
Invasion Potential of Ornamental Terrestrial Gastropods in Europe Based on Climate Matching


Abstract
1. Introduction
2. Materials and Methods
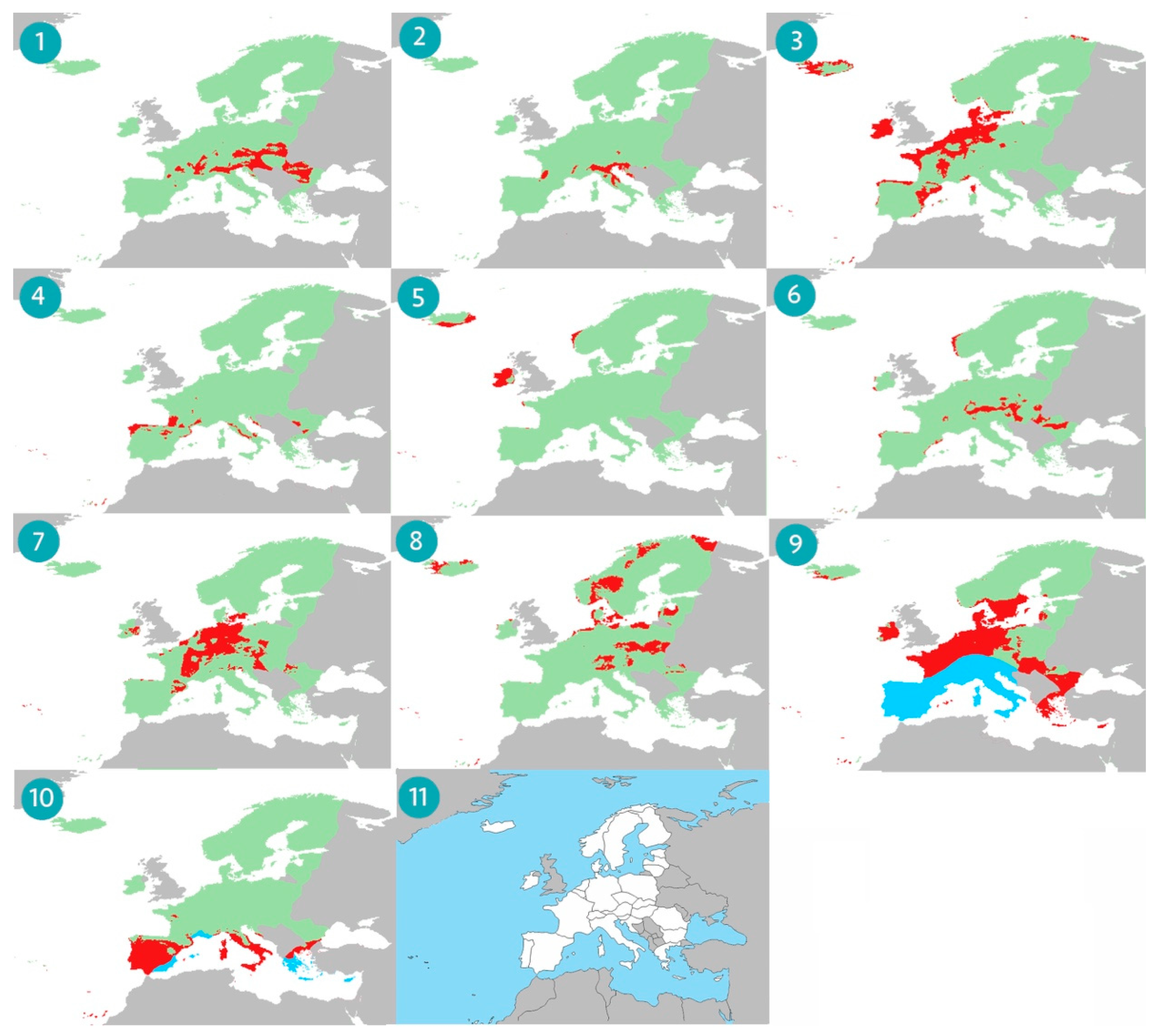
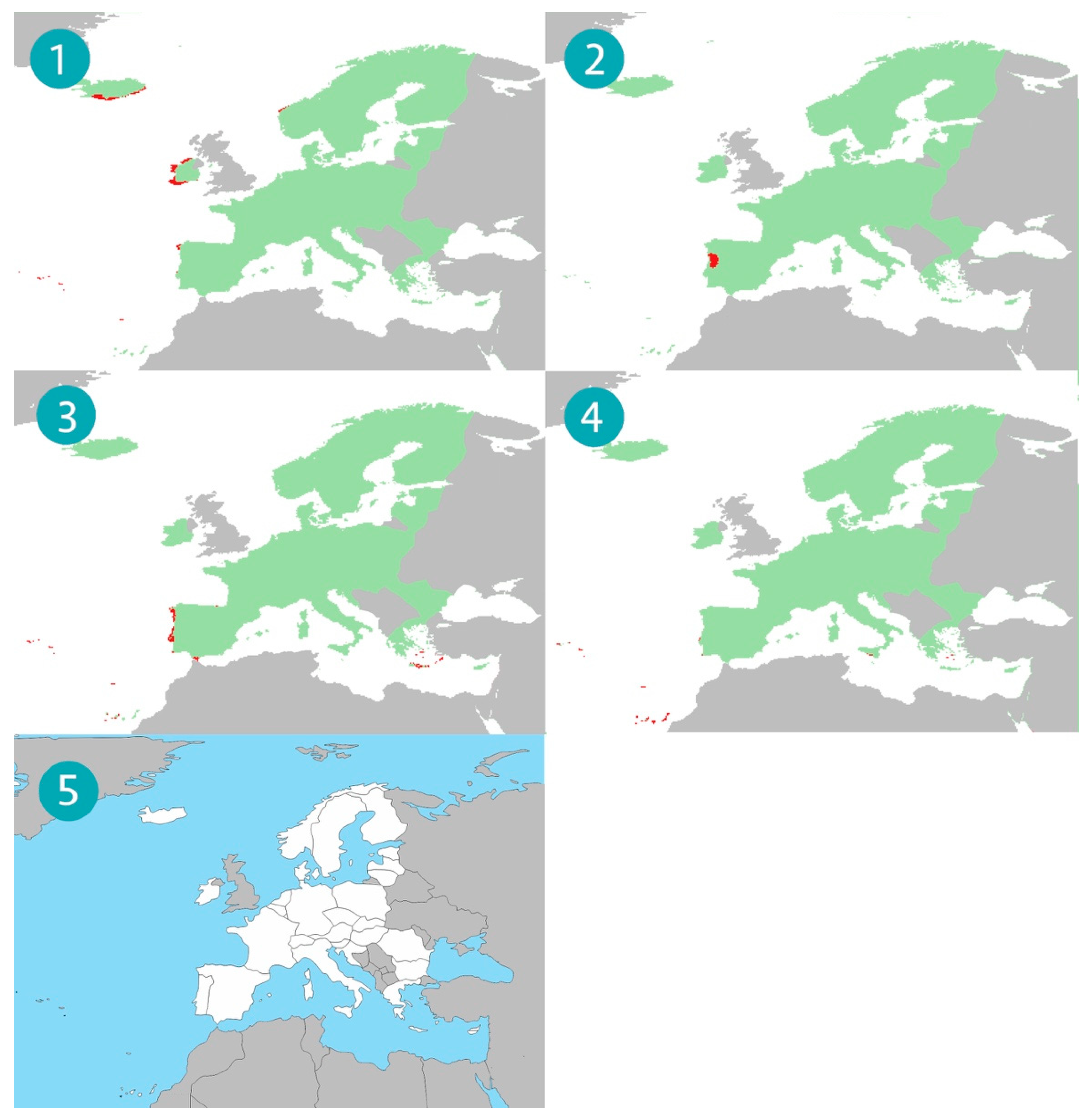
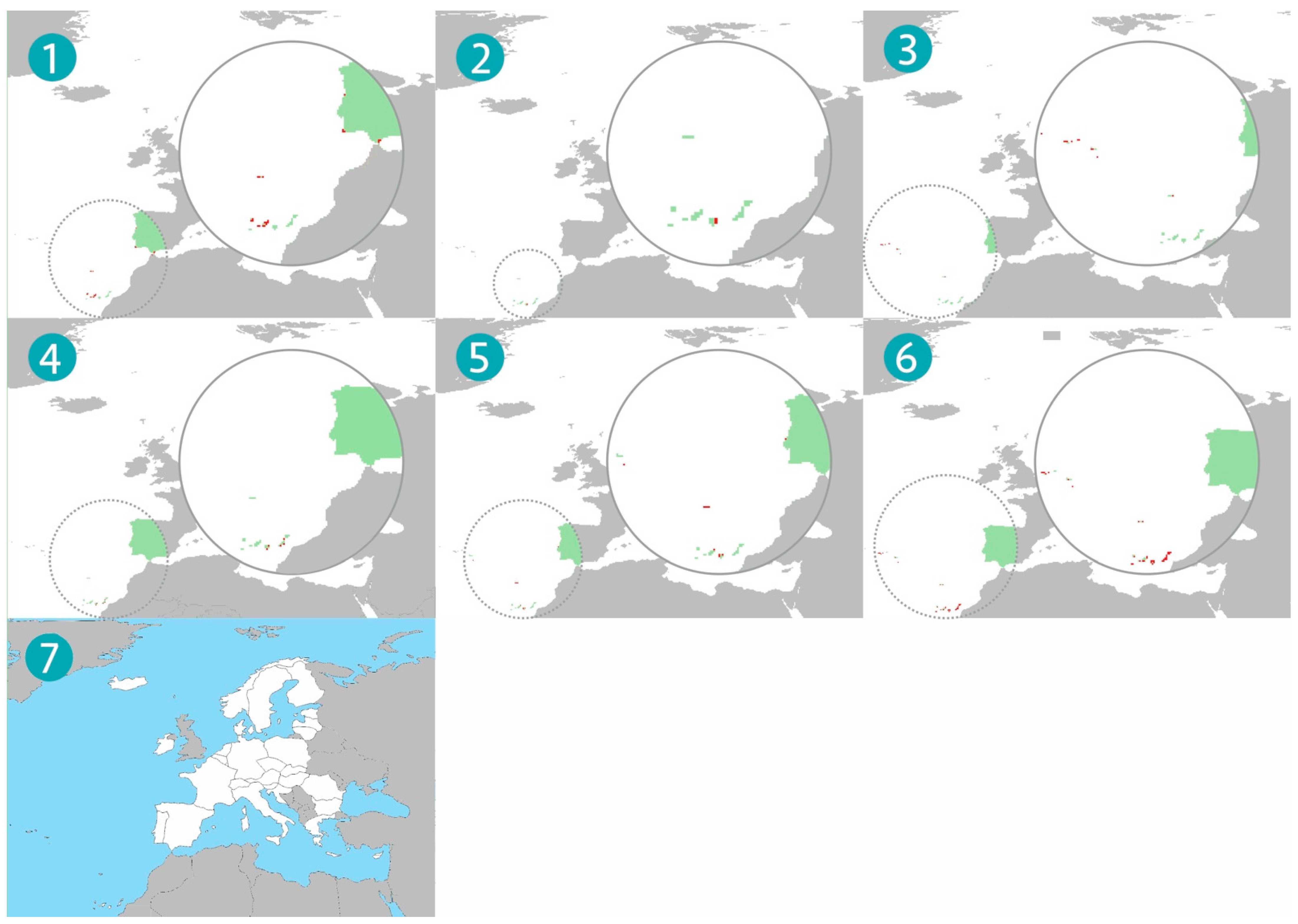
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clavero, M.; García-Berthou, E. Invasive species are a leading cause of animal extinctions. Trends Ecol. Evol. 2005, 20, 110. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Goudie, A.S. Human Impact on the Natural Environment: Past, Present and Future, 7th ed.; John Wiley & Sons: Oxford, UK, 2019. [Google Scholar]
- Pimentel, D. Biological Invasions: Economic and Environmental Costs of Alien Plant, Animal, and Microbe Species; Taylor & Francis: Abingdon, UK, 2011. [Google Scholar]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Nentwig, W. Biological Invasions: Why it Matters. In Biological Invasions; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–6. [Google Scholar]
- Juřičková, L. Subulina octona (Bruguière, 1798)—A new greenhouse species for the Czech Republic (Mollusca: Gastropoda: Subulinidae). Malacol. Bohemoslov. 2006, 5, 1–2. [Google Scholar] [CrossRef]
- Richling, I.; von Proschwitz, T. Identification problems of travelling snail species—New exotic introductions to tropical greenhouses in Gothenburg, Sweden (Gastropoda: Achatinellidae, Strobilopsidae, Helicarionidae). PeerJ 2021, 9, e11185. [Google Scholar] [CrossRef]
- Horsák, M.; Naggs, F.; Backeljau, T. Paropeas achatinaceum (Pfeiffer, 1846) and Other Alien Subulinine and Opeatine Land Snails in European Greenhouses (Gastropoda, Achatinidae). Malacologia 2020, 63, 123–130. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; de Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group Auckland: Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- Raut, S.K.; Barker, G.M. Achatina fulica Bowdich and other Achatinidae as pests in tropical agriculture. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI Publishing: Wallingford, UK, 2002; pp. 55–114. [Google Scholar]
- Nielsen, A.; Hatteland, B.A.; Malmstrøm, M.; von Proschwitz, T.; Velle, G.; de Boer, H.; Gjershaug, J.O.; Kirkendall, L.R.; Rueness, E.K.; Vandvik, V. Assessment of risks to Norwegian biodiversity from the import and keeping of terrestrial gastropods in terraria. In Scientific Opinion on the Panel on Alien Organisms and Trade in Endangered Species of the Norwegian Scientific Committee for Food Safety; Norwegian Scientific Committee for Food Safety: Oslo, Norway, 2017. [Google Scholar]
- Lv, S.; Zhang, Y.; Liu, H.-X.; Hu, L.; Yang, K.; Steinmann, P.; Chen, Z.; Wang, L.-Y.; Utzinger, J.; Zhou, X.-N. Invasive Snails and an Emerging Infectious Disease: Results from the First National Survey on Angiostrongylus cantonensis in China. PLoS Negl. Trop. Dis. 2009, 3, e368. [Google Scholar] [CrossRef]
- Cowie, R.H. Can snails ever be effective and safe biocontrol agents? Int. J. Pest Manag. 2001, 47, 23–40. [Google Scholar] [CrossRef]
