


DNA Barcode Library of Megadiverse Lepidoptera in an Alpine Nature Park (Italy) Reveals Unexpected Species Diversity

Abstract
1. Introduction
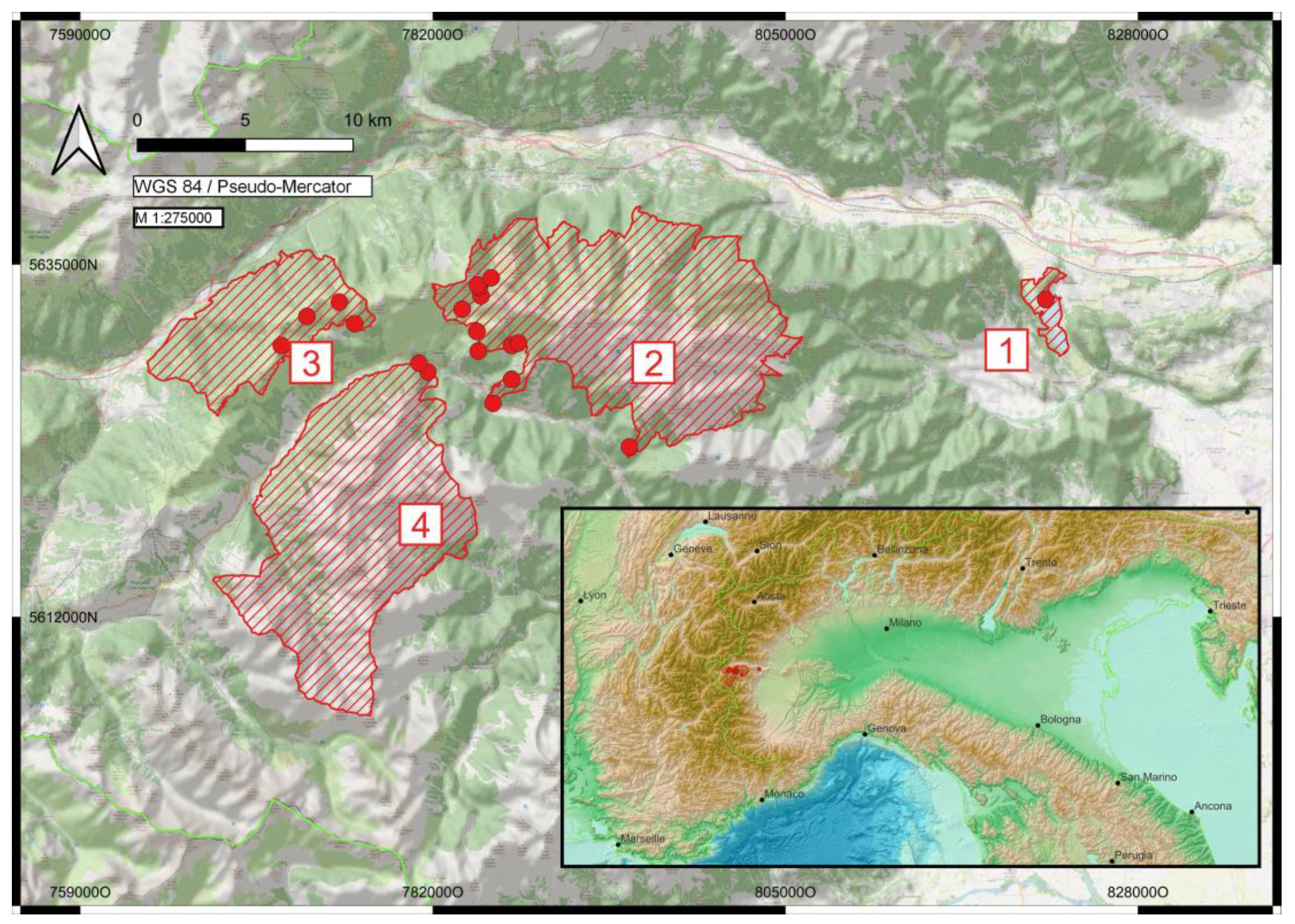
2. Materials and Methods
3. Results
3.1. General Overview—BIN System versus Linnean Taxa
3.2. BIN-Sharing Morphospecies
3.3. Cryptic Diversity
3.4. New Faunistic Records for Italy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigues, A.S.L.; Andelman, S.J.; Bakarr, M.I.; Boitani, L.; Brooks, T.M.; Cowling, R.M.; Fishpool, L.D.C.; da Fonseca, G.A.B.; Gaston, K.J.; Hoffmann, M.; et al. Effectiveness of the global protected area network in representing species diversity. Nature 2004, 428, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Bruner, A.G.; Gullison, R.E.; Rice, R.E.; Fonseca, G.A.B. Effectiveness of parks in protecting tropical biodiversity. Science 2001, 291, 125–128. [Google Scholar] [CrossRef]
- Engelbrecht, I. Invertebrate species inventories in protected area management: Are they useful? Afr. Entomol. 2010, 18, 235–245. [Google Scholar] [CrossRef]
- Encyclopaedia Britannica. Saving Earth. Species Inventories and Biodiversity Protection. Available online: https://www.britannica.com/explore/savingearth/species-inventories-and-biodiversity-protection (accessed on 29 December 2022).
