



Structural Organization of Zooplankton Communities in Different Types of River Mouth Areas

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
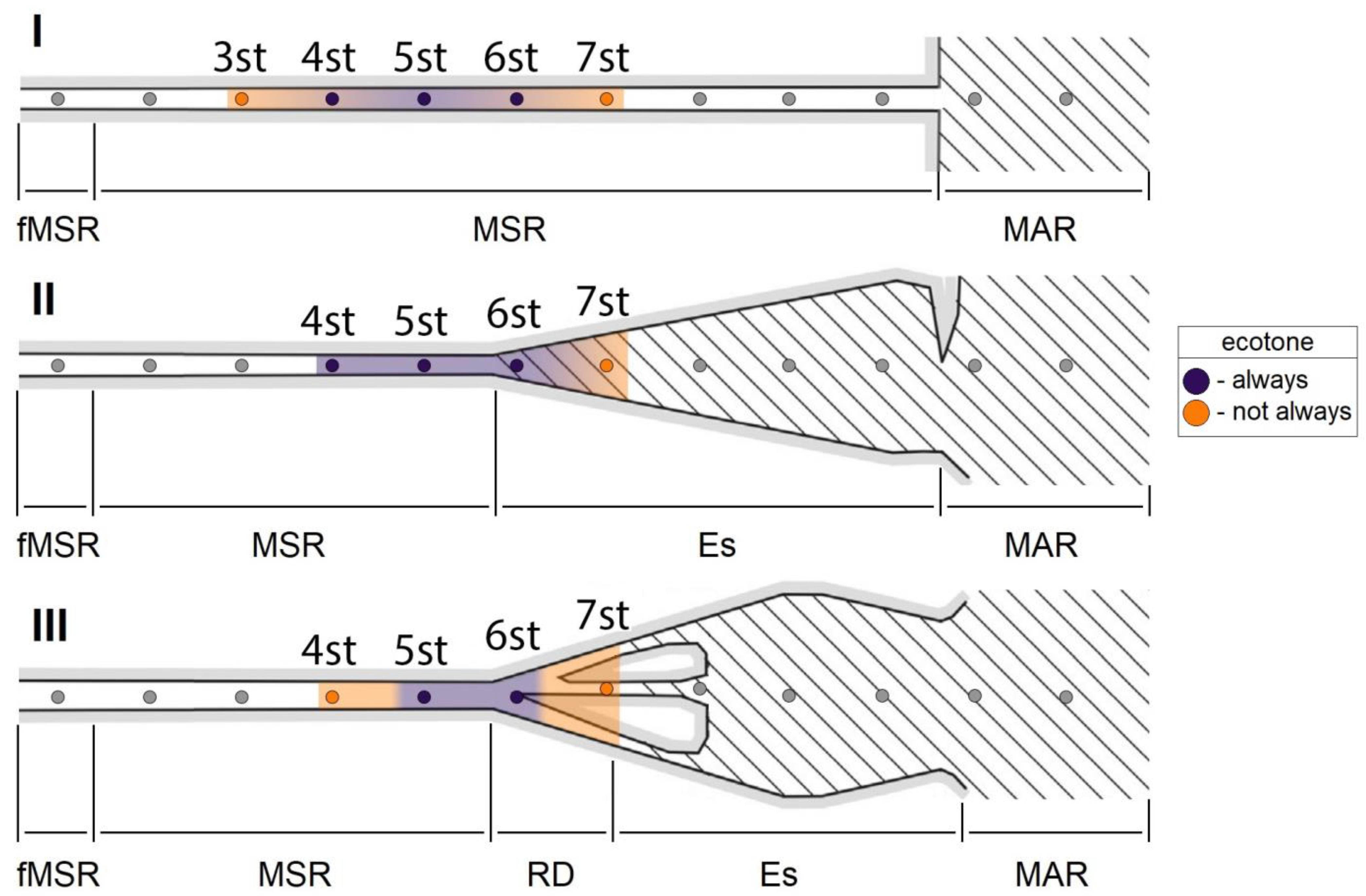
2.1. Study Area and Sample Collection
2.2. Species Identification
2.3. Environmental Indicators
2.4. Data Analysis
3. Results
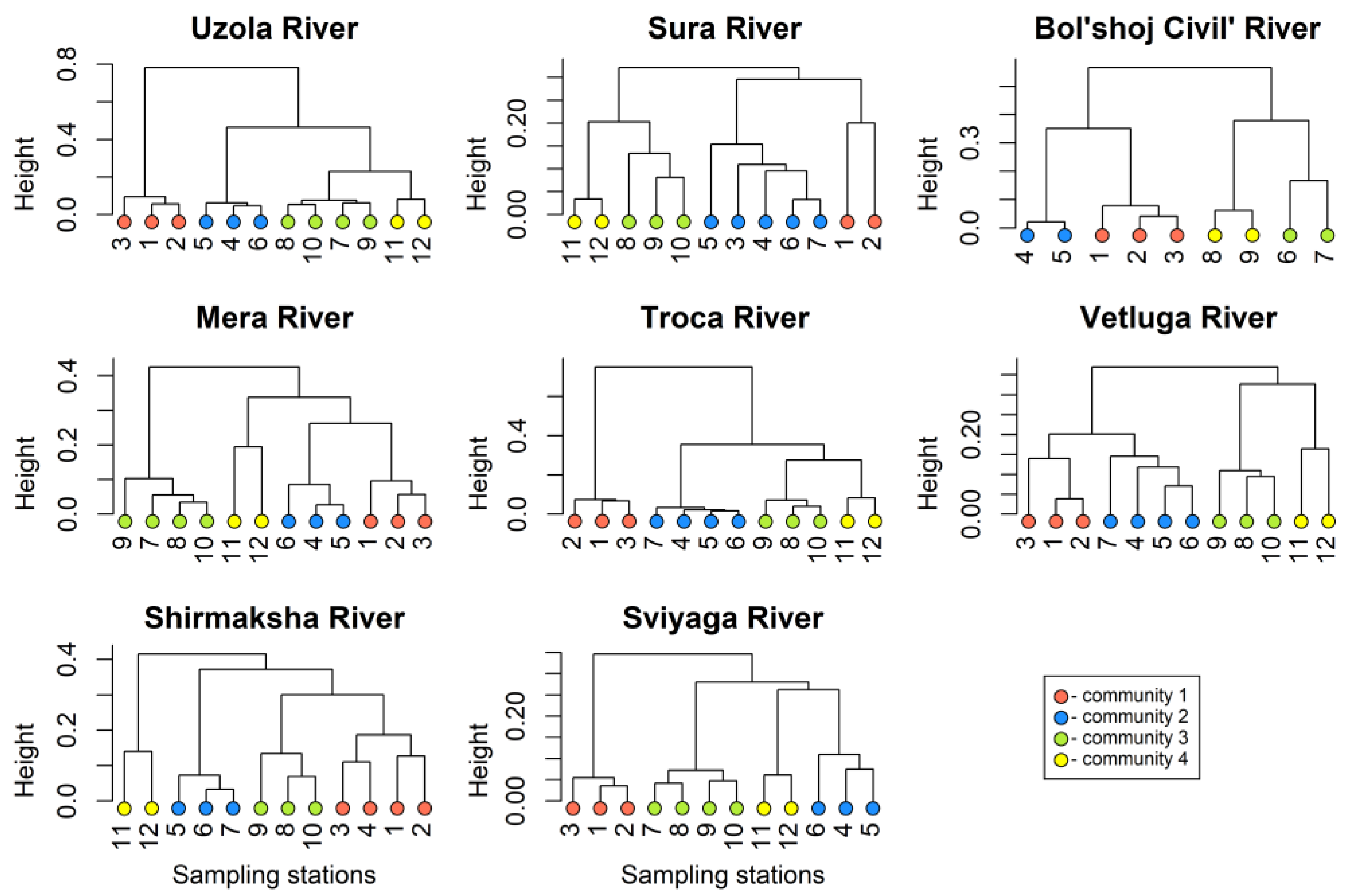
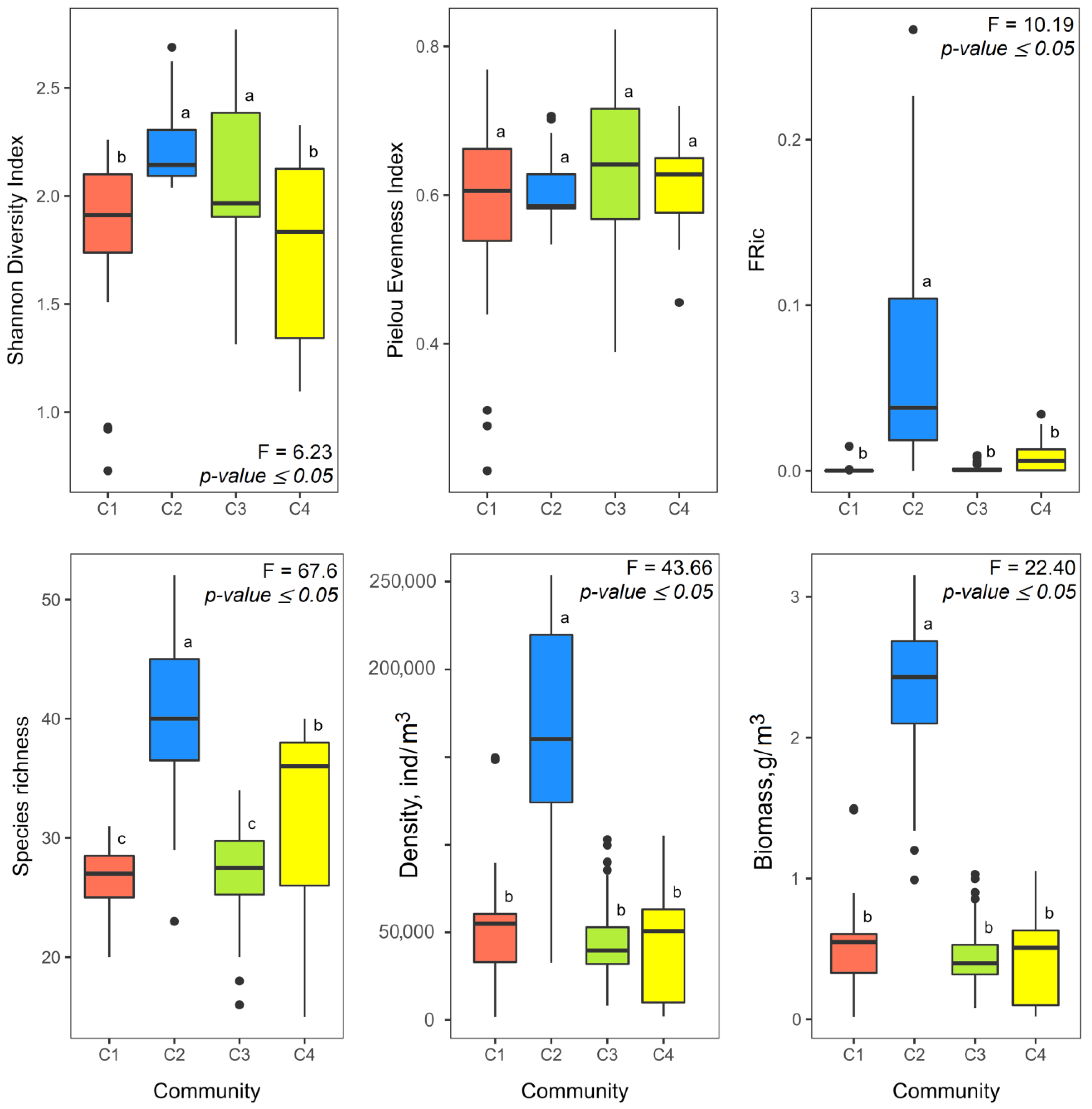
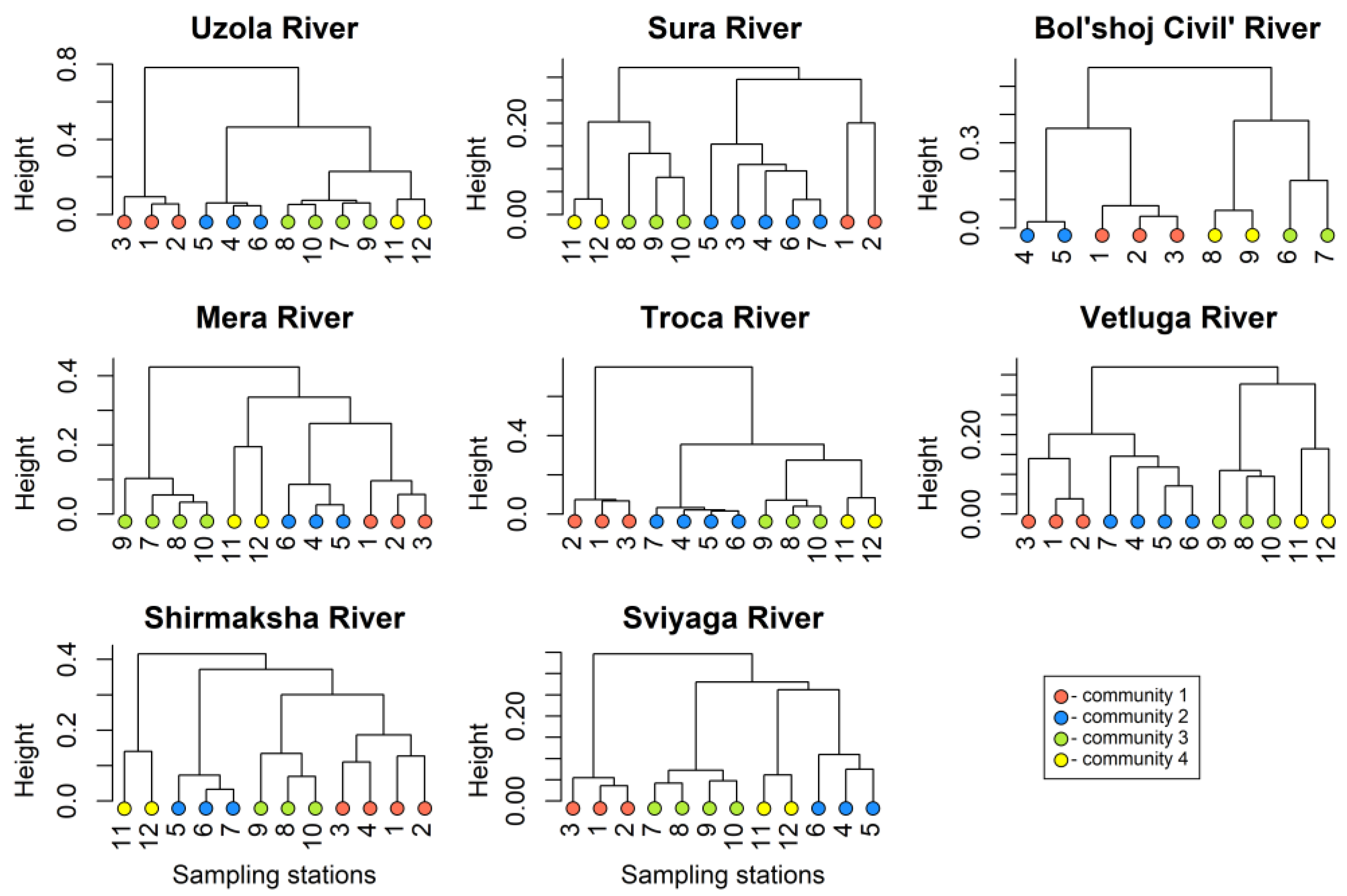