- Thiengo, S.C.; Faraco, F.A.; Salgado, N.C.; Cowie, R.H.; Fernandez, M.A. Rapid spread of an invasive snail in South America: The giant African snail, Achatina fulica, in Brasil. Biol. Invasions 2007, 9, 693–702. [Google Scholar] [CrossRef]
- Gerlach, J.; Barker, G.M.; Bick, C.S.; Bouchet, P.; Brodie, G.; Christensen, C.C.; Collins, T.; Coote, T.; Cowie, R.H.; Fiedler, G.C.; et al. Negative impacts of invasive predators used as biological control agents against the pest snail Lissachatina fulica: The snail Euglandina ‘rosea’and the flatworm Platydemus manokwari. Biol. Invasions 2021, 23, 997–1031. [Google Scholar] [CrossRef]
- Guiller, A.; Martin, M.-C.; Hiraux, C.; Madec, L. Tracing the Invasion of the Mediterranean Land snail Cornu aspersum aspersum Becoming an Agricultural and Garden Pest in Areas Recently Introduced. PLoS ONE 2012, 7, e49674. [Google Scholar] [CrossRef]
- Holland, B.S.; Chock, T.; Lee, A.; Sugiura, S. Tracking behavior in the snail Euglandina rosea: First evidence of preference for endemic vs. biocontrol target pest species in Hawaii. Am. Malacol. Bull. 2012, 30, 153–157. [Google Scholar] [CrossRef]
- Cowie, R.H. Patterns of introduction of non-indigenous non-marine snails and slugs in the Hawaiian Islands. Biodivers. Conserv. 1998, 7, 349–368. [Google Scholar] [CrossRef]
- De Francesco, C.G.; Lagiglia, H. A predatory land snail invades central-western Argentina. Biol. Invasions 2007, 9, 795–798. [Google Scholar] [CrossRef]
- Aubry, S.; Labaune, C.; Magnin, F.; Roche, P.; Kiss, L. Active and passive dispersal of an invading land snail in Mediterranean France. J. Anim. Ecol. 2006, 75, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.H. The Dispersal of Cernuella-Virgata (Mollusca, Helicidae). Aust. J. Zool. 1988, 36, 513–520. [Google Scholar] [CrossRef]
- Baur, B. Patterns of dispersion, density and dispersal in alpine populations of the land snail Arianta arbustorum (L.) (Helicidae). Ecography 1986, 9, 117–125. [Google Scholar] [CrossRef]
- Kramarenko, S. Active and passive dispersal of terrestrial mollusks: A review. Ruthen. Russ. Malacol. J. 2014, 24, 1–14. [Google Scholar]
- Simonová, J.; Simon, O.P.; Kapic, Š.; Nehasil, L.; Horsák, M. Medium-sized forest snails survive passage through birds’ digestive tract and adhere strongly to birds’ legs: More evidence for passive dispersal mechanisms. J. Molluscan Stud. 2016, 82, 422–426. [Google Scholar] [CrossRef]
- Wada, S.; Kawakami, K.; Chiba, S. Snails can survive passage through a bird’s digestive system. J. Biogeogr. 2012, 39, 69–73. [Google Scholar] [CrossRef]
- Kolenda, K.; Najbar, A.; Kuśmierek, N.; Maltz, T.K. A possible phoretic relationship between snails and amphibians. Folia Malacol. 2017, 25, 281–285. [Google Scholar] [CrossRef]
- Reynolds, C.; Miranda, N.A.; Cumming, G.S. The role of waterbirds in the dispersal of aquatic alien and invasive species. Divers. Distrib. 2015, 21, 744–754. [Google Scholar] [CrossRef]
- Maciorowski, G.; Urbanska, M.; Gierszal, H. An example of passive dispersal of land snails by birds-short note. Folia Malacol. 2012, 20, 139–141. [Google Scholar] [CrossRef]