- Pascher, K.; Švara, V.; Jungmeier, M. Environmental DNA-Based Methods in Biodiversity Monitoring of Protected Areas: Application Range, Limitations, and Needs. Diversity 2022, 14, 463. [Google Scholar] [CrossRef]
- De Biaggi, M.; Leccia, M.-F.; Kroupa, A.; Monje, J.C. Creating a biodiversity inventory in protected areas to increase knowledge of their natural heritage and to improve land management. J. Prot. Mt. Areas Res. Manag. 2010, 2, 49–52. [Google Scholar] [CrossRef]
- Ichter, J.; Leccia, M.-F.; Touroult, J.; Blandin, P.; Aberlenc, H.-P.; Holtof, J.-F.; Foret, J.; Bonet, R.; Pascal, O.; Dusoulier, F.; et al. Les inventaires généraux de la biodiversité en France et dans le monde. In Revue des All Taxa Biodiversity Inventories; UMS PatriNat/Parc national du Mercantour: Paris, France, 2010; 51p, Available online: http://www.patrinat.fr/sites/patrinat/files/atoms/files/2018/11/patrinat_2018_-_108_-_ichter_et_al_2018_les_inventaires_generaux_de_la_biodiversite_atbi.pdf (accessed on 2 January 2023).
- Hobern, D. BIOSCAN: DNA barcoding to accelerate taxonomy and biogeography for conservation and sustainability. Genome 2020, 64, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Mutanen, M.; Kivelä, S.M.; Vos, R.A.; Doorenweerd, C.; Ratnasingham, S.; Hausmann, A.; Huemer, P.; Dincă, V.; van Nieukerken, E.J.; Lopez-Vaamonde, C.; et al. Species-Level Para-and Polyphyly in DNA Barcode Gene Trees: Strong Operational Bias in European Lepidoptera. Syst. Biol. 2016, 65, 1024–1040. [Google Scholar] [CrossRef]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.N.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Res. 2021, 22, 803–822. [Google Scholar] [CrossRef]
- Bergsten, J.; Bilton, D.T.; Fujisawa, T.; Elliott, M.; Monaghan, M.T.; Balke, M.; Hendrich, L.; Geijer, J.; Herrmann, J.; Foster, G.N.; et al. The effect of geographical scale of sampling on DNA barcoding. Syst. Biol. 2012, 61, 851–869. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, A.; Haszprunar, G.; Segerer, A.H.; Speidel, W.; Behounek, G.; Hebert, P.D.N. Now DNA barcoded: The butterflies and larger moths of Germany (Lepidoptera: Rhopalocera, Macroheterocera). Spixiana 2011, 34, 47–58. [Google Scholar]
- Huemer, P.; Mutanen, M.; Sefc, K.M.; Hebert, P.D.N. Testing DNA Barcode Performance in 1000 Species of European Lepidoptera: Large Geographic Distances Have Small Genetic Impacts. PLoS ONE 2014, 9, e115774. [Google Scholar] [CrossRef] [PubMed]
- Huemer, P.; Hebert, P.D.N. DNA Barcode Bibliothek der Schmetterlinge Südtirols und Tirols (Italien, Österreich)–Impetus für integrative Artdifferenzierung im 21. Jahrhundert. Gredleriana 2016, 16, 141–164. [Google Scholar]
- Baldizzone, G. I microlepidotteri del Parco Naturale del Mont Avic e zone limitrofe (Valle d’Aosta-Val Chalamy-Alpi Graie orientali). Rev. Valdotaine Hist. Nat. 1997, 50, 55–141. [Google Scholar]
- Baldizzone, G. I Microlepidotteri del Parco Naturale Alpi Marittime (Italia, Piemonte) (Lepidoptera). Boll. Museo Reg. Sci. Nat. Torino 2004, 22, 1–318. [Google Scholar]
- Hellmann, E.; Bertaccini, E. I Macrolepidotteri della Valle di Susa Italia Nord-occidentale (Alpi Cozie-Graie). Monogr. Museo Reg. Sci. Nat. Torino 2004, 40, 392. [Google Scholar]