3.1. Zooplankton Communities
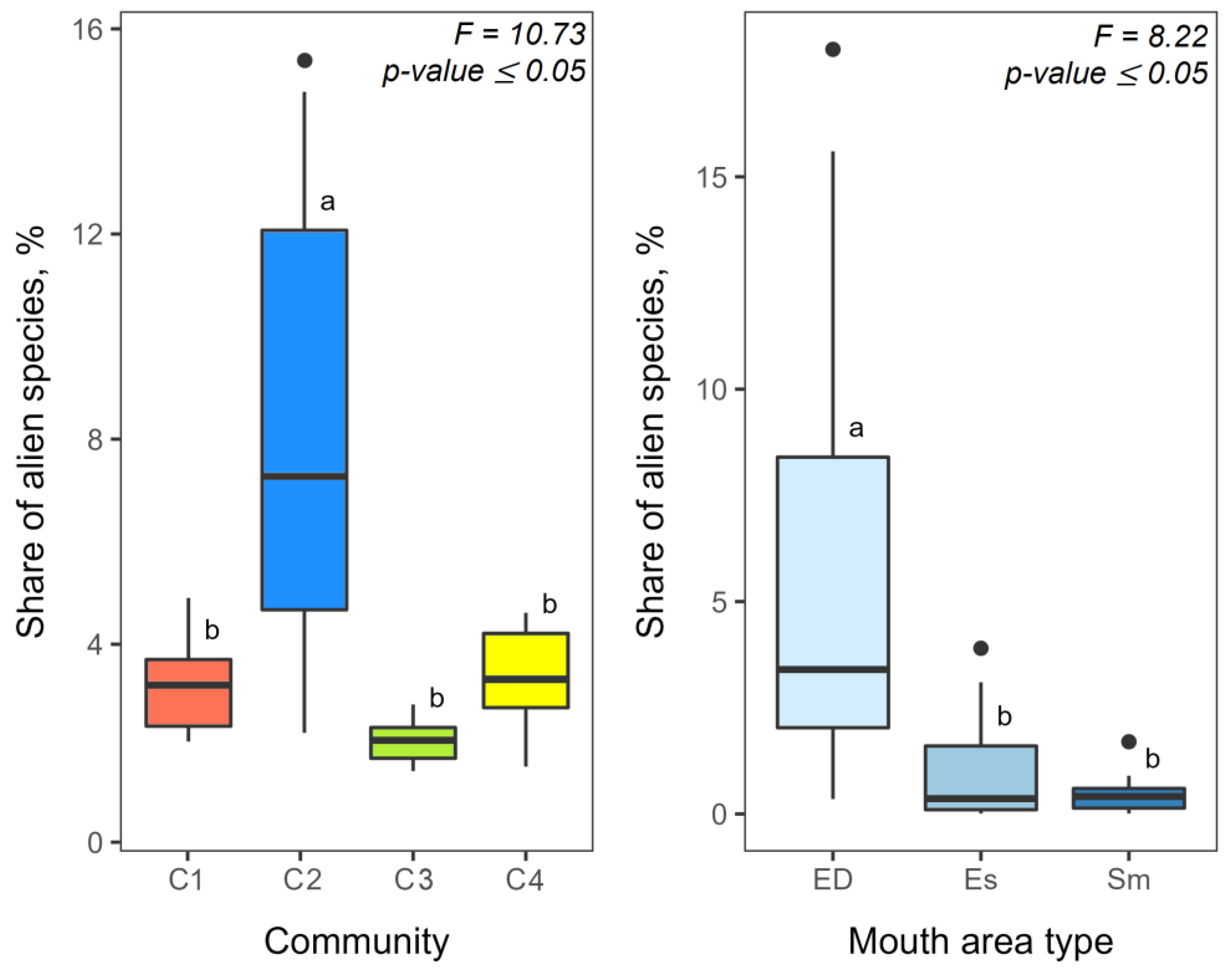
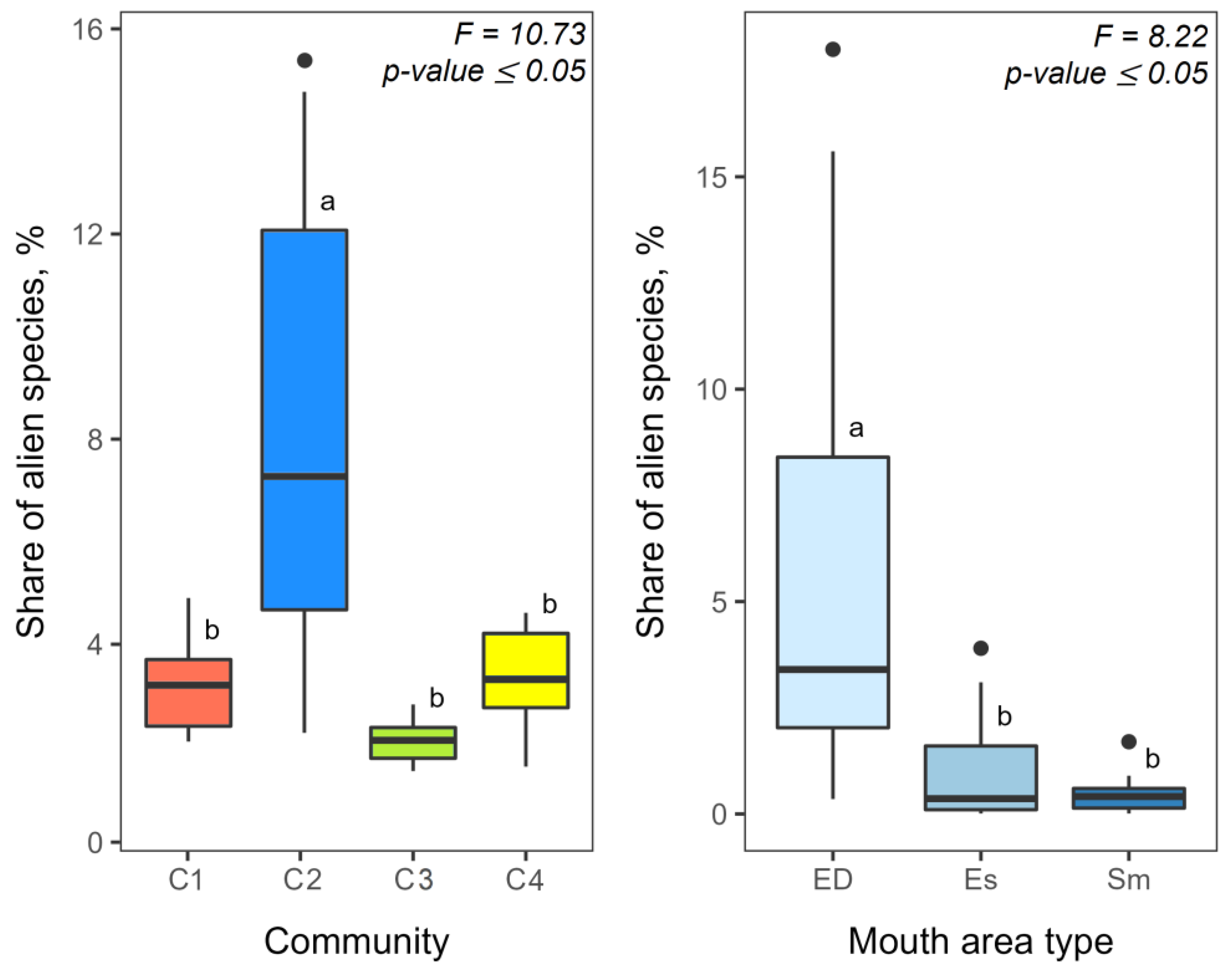
3.2. Alien Species and Its Share in Zooplankton Communities
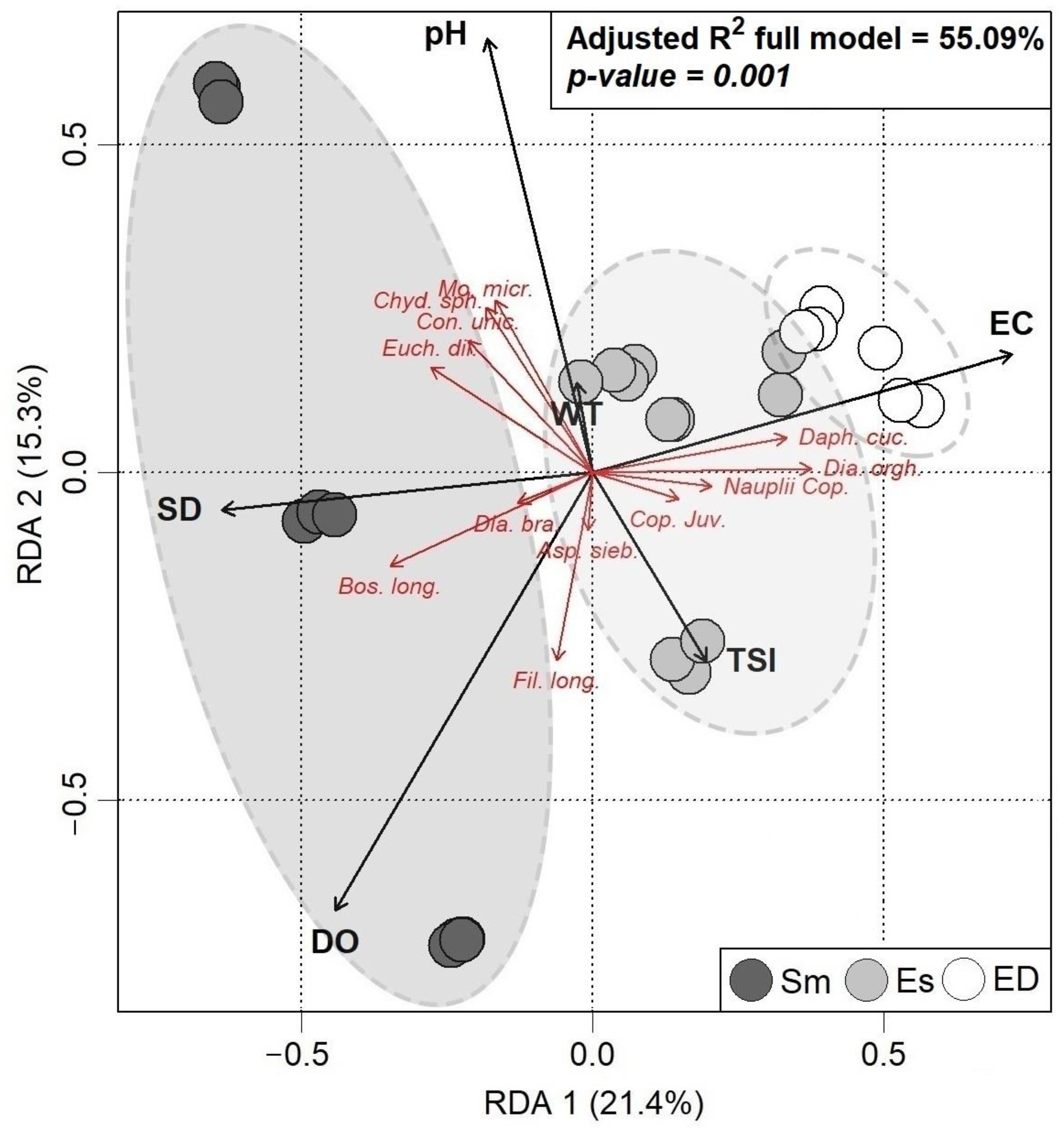
3.3. RDA of the Species Structure of Community 2
4. Discussion
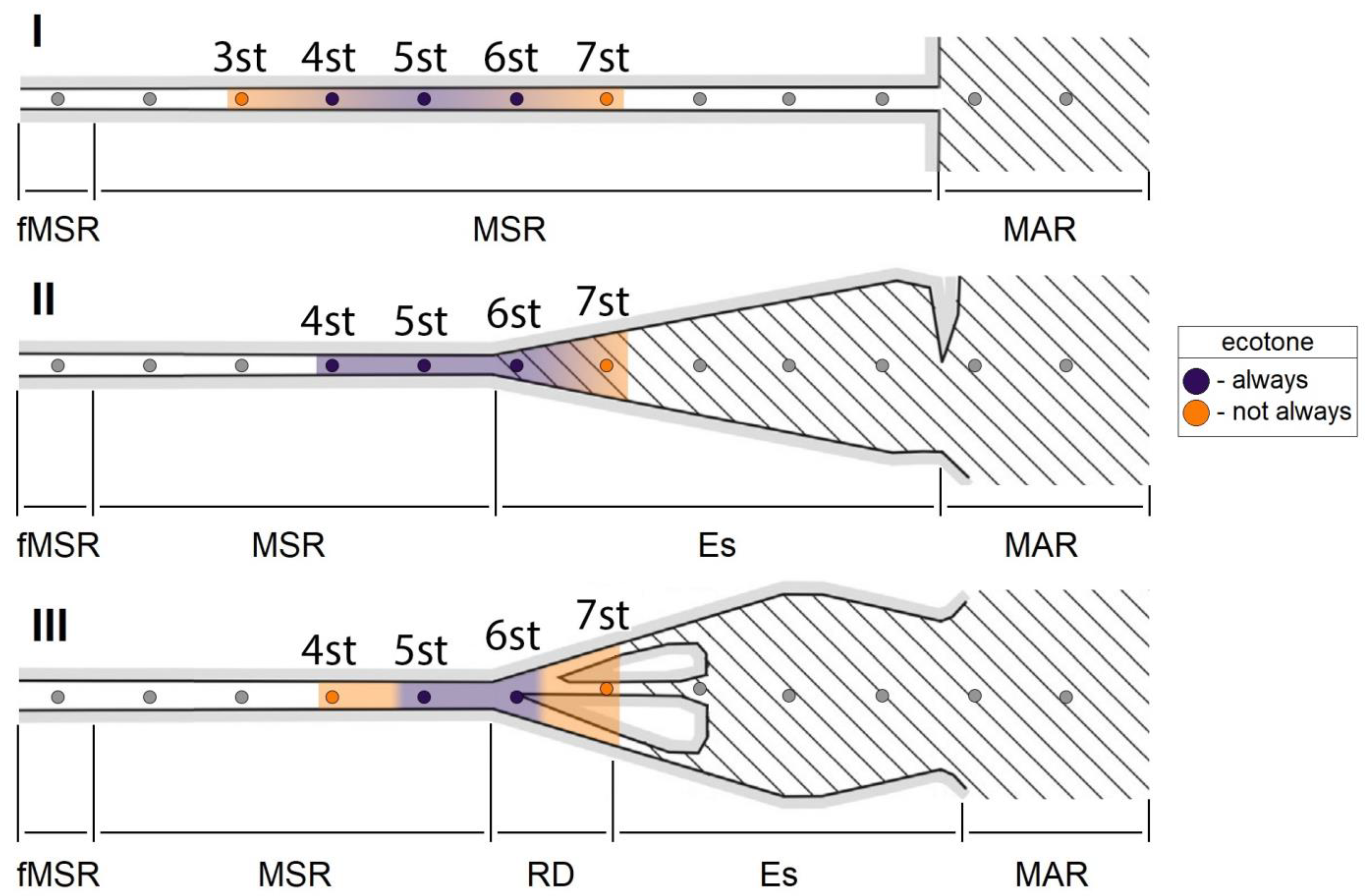
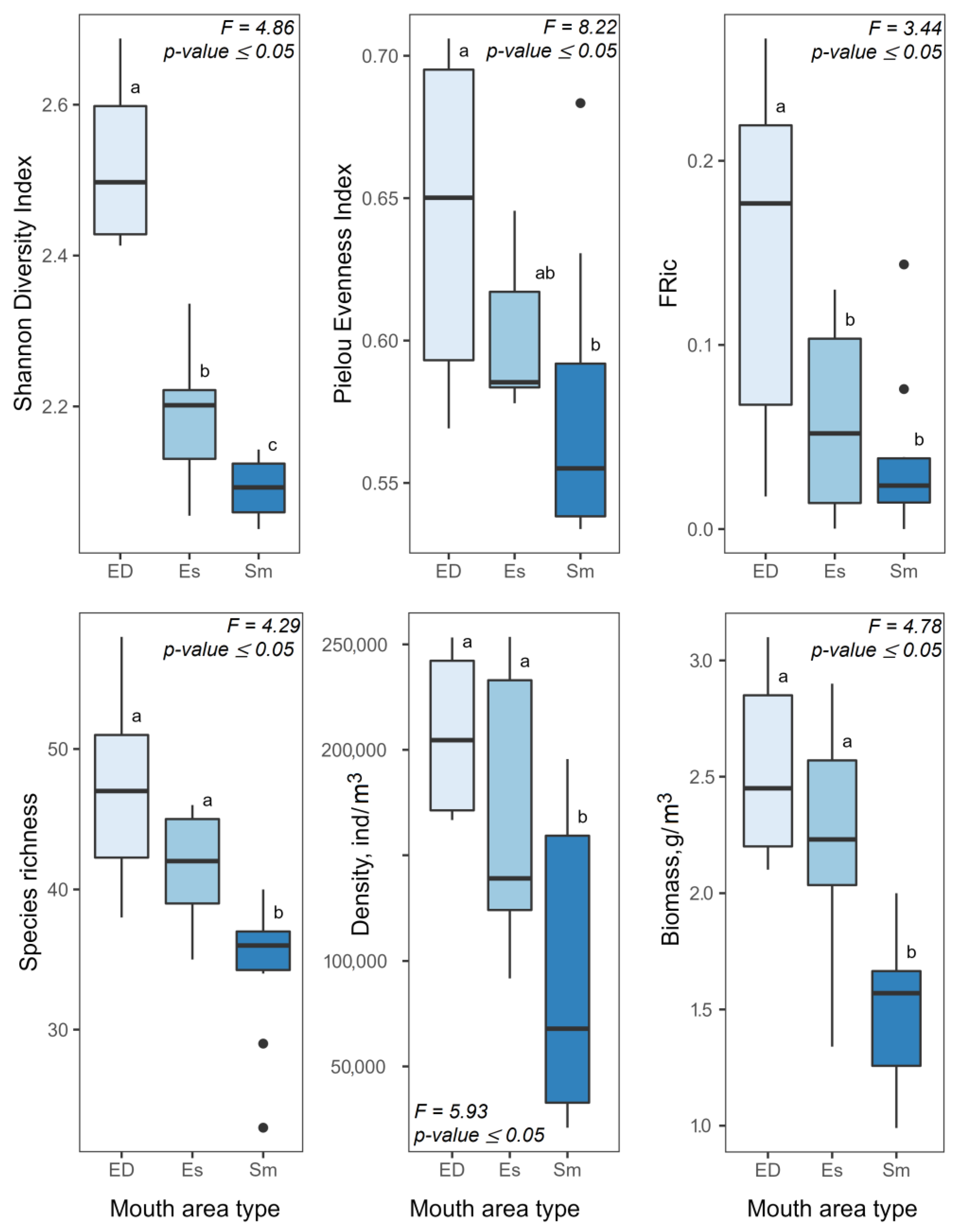
4.1. Structural Organization of Zooplankton Communities in Mouth Areas