- Davis, M.A. Invasion Biology; Oxford University Press Inc.: New York, NY, USA, 2009; p. 241. [Google Scholar]
- Turbelin, A.J.; Malamud, B.D.; Francis, R.A. Mapping the global state of invasive alien species: Patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 2017, 26, 78–92. [Google Scholar] [CrossRef]
- Dörge, N.; Walther, C.; Beinlich, B.; Plachter, H. The significance of passive transport for dispersal in terrestrial snails (Gastropoda, Pulmonata). Z. Ökol. Nat. 1999, 8, 10. [Google Scholar]
- De Jager, K.; Daneel, M. Urocyclus flavescens Kerferstein (Urocyclidae) as a pest of banana in South Africa. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI Publishing: Wallingford, UK, 2002; pp. 235–239. [Google Scholar]
- Chang, C.-P. Bradybaena similaris (de Férussac) (Bradybaenidae) as a Pest on Grapevines of Taiwan. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI Publishing: Wallingford, UK, 2002; p. 241. [Google Scholar]
- Hammond, R.B.; Byers, R.A. Agriolimacidae and Arionidae as pests in conservation-tillage soybean and maize cropping in North America. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI Publishing: Wallingford, UK, 2002; pp. 301–314. [Google Scholar]
- Rueda, A.; Caballero, R.; Kaminsky, R.; Andrews, K.L. Vaginulidae in Central America, with emphasis on the bean slug Sarasinula plebeia (Fischer). In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI Publishing: Wallingford, UK, 2002; pp. 115–144. [Google Scholar]
- Gurevitch, J.; Padilla, D.K. Are invasive species a major cause of extinctions? Trends Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef]
- Patoka, J.; Bláha, M.; Kalous, L.; Kouba, A. Irresponsible vendors: Non-native, invasive and threatened animals offered for garden pond stocking. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 692–697. [Google Scholar] [CrossRef]
- Gherardi, F. Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Yonvitner, Y.; Patoka, J.; Yuliana, E.; Bohatá, L.; Tricarico, E.; Karella, T.; Kouba, A.; Reynolds, J.D. Enigmatic hotspot of crayfish diversity at risk: Invasive potential of non-indigenous crayfish if introduced to New Guinea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 219–224. [Google Scholar] [CrossRef]
- Wittenberg, R.; Cock, M.J. Invasive Alien Species: A Toolkit of Best Prevention and Management Practices; CABI Publishing: Wallingford, UK, 2001. [Google Scholar]
- Touza, J.; Dehnen-Schmutz, K.; Jones, G. Economic Analysis of Invasive Species Policies. In Biological Invasions; Springer: Berlin/Heidelberg, Germany, 2008; pp. 353–366. [Google Scholar]
- Mehta, S.V.; Haight, R.G.; Homans, F.R.; Polasky, S.; Venette, R.C. Optimal detection and control strategies for invasive species management. Ecol. Econ. 2007, 61, 237–245. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Robinet, C.; Kehlenbeck, H.; Van der Werf, W. Modelling the Spread of Invasive Species to Support Pest Risk Assessment: Principles and Application of a Suite of Generic Models. In Pest Risk Modelling and Mapping for Invasive Alien Species; Venette, R.C., Ed.; CABI Publishing: Wallingford, UK, 2015; pp. 115–130. [Google Scholar]