- Huemer, P.; Hebert, P.D.N. DNA-Barcoding von Schmetterlingen (Lepidoptera) in Waldstandorten Südtirols (IT01 Ritten und IT02 Montiggl). For. Obs. 2012, 6, 75–98. [Google Scholar]
- Piccini, I.; Depetris, M.; Paradiso, F.; Cochis, F.; Audisio, M.; Artioli, P.; Smargiassi, S.; Bonifacino, M.; Giuliano, D.; La Cava, S.; et al. Macro-moth (Lepidoptera) Diversity of a Newly Shaped Ecological Corridor and the Surrounding Forest Area in the Western Italian Alps. Diversity 2023, 15, 95. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. Available online: www.barcodinglife.org (accessed on 15 December 2022). [CrossRef]
- Ente di Gestione Dell Aree Protette Delle Alpi Cozie. Available online: https://www.parchialpicozie.it/ (accessed on 29 December 2022).
- Robinson, G.S. The preparation of slides of Lepidoptera genitalia with special reference to the Microlepidoptera. Entomol. S Gaz. 1976, 27, 127–132. [Google Scholar]
- deWaard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA Barcodes: Analytical Protocols. In Methods in Molecular Biology: Environmental Genomics; Martin, C.C., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 275–293. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef]
- Italian Ministry of the Environment. Checklist of the Italian Fauna On-Line. 2003. Available online: https://www.faunaitalia.it/checklist/ (accessed on 15 December 2022).
- Karsholt, O.; van Nieukerken, E.J. Lepidoptera, Moths. Fauna Europaea 2013, Version 2021.12. Available online: https://fauna-eu.org (accessed on 15 December 2022).
- Lepiforum: Website zur Bestimmung von Schmetterlingen (Lepidoptera) und Ihren Präimaginalstadien. Available online: http://www.lepiforum.org (accessed on 15 December 2022).
- Huemer, P.; Karsholt, O.; Wieser, C. Megacraspedus cottiensis sp. nov. (Lepidoptera, Gelechiidae) from northern Italy–a case of taxonomic confusion. ZooKeys 2020, 963, 141–152. [Google Scholar] [CrossRef]
- Huemer, P. Integrative revision of the Caryocolum schleichi species group–A striking example of a temporally changing species concept (Lepidoptera, Gelechiidae). Alp. Entomol. 2020, 4, 39–63. [Google Scholar] [CrossRef]
- Hellmann, F.; Parenzan, P. I Macrolepidotteri del Piemonte. Museo Regionale die Scienze Naturali; Monografie XLVI: Torino, Italy, 2010; pp. 1–1058. [Google Scholar]
- Huemer, P.; Wieser, C. Bemerkenswerte Neufunde von Schmetterlingen (Lepidoptera) für Italien (Cottische Alpen). Carinth. II 2020, 210, 457–470. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D.N. BOLD’s role in barcode data management and analysis: A response. Mol. Ecol. Notes 2011, 11, 941–942. [Google Scholar] [CrossRef]
- deWaard, J.R.; Ratnasingham, S.; Zakharov, E.V.; Borisenko, A.V.; Steinke, D.; Telfer, A.C.; Perez, K.H.J.; Sones, J.E.; Young, M.R.; Levesque-Beaudin, V.; et al. A reference library for the identification of Canadian invertebrates: 1.5 million DNA barcodes, voucher specimens, and genomic samples. Sci. Data 2019, 6, 308. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; deWaard, J.R.; Zakharov, E.V.; Prosser, S.W.J.; Sones, J.E.; McKeown, J.T.A.; Mantle, B.; La Salle, J. A DNA ‘barcode blitz’: Rapid digitization and sequencing of a natural history collection. PLoS ONE 2013, 10, e68535. [Google Scholar] [CrossRef] [PubMed]
