccà, M.L.; Medlin, L.K. A New Fluorescent Oligonucleotide Probe for In-Situ Identification of Microcystis Aeruginosa in Freshwater. Microchem. J. 2019, 148, 503–513. [Google Scholar] [CrossRef]
- Morais, S.L.; Barros, P.; Santos, M.; Delerue-Matos, C.; Gomes, A.C.; Fátima Barroso, M. Electrochemical Genosensor for the Detection of Alexandrium Minutum Dinoflagellates. Talanta 2021, 222, 121416. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, C.; Wang, Y.; Chen, G. A Review of the Current and Emerging Detection Methods of Marine Harmful Microalgae. Sci. Total Environ. 2022, 815, 152913. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.K.; Orozco, J. Molecular Techniques for the Detection of Organisms in Aquatic Environments, with Emphasis on Harmful Algal Bloom Species. Sensors 2017, 17, 1184. [Google Scholar] [CrossRef] [PubMed]
- Toebe, K. Whole Cell Hybridisation for Monitoring Harmful Marine Microalgae. Environ. Sci. Pollut. Res. 2013, 20, 6816–6823. [Google Scholar] [CrossRef]
- McDonald, S.M.; Sarno, D.; Scanlan, D.J.; Zingone, A. Genetic Diversity of Eukaryotic Ultraphytoplankton in the Gulf of Naples during an Annual Cycle. Aquat. Microb. Ecol. 2007, 50, 75–89. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals: Proceedings —1st Conference on Culture of Marine Invertebrate Animals Greenport; Smith, W.L., Chanley, M.H., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. ISBN 978-1-4615-8714-9. [Google Scholar]
- Zingone, A.; Totti, C.; Sarno, D.; Cabrini, M.; Caroppo, C.; Giacobbe, M.; Lugliè, A.; Nuccio, C.; Socal, G. Fitoplancton: Metodiche Di Analisi Quali-Quantitativa. In Metodologie di Studio del Plancton Marino. Manuali e Linee Guida 56/2010; ISPRA: Roma, Italy, 2010; pp. 213–237. ISBN 978-88-448-0427-5. [Google Scholar]
- Miller, P.E.; Scholin, C.A. On Detection of Pseudo-nitzschia (Bacillariophyceae) Species Using Whole Cell Hybridization: Sample Fixation and Stability. J. Phycol. 2000, 36, 238–250. [Google Scholar] [CrossRef]
- Groben, R.; Medlin, L. In Situ Hybridization of Phytoplankton Using Fluorescently Labeled RRNA Probes. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2005; Volume 395, pp. 299–310. ISBN 0076-6879. [Google Scholar]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Barra, L.; Ruggiero, M.V.; Chen, J.; Kooistra, W.H.C.F. Specificity of LSU rRNA-Targeted Oligonucleotide Probes for Pseudo-nitzschia Species Tested through Dot-Blot Hybridisation. Environ. Sci. Pollut. Res. 2013, 21, 548–557. [Google Scholar] [CrossRef]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar; Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; et al. ARB: A Software Environment for Sequence Data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.Y. The Errors of the Sedgwick-Rafter Counting Chamber in the Enumeration of Phytoplankton. Trans. Am. Microsc. Soc. 1942, 61, 217–226. [Google Scholar] [CrossRef]
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The Inverted Microscope Method of Estimating Algal Numbers and the Statistical Basis of Estimations by Counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- DeLong, E.; Wickham, G.; Pace, N. Phylogenetic Stains: Ribosomal RNA-Based Probes for the Identification of Single Cells. Science 1989, 243, 1360–1363. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Keafer, B.A.; Berdalet, E. Detection of the Toxic Dinoflagellate Alexandrium Fundyense (Dinophyceae) with Oligonucleotide and Antibody Probes: Variability in Labeling Intensity with Physiological Condition. J. Phycol. 1999, 35, 870–883. [Google Scholar] [CrossRef]
- Dittami, S.M.; Edvardsen, B. Culture conditions influence cellular RNA content in ichthyotoxic flagellates of the genus Pseudochattonella (Dictyochophyceae). J. Phycol. 2012, 48, 1050–1055. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.K.; Kegel, J.U. Validation of the Detection of Pseudo-nitzschia Spp. Using Specific RNA Probes Tested in a Microarray Format: Calibration of Signal Based on Variability of RNA Content with Environmental Conditions. Harmful Algae 2014, 37, 183–193. [Google Scholar] [CrossRef]