- Bomford, M.; Kraus, F.; Barry, S.C.; Lawrence, E. Predicting establishment success for alien reptiles and amphibians: A role for climate matching. Biol. Invasions 2009, 11, 713–724. [Google Scholar] [CrossRef]
- Akmal, S.G.; Jerikho, R.; Yulianda, F.; Wardiatno, Y.; Novák, J.; Kalous, L.; Slavík, O.; Patoka, J. Culture, trade and establishment of Polypterus senegalus in Indonesia with first record of wild populations. Aquac. Environ. Interact. 2022, 14, 127–133. [Google Scholar] [CrossRef]
- Patoka, J.; Magalhães, A.L.B.; Kouba, A.; Faulkes, Z.; Jerikho, R.; Vitule, J.R.S. Invasive aquatic pets: Failed policies increase risks of harmful invasions. Biodivers. Conserv. 2018, 27, 3037–3046. [Google Scholar] [CrossRef]
- Magalhães, A.L.B.; Andrade, R.F. Has the import ban on non-native red swamp crayfish (Crustacea: Cambaridae) been effective in Brazil? Neotrop. Biol. Conserv. 2014, 10, 48–52. [Google Scholar]
- Hulme, P.E. Handbook of Alien Species in Europe; Springer: Berlin/Heidelberg, Germany, 2009; Volume 569. [Google Scholar]
- Patoka, J.; Kopecký, O.; Vrabec, V.; Kalous, L. Aquarium molluscs as a case study in risk assessment of incidental freshwater fauna. Biol. Invasions 2017, 19, 2039–2046. [Google Scholar] [CrossRef]
- Padilla, D.K.; Williams, S.L. Beyond ballast water: Aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front. Ecol. Environ. 2004, 2, 131–138. [Google Scholar] [CrossRef]
- Ng, T.H.; Tan, S.K.; Wong, W.H.; Meier, R.; Chan, S.-Y.; Tan, H.H.; Yeo, D.C. Molluscs for sale: Assessment of freshwater gastropods and bivalves in the ornamental pet trade. PLoS ONE 2016, 11, e0161130. [Google Scholar] [CrossRef]
- Patoka, J.; Bláha, M.; Kalous, L.; Vrabec, V.; Buřič, M.; Kouba, A. Potential pest transfer mediated by international ornamental plant trade. Sci. Rep. 2016, 6, 25896. [Google Scholar] [CrossRef] [PubMed]
- Auliya, M.; Altherr, S.; Ariano-Sanchez, D.; Baard, E.H.; Brown, C.; Brown, R.M.; Cantu, J.-C.; Gentile, G.; Gildenhuys, P.; Henningheim, E. Trade in live reptiles, its impact on wild populations, and the role of the European market. Biol. Conserv. 2016, 204, 103–119. [Google Scholar] [CrossRef]
- Kalous, L.; Patoka, J.; Kopecký, O. European hub for invaders: Risk assessment of freshwater aquarium fishes exported from the Czech Republic. Acta Ichthyol. Piscat. 2015, 15, 239–245. [Google Scholar] [CrossRef]
- Bohatá, L.; Patoka, J. List of pet-traded terrestrial gastropods based on data from the Czech Republic. In Proceedings of the 11th Workshop on Biodiversity, Jevany, Czech Republic, 10 July 2019. [Google Scholar]
- Pyšek, P.; Danihelka, J.; Sádlo, J.; Chrtek Jr, J.; Chytrý, M.; Jarošík, V.; Kaplan, Z.; Krahulec, F.; Moravcová, L.; Pergl, J. Catalogue of alien plants of the Czech Republic: Checklist update, taxonomic diversity and invasion patterns. Preslia 2012, 84, 155–255. [Google Scholar]
- Phillips, S.J. A brief tutorial on Maxent. ATT Res. 2005, 190, 231–259. [Google Scholar]
- Roll, U.; Dayan, T.; Simberloff, D.; Mienis, H.K. Non-indigenous land and freshwater gastropods in Israel. Biol. Invasions 2009, 11, 1963–1972. [Google Scholar] [CrossRef]