- Zahiri, R.; Lafontaine, J.D.; Schmidt, B.C.; deWaard, J.R.; Zakharov, E.V.; Hebert, P.D.N. A transcontinental challenge—A test of DNA barcode performance for 1,541 species of Canadian Noctuoidea (Lepidoptera). PLoS ONE 2014, 9, e92797. [Google Scholar] [CrossRef]
- Dinca, V.; Dapporto, L.; Somervuo, P.; Vodă, R.; Cuvelier, S.; Gascoigne-Pees, M.; Huemer, P.; Mutanen, M.; Hebert, P.D.N.; Vila, R. High resolution DNA barcode library for European butterflies reveals continental patterns of mitochondrial genetic diversity. Commun. Biol. 2021, 4, 315. [Google Scholar] [CrossRef]
- Huemer, P.; Karsholt, O.; Aarvik, L.; Berggren, K.; Bidzilya, O.; Junnilainen, J.; Landry, J.-F.; Mutanen, M.; Nupponen, K.; Segerer, A.; et al. DNA barcode library for European Gelechiidae (Lepidoptera) suggests greatly underestimated species diversity. ZooKeys 2020, 921, 141–157. [Google Scholar] [CrossRef]
- Lopez-Vaamonde, C.; Kirichenko, N.; Cama, A.; Doorenweerd, C.; Godfray, H.C.J.; Guiguet, A.; Gomboc, S.; Huemer, P.; Landry, J.-F.; Laštůvka, A.; et al. Evaluating DNA Barcoding for Species Identification and Discovery in European Gracillariid Moths. Front. Ecol. Evol. 2021, 9, 626752. [Google Scholar] [CrossRef]
- Huemer, P.; Mutanen, M. An Incomplete European Barcode Library Has a Strong Impact on the Identification Success of Lepidoptera from Greece. Diversity 2022, 14, 118. [Google Scholar] [CrossRef]
- Hausmann, A.; Godfray, H.C.J.; Huemer, P.; Mutanen, M.; Rougerie, R.; van Nieukerken, E.J.; Ratnasingham, S.; Hebert, P.D.N. Genetic Patterns in European Geometrid Moths Revealed by the Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e84518. [Google Scholar] [CrossRef]
- Huemer, P.; Wieser, C.; Stark, W.; Hebert, P.D.N.; Wiesmair, B. DNA barcode library of megadiverse Austrian Noctuoidea (Lepidoptera)—A nearly perfect match of Linnean taxonomy. Biodivers. Data J. 2019, 7, e37734. [Google Scholar] [CrossRef]
- Ortiz, A.; Rubio, R.; Guerrero, J.; Garre, M.; Serrano, J.; Hebert, P.D.N.; Hausmann, A. Close congruence between Barcode Index Numbers (bins) and species boundaries in the Erebidae (Lepidoptera: Noctuoidea) of the Iberian Peninsula. Biodivers. Data J. 2017, 5, e19840. [Google Scholar] [CrossRef]
- Talavera, G.; Lukhtanov, V.A.; Rieppel, L.; Pirece, N.E. In the shadow of phylogenetic uncertainty: The recent diversification of Lysandra butterflies through chromosomal change. Mol. Phylogenet. Evol. 2013, 69, 449–478. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, J.R.; Roe, A.D.; Sperling, F.A.H. Multi-locus species delimitation in closely related animal and fungi: One marker is not enough. Mol. Ecol. 2012, 21, 4422–4436. [Google Scholar] [CrossRef]
- Huemer, P.; Karsholt, O. Revision of the genus Megacraspedus Zeller, 1839, a challenging taxonomic tightrope of species delimitation (Lepidoptera, Gelechiidae). ZooKeys 2018, 800, 1–278. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–417. [Google Scholar] [CrossRef]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Janzen, D.H.; Hallwachs, W. DNA barcoding the Lepidoptera inventory of a large complex tropical conserved wildland, Area de Conservacion Guanacaste, northwestern Costa Rica. Genome 2016, 59, 641–660. [Google Scholar] [CrossRef] [PubMed]