- Wright, E.S.; Yilmaz, L.S.; Corcoran, A.M.; Ökten, H.E.; Noguera, D.R. Automated Design of Probes for RRNA-Targeted Fluorescence In Situ Hybridization Reveals the Advantages of Using Dual Probes for Accurate Identification. Appl. Environ. Microbiol. 2014, 80, 5124–5133. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Ono, N.; Furusawa, C.; Kashiwagi, A.; Yomo, T. Experimental Optimization of Probe Length to Increase the Sequence Specificity of High-Density Oligonucleotide Microarrays. BMC Genom. 2007, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Metfies, K.; Medlin, L.K. Feasibility of Transferring Fluorescent in Situ Hybridization Probes to an 18S RRNA Gene Phylochip and Mapping of Signal Intensities. Appl. Environ. Microbiol. 2008, 74, 2814–2821. [Google Scholar] [CrossRef] [PubMed]
- Behrens, S.; Fuchs, B.M.; Mueller, F.; Amann, R. Is the In Situ Accessibility of the 16S RRNA of Escherichia Coli for Cy3-Labeled Oligonucleotide Probes Predicted by a Three-Dimensional Structure Model of the 30S Ribosomal Subunit? Appl. Environ. Microbiol. 2003, 69, 4935–4941. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Probe a | Sequence (5′-3′) | Target Species | Target Gene | Tm (°C) |
|---|---|---|---|---|---|
| P. delicatissima | Pdel4D02_25 | GATTGTGCAAATATCCAACCACTGT | P. allochrona | 28S | 66.6 |
| Pdel4D03_25 | TGACAACGACTCACTCTACCAGGC | P. allochrona | 28S | 69.4 | |
| Pdel3D01_25 | GACAAAAACTCACTCTACCAGGCGG | P. arenysensis | 28S | 69.5 | |
| Pdel3D02_25 | TAATGTTAAAGTCTATAGACCACAA | P. arenysensis | 28S | 55.5 | |
| Pdel2D01_25 | TCCAACCACTGTTACTTTCATTACG | P. delicatissima | 28S | 65.6 | |
| Pdel1D01_25 | TTGACAACGACTCACTCCACCAGG | P. dolorosa | 28S | 71.5 | |
| P. pseudodelicatissima | Pman2D03_25 | CTTCAGACCACAATTCGGCGCTTAAA | P. calliantha | 28S | 65.6 |
| PpdelD02_25 | CCCGGCAGATAACGTCAAGGTCTAT | P. pseudodelicatissima | 28S | 70.4 | |
| PNFRAGA | ATTCCACCCAAACATGGC | Pseudo-nitzschia spp. | 18S | 63.3 | |
| PfrauD04_25 | ACGGGAGTTTCACCCTCTCAGCTGTC | P. fraudulenta | 28S | 66.3 | |
| PgalaD02_25 | CCAAAGGAATCAACCAAAGCAAACC | P. galaxiae | 28S | 71.8 | |
| PmulaD03_25 | AACCCAAACTCACGAAAGCTCACAG | P. multistriata | 28S | 69.8 | |
| Controls a | Uni-C | GWATTACCGCGGKGCTG | Eukaryotic | 18S | 64.6 |
| Uni-R | CAGCMGCCGCGGUAAUWC | Prokaryotic | 16S | 60.6 |
| Probe | Target Species | Hybridization T (°C) | FA (%) | Washing T (°C) | Washing Buffer Concentration | Epifluorescent Signal |
|---|---|---|---|---|---|---|
| Pdel4D02_25 | P. allochrona | 45 | 40 | 55 | 0.5× | ++ |
| Pdel4D03_25 | P. allochrona | 45 | 40 | 55 | 0.5× | ++ |
| Pdel3D01_25 | P. arenysensis | 45 | 40 | 55 | 0.5× | ++ |
| Pdel3D02_25 | P. arenysensis | 45 | 40 | 55 | 0.5× | ++ |
| Pdel2D01_25 | P. delicatissima | 45 | 40 | 55 | 0.5× | ++ |
| Pdel1D01_25 | P. dolorosa | 45 | 40 | 55 | 0.5× | ++ |
| Pman2D03_25 | P. calliantha | 45 | 40 | 55 | 0.5× | ++ |
| PpdelD02_25 | P. pseudodelicatissima | 45 | 40 | 50 | 0.5× | ++ |
| PfrauD04_25 | P. fraudulenta | 45 | 40 | 55 | 2× | ++ |
| PgalaD02_25 | P. galaxiae | 45 | 40 | 55 | 2× | ++ |
| PmulaD03_25 | P. multistriata | 45 | 40 | 55 | 0.5× | ++ |
| Species | Probe | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pdel4D03_25 | Pdel3D01_25 | Pdel2D01_25 | Pdel1D01_25 | Pman2D03_25 | PpdelD02_25 | PfrauD04_25 | PgalaD02_25 | PmulaD03_25 | Uni-C | Uni-R | |
| P. allochrona | ++ | – | – | – | – | – | – | – | – | ++ | – |
| P. arenysensis | – | ++ | – | – | – | – | + | – | – | ++ | – |
| P. delicatissima | – | – | ++ | – | – | – | – | – | – | ++ | – |
| P. dolorosa | – | – | – | ++ | – | – | + | – | – | ++ | – |
| P. calliantha | – | – | – | – | ++ | + | – | + | – | ++ | – |
| P. pseudodelicatissima | – | – | – | – | + | ++ | – | + | – | ++ | – |