- Cowie, R.H.; Dillon, R.T.; Robinson, D.G.; Smith, J.W. Alien Non-Marine Snails and Slugs of Priority Quarantine Importance in the United States: A Preliminary Risk Assessment. Am. Malacol. Bull. 2009, 27, 113–132. [Google Scholar] [CrossRef]
- Vogler, R.E.; Beltramino, A.A.; Sede, M.M.; Gregoric, D.E.G.; Núñez, V.; Rumi, A. The Giant African Snail, Achatina fulica (Gastropoda: Achatinidae): Using Bioclimatic Models to Identify South American Areas Susceptible to Invasion. Am. Malacol. Bull. 2013, 31, 39–50. [Google Scholar] [CrossRef]
- Prévot, V.; Jordaens, K.; Backeljau, T. Predominance of a single phylogenetic species in colonization events among a sextet of decollate land snail, Rumina decollata (Mollusca: Pulmonata: Subulinidae), species. Genome 2014, 57, 161–167. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Ward, D.F. Modelling the potential geographic distribution of invasive ant species in New Zealand. Biol. Invasions 2007, 9, 723–735. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Jiang, L.; Qiao, G. Predicting potential distribution of chestnut phylloxerid (Hemiptera: Phylloxeridae) based on GARP and Maxent ecological niche models. J. Appl. Entomol. 2010, 134, 45–54. [Google Scholar] [CrossRef]
- Giovanelli, J.G.; Haddad, C.F.; Alexandrino, J. Predicting the potential distribution of the alien invasive American bullfrog (Lithobates catesbeianus) in Brazil. Biol. Invasions 2008, 10, 585–590. [Google Scholar] [CrossRef]
- Oliveira, M.; Hamilton, S.; Calheiros, D.; Jacobi, C.; Latini, R. Modeling the potential distribution of the invasive golden mussel Limnoperna fortunei in the Upper Paraguay River system using limnological variables. Braz. J. Biol. 2010, 70, 831–840. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Shivambu, T.C.; Shivambu, N.; Downs, C.T. Exotic gastropods for sale: An assessment of land and aquatic snails in the South African pet trade. Manag. Biol. Invasions 2020, 11, 512–524. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Welbourne, D.J.; Romagosa, C.M.; Cassey, P.; Mandrak, N.E.; Strecker, A.; Leung, B.; Stringham, O.C.; Udell, B.; Episcopio-Sturgeon, D.J.; et al. When pets become pests: The role of the exotic pet trade in producing invasive vertebrate animals. Front. Ecol. Environ. 2019, 17, 323–330. [Google Scholar] [CrossRef]
- Vinarski, M.V.; Aksenova, O.V.; Bolotov, I.N.; Kondakov, A.V.; Khrebtova, I.S.; Gofarov MYu, S.M.; Zuykov, M. A new alien snail Ampullaceana balthica for the Canadian fauna, with an overview of Transatlantic malacofaunal exchange in the Anthropocene. Aquat. Invasions 2022, 17, 21–35. [Google Scholar] [CrossRef]
- Hossain, M.S.; Patoka, J.; Kouba, A.; Buřič, M. Clonal crayfish as biological model: A review on marbled crayfish. Biologia 2018, 73, 841–855. [Google Scholar] [CrossRef]
- Niemelä, P.; Mattson, W.J. Invasion of North American Forests by European Phytophagous Insects. BioScience 1996, 46, 741–753. [Google Scholar] [CrossRef]