- Deharveng, L.; Bedos, A.; Daugeron, C.; Villemant, C.; Judson, M.L.I. Organization, usefulness and limitations of an ATBI (All Taxa Biodiversity Inventory): The inventory in the Mercantour National Park. Zoosystema 2015, 37, 9–30. [Google Scholar] [CrossRef]
- Ichter, J.; Gargominy, O.; Leccia, M.-F.; Robert, S.; Poncet, L. The first large-scale All Taxa Biodiversity Inventory in Europe: Description of the Mercantour National Park ATBI datasets. Biodivers. Data J. 2022, 10, e85901. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Family | Species | BIN | BIN-sp |
|---|---|---|---|
| Coleophoridae | Coleophora nutantella | BOLD:AAE1255 | 4 |
| Coleophora silenella | |||
| Depressariidae | Agonopterix alpigena | BOLD:AAC0206 | 48 |
| Agonopterix angelicella | |||
| Agonopterix atomella | |||
| Agonopterix ivinskisi | |||
| Agonopterix kaekeritziana | |||
| Agonopterix scopariella | |||
| Geometridae | Chloroclysta miata | BOLD:AAA9515 | 2 |
| Chloroclysta siterata | |||
| Geometridae | Perizoma affinitata | BOLD:AAC0743 | 2 |
| Perizoma hydrata | |||
| Geometridae | Thera britannica | BOLD:AAA7522 | 4 |
| Thera obeliscata | |||
| Thera variata | |||
| Thera vetustata | |||
| Lasiocampidae | Poecilocampa alpina | BOLD:AAC8994 | 4 |
| Poecilocampa coluchei | |||
| Lycaenidae | Lysandra bellargus | BOLD:AAA3305 | 12 |
| Lysandra coridon | |||
| Lycaenidae | Polyommatus eros | BOLD:AAA3303 | 25 |
| Polyommatus icarus | |||
| Noctuidae | Conistra ligula | BOLD:AAB7880 | 4 |
| Conistra vaccinii | |||
| Noctuidae | Euxoa obelisca | BOLD:ACE8354 | 10 |
| Euxoa tritici | |||
| Euxoa vitta | |||
| Noctuidae | Lacanobia oleracea | BOLD:ABY4614 | 3 |
| Lacanobia splendens | |||
| Noctuidae | Mesapamea secalella | BOLD:AAB2749 | 4 |
| Mesapamea secalis | |||
| Tortricidae | Apotomis capreana | BOLD:ABZ6958 | 14 |
| Apotomis semifasciana | |||
| Yponomeutidae | Yponomeuta cagnagella | BOLD:AAA7740 | 10 |
| Yponomeuta padella |
| Family | Species | BIN |
|---|---|---|
| Coleophoridae | Coleophora nutantella | BOLD:AAE1255 |
| Coleophora silenella | ||
| Geometridae | Perizoma affinitata | BOLD:AAC0743 |
| Perizoma hydrata | ||
| Geometridae | Thera britannica | BOLD:AAA7522 |
| Thera obeliscata | ||
| Thera variata | ||
| Lasiocampidae | Poecilocampa alpina | BOLD:AAC8994 |
| Poecilocampa coluchei | ||
| Lycaenidae | Lysandra bellargus | BOLD:AAA3305 |
| Lysandra coridon | ||
| Noctuidae | Euxoa obelisca | BOLD:ACE8354 |
| Euxoa tritici | ||
| Euxoa vitta | ||
| Yponomeutidae | Yponomeuta cagnagella | BOLD:AAA7740 |
| Yponomeuta padella |
| Species | BIN | BM | BMC | Dist.NN | BIN NN | NN |
|---|---|---|---|---|---|---|
| Ephysteris sp. 1 | BOLD:ADZ8459 | 3 | 3 | 2.01% | BOLD:AEF8437 | Ephysteris inustella |
| Orophia sp. | BOLD:AEA1426 | 1 | 1 | 2.06% | BOLD:AAK0640 | Orophia sordidella |
| Agapeta sp. | BOLD:AEG7231 | 1 | 1 | 2.20% | BOLD:AAA6574 | Agapeta zoegana |
| Alabonia sp.* | BOLD:AEA2906 | 7 | 6 | 2.24% | BOLD:AAJ2485 | Alabonia geoffrella |
| Odezia sp. | BOLD:AEC3295 | 1 | 1 | 2.4% | BOLD:ACB1463 | Odezia atrata |
| Coleophora sp. 1 | BOLD:AET5871 | 1 | 1 | 2.51% | BOLD:AAE8838 | Coleophora lithargyrinella |
| Ephestia sp. ** | BOLD:ACY1394 | 2 | 1 | 2.56% | BOLD:AEC9178 | Ephestia welseriella |