4.2. Alien Species of Zooplankton in Mouth Areas
4.3. Environmental Factors Regulating the Ecotone Community
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Novoselov, A.P.; Imant, E.N.; Artem’ev, S.N.; Matveev, N.Y.; Matveeva, A.D. Current State of Planktonic and Benthic Communities in the Mouth Part of the Northern Dvina River. Russ. J. Ecol. 2022, 53, 212–220. [Google Scholar] [CrossRef]
- Zalewski, M.; Frankiewicz, P.; Nowak, M. Biomanipulation by ecotone management in lowland reservoir. Hydrobiologia 1995, 303, 49–60. [Google Scholar] [CrossRef]
- Di Castri, F.; Hansen, A.J.; Holland, M.M. A new look at ecotones: Emerging international projects on landscape boundaries. Biol. Int. 1988, 17, 163. [Google Scholar]
- Hansen, A.J.; Castri, F. Landscape Boundaries; Springer: New York, NY, USA, 1992; p. 452. [Google Scholar]
- Risser, P.G. The status of the science examining ecotones. BioScience 1995, 45, 318–325. [Google Scholar] [CrossRef]
- Lachavanne, J.B.; Juge, R. Biodiversity in Land-Inland Water Ecotones; Taylor & Francis: New York, NY, USA, 1977; p. 308. [Google Scholar]
- Krylov, A.V.; Tsvetkov, A.I.; Malin, M.I.; Romanenko, S.A.; Poddubnii, S.A.; Otjukova, N.G. Communities of hydrobionts and the physical-chemical characteristics of the estuary area of