- Gregoric, D.E.G.; Núñez, V.; Vogler, R.; Rumi, A. Invasion of the Argentinean Paranense Rainforest by the Giant African Snail Achatina fulica. Am. Malacol. Bull. 2011, 29, 135–137. [Google Scholar] [CrossRef]
- dos Santos, S.B.; Miyahira, I.C.; Mansur, M.C.D. Freshwater and terrestrial molluscs in Brasil: Current status of knowledge and conservation. Biol. Conserv. 2013, 90, 21–31. [Google Scholar]
- Vellend, M. The Biodiversity Conservation Paradox. Am. Sci. 2017, 105, 94–101. [Google Scholar] [CrossRef]
- Marková, J.; Jerikho, R.; Wardiatno, Y.; Kamal, M.M.; Magalhães, A.L.B.; Bohatá, L.; Kalous, L.; Patoka, J. Conservation paradox of giant arapaima Arapaima gigas (Schinz, 1822) (Pisces: Arapaimidae): Endangered in its native range in Brazil and invasive in Indonesia. Knowl. Manag. Aquat. Ecosyst. 2020, 47, 421. [Google Scholar] [CrossRef]
- Robinson, D.; Slapcinsky, J. Recent introductions of alien land snails into North America. Am. Malacol. Bull. 2005, 20, 89–93. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Author | Family | Breeding Difficulty | Native Geographic Distribution | Status |
|---|---|---|---|---|---|
| Acavus haemastoma | (Linnaeus, 1758) | Acavidae | medium | OL | x |
| Acavus superbus | (Pfeiffer, 1850) | Acavidae | medium | OL | x |
| Helicophanta bicingulata | (Smith, 1882) | Acavidae | medium | AT | x |
| Helicophanta magnifica | Férussac, 1819 | Acavidae | medium | AT | x |
| Oligospira waltoni | (Reeve, 1842) | Acavidae | medium | OL | x |
| Achatina achatina | (Linnaeus, 1758) | Achatinidae | easy | AT | P a |
| Achatina balteata | (Reeve, 1849) | Achatinidae | easy | AT | P a |
| Achatina craveni | (Smith, 1881) | Achatinidae | easy | AT | P a |
| Achatina schweinfurthi | (von Martens, 1874) | Achatinidae | medium | AT | P a |
| Achatina tincta | (Reeve, 1849) | Achatinidae | easy | AT | P a |
| Achatina weynsi | (Dautzenberg, 1900) | Achatinidae | easy | AT | P a |
| Archachatina degneri | (Bequaert and Clench, 1936) | Achatinidae | easy | AT | P a |
| Archachatina marginata | (Swainson, 1821) | Achatinidae | easy | AT | P a |
| Archachatina papyracea | (Pfeiffer, 1845) | Achatinidae | - | AT | P a |
| Archachatina purpurea | (Gmelin, 1790) | Achatinidae | easy | AT | P a |
| Archachatina puylaerti | (Mead, 1998) | Achatinidae | easy | AT | P a |
| Archachatina rhodostoma | (Philippi, 1849) | Achatinidae | easy | AT | P a |
| Archachatina ventricosa | (Gould, 1850) | Achatinidae | - | AT | P a |
| Ceras dautzenbergi | (Dupuis and Putzeys, 1901) | Achatinidae | easy | AT | x |
| Cochlitoma varicosa | (Pfeiffer, 1861) | Achatinidae | - | AT | P a |
| Limicolaria aurora | (Jay, 1839) | Achatinidae | easy | AT | P a |
| Limicolaria flammea | (Müller, 1774) | Achatinidae | easy | AT | P a |
| Limicolaria martensiana | (Smith, 1880) | Achatinidae | easy | AT | P a |
| Lissachatina albopicta | (Smith, 1878) | Achatinidae | easy | AT | P a |
| Lissachatina allisa | (Reeve, 1849) | Achatinidae | easy | AT | P a |
| Lissachatina fulica | (Bowdich, 1822) | Achatinidae | easy | AT | I b |
| Lissachatina immaculata | (Lamarck, 1822) | Achatinidae | easy | AT | P a |