| P. fraudulenta | – | + | – | + | – | – | ++ | – | – | ++ | – |
| P. galaxiae | – | – | – | – | + | + | – | ++ | – | ++ | – |
| P. multistriata | – | – | – | – | – | – | – | – | ++ | ++ | – |
| Probe | Target Species | Hybridization T (°C) | FA (%) | Washing T (°C) | Washing Buffer Concentration |
|---|---|---|---|---|---|
| Pdel4D02_25 | P. allochrona | 45 | 40 | 55 | 2× |
| Pdel3D01_25 | P. arenysensis | 45 | 40 | 58 | 0.5× |
| Pdel2D01_25 | P. delicatissima | 45 | 40 | 45 | 5× |
| Pdel1D01_25 | P. dolorosa | 45 | 40 | 58 | 0.5× |
| Pman2D03_25 | P. calliantha | 45 | 40 | 55 | 0.5× |
| PpdelD02_25 | P. pseudodelicatissima | 45 | 40 | 55 | 2× |
| PfrauD04_25 | P. fraudulenta | 45 | 40 | 55 | 2× |
| PgalaD02_25 | P. galaxiae | 45 | 40 | 58 | 5× |
| PmulaD03_25 | P. multistriata | 45 | 40 | 58 | 2× |
| Species | Probe | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pdel4D03_25 | Pdel3D01_25 | Pdel2D01_25 | Pdel1D01_25 | Pman2D03_25 | PpdelD02_25 | PfrauD04_25 | PgalaD02_25 | PmulaD03_25 | Uni-C | Uni-R | |
| P. allochrona | ++ | – | – | – | – | – | – | – | – | ++ | – |
| P. arenysensis | – | ++ | – | – | – | – | – | – | – | ++ | – |
| P. delicatissima | – | – | ++ | – | – | – | – | – | – | ++ | – |
| P. dolorosa | – | – | – | ++ | – | – | – | – | – | ++ | – |
| P. calliantha | – | – | – | – | ++ | + | – | + | – | ++ | – |
| P. pseudodelicatissima | – | – | – | – | + | ++ | – | + | – | ++ | – |
| P. fraudulenta | – | – | – | – | – | – | ++ | – | – | ++ | – |
| P. galaxiae | – | – | – | – | + | + | – | ++ | – | ++ | – |
| P. multistriata | – | – | – | – | – | – | – | – | ++ | ++ | – |
| Sampling Date | Light Microscopy Counting (Utermöhl Method) (Cells mL−1) | Epifluorescence Microscopy Counting (WC-FISH) (Cells mL−1) | % Detection Efficiency | ||||
|---|---|---|---|---|---|---|---|
| P. delicatissima complex | P. allochrona | P. arenysensis | P. delicatissima | P. dolorosa | Total cryptic species | ||
| May | 210,490 ± 12,735 | 6900 ± 434 | 3900 ± 273 | 12,100 ± 726 | - | 22,900 ± 1433 | 10.9 |
| June | 16,498 ± 989 | - | - | 14,400 ± 864 | - | 14,400 ± 864 | 87.3 |
| July | 33,235 ± 2126 | - | - | 29,600 ± 1776 | - | 29,600 ± 1776 | 89.1 |
| September | 288,038 ± 17,162 | 3800 ± 252 | 7000 ± 470 | 28,200 ± 1792 | 2200 ± 153 | 41,200 ± 2667 | 14.3 |
| February | - | - | - | - | - | - | - |
| March | 177,855 ± 10,749 | - | 13,517 ± 821 | 4091 ± 275 | - | 17,608 ± 1096 | 10 |
| April | 686,861 ± 41,680 | - | 103,029 ± 6181 | 61,130 ± 3767 | - | 164,159 ± 9948 | 23.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrari, M.; Barra, L.; Ruffolo, L.; Muto, A.; Galasso, C.; Percopo, I.; Greco, S.; Cozza, R. Identification of Pseudo-nitzschia Cryptic Species Collected in the Gulf of Naples Using Whole-Cell Fluorescent In Situ Hybridization: From Cultured Sample to Field Test. Diversity 2023, 15, 521. https://doi.org/10.3390/d15040521
Ferrari M, Barra L, Ruffolo L, Muto A, Galasso C, Percopo I, Greco S, Cozza R. Identification of Pseudo-nitzschia Cryptic Species Collected in the Gulf of Naples Using Whole-Cell Fluorescent In Situ Hybridization: From Cultured Sample to Field Test. Diversity. 2023; 15(4):521. https://doi.org/10.3390/d15040521
Chicago/Turabian StyleFerrari, Michele, Lucia Barra, Luisa Ruffolo, Antonella Muto, Christian Galasso, Isabella Percopo, Silvestro Greco, and Radiana Cozza. 2023. "Identification of Pseudo-nitzschia Cryptic Species Collected in the Gulf of Naples Using Whole-Cell Fluorescent In Situ Hybridization: From Cultured Sample to Field Test" Diversity 15, no. 4: 521. https://doi.org/10.3390/d15040521
APA StyleFerrari, M., Barra, L., Ruffolo, L., Muto, A., Galasso, C., Percopo, I., Greco, S., & Cozza, R. (2023). Identification of Pseudo-nitzschia Cryptic Species Collected in the Gulf of Naples Using Whole-Cell Fluorescent In Situ Hybridization: From Cultured Sample to Field Test. Diversity, 15(4), 521. https://doi.org/10.3390/d15040521