| Scrobipalpa sp. ** | BOLD:ACX8455 | 5 | 1 | 2.56% | BOLD:ACY6331 | Scrobipalpa reiprichi |
| Eupithecia sp. | BOLD:AEC3281 | 6 | 6 | 2.56% | BOLD:AEC9229 | Eupithecia graphata hesperia |
| Bijugis sp. | BOLD:ADZ9474 | 1 | 1 | 2.58% | BOLD:AAF7451 | Bijugis bombycella |
| Infurcitinea sp. | BOLD:AEM6427 | 1 | 1 | 2.72% | BOLD:AEM9389 | Infurcitinea atrifasciella |
| Ochsenheimeria sp. | BOLD:AET5252 | 1 | 1 | 2.72% | BOLD:AER7793 | Ochsenheimeria taurella |
| Ephysteris sp. 2 | BOLD:AEA3472 | 3 | 3 | 2.88% | BOLD:ADM1235 | Ephysteris olympica |
| Psodos sp. ** | BOLD:AAE6737 | 2 | 1 | 3.21% | BOLD:AAO1675 | Glacies canaliculata |
| Trifurcula sp. ** | BOLD:ADQ7627 | 2 | 1 | 3.21% | BOLD:ACG9008 | Trifurcula orientella |
| Caryocolum sp. ** | BOLD:ADG5831 | 5 | 1 | 4.33% | BOLD:AAV0572 | Carycolum alsinella |
| Denisia sp. | BOLD:AEC5998 | 1 | 1 | 4.49% | BOLD:AAU0845 | Denisia rhaetica |
| Kessleria sp. * | BOLD:AEE4715 | 7 | 6 | 4.49% | BOLD:AAK3930 | Kessleria caflischiella |
| Coleophora sp. 2 ** | BOLD:ACT0442 | 2 | 1 | 5.13% | BOLD:ABA4033 | Coleophora vulpecula |
| Denisia sp. | BOLD:AEF7084 | 3 | 3 | 5.43% | BOLD:AAD4867 | Denisia haydenella |
| Elachista sp. * | BOLD:AEM6237 | 4 | 2 | 6.19% | BOLD:ACG7227 | Elachista sp. |
| Glyphipterix sp. ** | BOLD:ABY3946 | 5 | 1 | 7.21% | BOLD:AAT9919 | Glyphipterix equitella |
| Taxon | Family |
|---|---|
| Coleophora tricolor Walsingham, 1899 | Coleophoridae |
| Agonopterix ivinskisi Lvovsky, 1992 * | Depressariidae |
| Depressaria gallicella Chrétien, 1908 | Depressariidae |
| Elachista wieseriella Huemer, 2000 | Elachistidae |
| Athrips pruinosella (Lienig and Zeller, 1846) | Gelechiidae |
| Aproaerema incognitana (Gozmány, 1957) * | Gelechiidae |
| Ephysteris olympica Povolný, 1968 * | Gelechiidae |
| Megacraspedus peslieri Huemer and Karsholt, 2018 * | Gelechiidae |
| Stomopteryy pyrenaeella Varenne and Nel, 2020 | Gelechiidae |
| Tila capsophilella (Chrétien, 1900) | Gelechiidae |
| Mercantouria neli Huemer, Lopez-Vaamonde, and Triberti, 2016 | Gracillariidae |
| Agnoea langohri (Palm, 1990) * | Lypusidae |
| Procapperia linariae (Chrétien, 1922) * | Pterophoridae |
| Sphinx maurorum (Jordan, 1931) | Sphingidae |
| Pammene epanthista (Meyrick, 1932) * | Tortricidae |
| Euhyponomeutoides albithoracellus Gaj, 1954 | Yponomeutidae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huemer, P.; Wieser, C. DNA Barcode Library of Megadiverse Lepidoptera in an Alpine Nature Park (Italy) Reveals Unexpected Species Diversity. Diversity 2023, 15, 214. https://doi.org/10.3390/d15020214
Huemer P, Wieser C. DNA Barcode Library of Megadiverse Lepidoptera in an Alpine Nature Park (Italy) Reveals Unexpected Species Diversity. Diversity. 2023; 15(2):214. https://doi.org/10.3390/d15020214
Chicago/Turabian StyleHuemer, Peter, and Christian Wieser. 2023. "DNA Barcode Library of Megadiverse Lepidoptera in an Alpine Nature Park (Italy) Reveals Unexpected Species Diversity" Diversity 15, no. 2: 214. https://doi.org/10.3390/d15020214
APA StyleHuemer, P., & Wieser, C. (2023). DNA Barcode Library of Megadiverse Lepidoptera in an Alpine Nature Park (Italy) Reveals Unexpected Species Diversity. Diversity, 15(2), 214. https://doi.org/10.3390/d15020214