| Lissachatina reticulata | (Pfeiffer, 1845) | Achatinidae | easy | AT | P a |
| Lissachatina zanzibarica | (Bourguignat, 1879) | Achatinidae | easy | AT | P a |
| Paropeas achatinaceum | (Pfeiffer, 1846) | Achatinidae | easy | OL | NN f |
| Pseudachatina downesii | (Sowerby I, 1838) | Achatinidae | hard | AT | P a |
| Rumina decollata | (Linnaeus, 1758) | Achatinidae | easy | PA | I a |
| Rumina saharica | (Pallary, 1901) | Achatinidae | easy | PA | MI, NN [12,60] |
| Subulina octona | (Bruguière, 1789) | Achatinidae | easy | NT | NN [7] |
| Ariophanta exilis | (Müller, 1774) | Airophantidae | easy | OL | x |
| Hemiplecta distincta | (Pfeiffer, 1850) | Airophantidae | medium | OL | x |
| Macrochlamys amboinensis | (von Martens, 1864) | Airophantidae | easy | OL | NN c |
| Hadra webbi | (Pilsbry, 1900) | Camaenidae | easy | AU | x |
| Oospira vanbuensis | (Bavay and Dautzenberg, 1899) | Clausiliidae | easy | OL | x |
| Phaedusa paviei | (Morlet, 1893) | Clausiliidae | easy | OL | x |
| Anguispira alternata | (Say, 1817) | Discidae | easy | NA | x |
| Anguispira strongylodes | (Pfeiffer, 1855) | Discidae | easy | NA | x |
| Pleurodonte isabella | (Férussac, 1822) | Pleurodontidae | easy | NT | x |
| Caracolus excellens | (Pfeiffer, 1853) | Solaropsidae | easy | NT | x |
| Caracolus marginella | (Gmelin, 1791) | Solaropsidae | easy | NT | x |
| Caracolus sagemon | (Beck, 1837) | Solaropsidae | easy | NT | x |
| Megalobulimus oblongus | (Müller, 1774) | Strophocheilidae | - | NT | NN d |
| Laevicaulis alte | (Férussac, 1822) | Veronicellidae | easy | AT | I [12,61] |
| Leidyula sloanii | (Cuvier, 1816) | Veronicellidae | easy | NT | P e, NN f |
| Zachrysia guanensis | (Poey, 1858) | Zachrysiidae | medium | NT | P d |
| Zachrysia provisoria | (Pfeiffer, 1858) | Zachrysiidae | - | NT | I e |
| Number | Variable | Minimum Temp (°C) | Maximum Temp (°C) | Rainfall (mm month−1) | Pan Evaporation (mm d−1) |
|---|---|---|---|---|---|
| Bio01 | Annual mean temperature (°C) | × | × | ||
| Bio02 | Mean diurnal temperature range (mean (period max–min)) (°C) | × | × | ||
| Bio03 | Isothermality (Bio02 ÷ Bio07) | × | × | ||
| Bio04 | Temperature seasonality (C of V) | × | × | ||
| Bio05 | Max temperature of warmest week (°C) | × | |||
| Bio06 | Min temperature of coldest week (°C) | × | |||
| Bio07 | Temperature annual range (Bio05–Bio06) (°C) | × | × | ||
| Bio08 | Mean temperature of wettest quarter (°C) | × | × | × | |
| Bio09 | Mean temperature of driest quarter (°C) | × | × | × | |
| Bio10 | Mean temperature of warmest quarter (°C) | × | × | ||
| Bio11 | Mean temperature of coldest quarter (°C) | × | × | ||
| Bio12 | Annual precipitation (mm) | × | |||
| Bio13 | Precipitation of wettest week (mm) | × | |||
| Bio14 | Precipitation of driest week (mm) | × | |||
| Bio15 | Precipitation seasonality (C of V) | × | |||
| Bio16 | Precipitation of wettest quarter (mm) | × | |||
| Bio17 | Precipitation of driest quarter (mm) | × | |||
| Bio18 | Precipitation of warmest quarter (mm) | × | × | × | |
| Bio19 | Precipitation of coldest quarter (mm) | × | × | × | |
| Bio28 | Annual mean moisture index | × | × | ||
| Bio29 | Highest weekly moisture index | × | × | ||
| Bio30 | Lowest weekly moisture index | × | × | ||
| Bio31 | Moisture index seasonality (C of V) | × | × | ||
| Bio32 | Mean moisture index of wettest quarter | × | × | ||
| Bio33 | Mean moisture index of driest quarter | × | × | ||
| Bio34 | Mean moisture index of warmest quarter | × | × | × | × |
| Bio35 | Mean moisture index of coldest quarter | × | × | × | × |
| Species | Family | CM (1–19, 28–35) | Threshold | AUC |
|---|---|---|---|---|
| Balance | ||||
| CM (1–19, 28–35) | CM (1–19, 28–35) | |||
| Acavus superbus | Acavidae | M | 0.757 | 0.994 |
| Helicophanta bicingulata | Acavidae | L | 1.168 | 0.998 |
| Helicophanta magnifica | Acavidae | S | 1.770 | 0.997 |
| Achatina achatina | Achatinidae | N | 1.226 | 0.997 |
| Achatina balteata | Achatinidae | S | 3.130 | 0.976 |
| Achatina schweinfurthi | Achatinidae | N | 0.771 | 0.985 |
| Archachatina marginata | Achatinidae | M | 1.094 | 0.993 |
| Archachatina ventricosa | Achatinidae | M | 2.372 | 0.999 |
| Cochlitoma varicosa | Achatinidae | L | 1.751 | 0.998 |
| Limicolaria flammea | Achatinidae | S | 2.164 | 0.963 |
| Limicolaria aurora | Achatinidae | S | 2.118 | 0.988 |
| Limicolaria martensiana | Achatinidae | N | 1.609 | 0.993 |
| Lissachatina allisa | Achatinidae | M | 2.177 | 0.988 |
| Lissachatina fulica | Achatinidae | S | 1.196 | 0.997 |
| Lissachatina reticulata | Achatinidae | S | 1.633 | 0.995 |
| Rumina decollata | Achatinidae | L | 1.694 | 0.982 |
| Rumina saharica | Achatinidae | L | 2.128 | 0.997 |
| Subulina octona | Achatinidae | N | 0.611 | 0.999 |
| Hemiplecta distincta | Airophantidae | N | 0.937 | 0.998 |
| Hadra webbi | Camaenidae | L | 1.068 | 0.990 |
| Phaedusa paviei | Clausiliidae | L | 3.337 | 0.998 |
| Anguispira alternata | Discidae | L | 1.422 | 0.955 |
| Anguispira strongylodes | Discidae | L | 1.358 | 0.995 |
| Caracolus marginella | Solaropsidae | N | 0.617 | 0.999 |
| Caracolus sagemon | Solaropsidae | N | 0.968 | 0.998 |
| Megalobulimus oblongus | Strophocheilidae | L | 2.722 | 0.980 |
| Laevicaulis alte | Veronicellidae | L | 3.806 | 0.98 |
| Zachrysia guanensis | Zachrysiidae | N | 0.993 | 0.999 |
| Zachrysia provisoria | Zachrysiidae | N | 1.066 | 0.999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bohatá, L.; Patoka, J. Invasion Potential of Ornamental Terrestrial Gastropods in Europe Based on Climate Matching. Diversity 2023, 15, 272. https://doi.org/10.3390/d15020272
Bohatá L, Patoka J. Invasion Potential of Ornamental Terrestrial Gastropods in Europe Based on Climate Matching. Diversity. 2023; 15(2):272. https://doi.org/10.3390/d15020272
Chicago/Turabian StyleBohatá, Lucie, and Jiří Patoka. 2023. "Invasion Potential of Ornamental Terrestrial Gastropods in Europe Based on Climate Matching" Diversity 15, no. 2: 272. https://doi.org/10.3390/d15020272
APA StyleBohatá, L., & Patoka, J. (2023). Invasion Potential of Ornamental Terrestrial Gastropods in Europe Based on Climate Matching. Diversity, 15(2), 272. https://doi.org/10.3390/d15020272