inflow of a flat water basin. Inland Water Biol. 2010, 3, 59–69. [Google Scholar] [CrossRef]
- Bolotov, S.E.; Krylov, A.V.; Eskov, V.M.; Kozlova, V.V.; Mukhortova, O.V. Comparative Analysis of Ecological Structure and Parameters of the Chaotic Organization of Zooplankton in the Outfall of a Tributary of a Flatland Water Reservoir. Proc. Samara Sci. Cent. Russ. Acad. Sci. 2014, 16, 223–226. [Google Scholar]
- Bolotov, S.E.; Tsvetkov, A.I.; Krylov, A.V. Zooplankton in the zones of confluence of unregulated rivers. Inland Water Biol. 2012, 5, 184–191. [Google Scholar] [CrossRef]
- Hydroecology of Lowland Water Reservoir Tributaries’ Mouth; Filigran: Yaroslavl, Russia, 2015; p. 466.
- Naiman, R.J.; Décamps, H.; Pastor, J.; Johnston, C.A. The Potential Importance of Boundaries of Fluvial Ecosystems. J. N. Am. Benthol. Soc. 1988, 7, 289–306. [Google Scholar] [CrossRef]
- Kozuharov, D.; Evtimova, V.; Zaharieva, D. Long-term changes of zooplankton and dynamics of eutrophication in the polluted system of Strouma River—Pchelina Reservoir (South-western Bulgaria). Acta Zool. Bulg. 2007, 59, 191–202. [Google Scholar]
- Traykov, I.; Boyanovsky, B.; Zivkov, M. Composition and abundance of Zooplankton in Kardzhaly reservoir. Bulg. J. Agric. Sci. 2011, 4, 501–511. [Google Scholar]
- Mieczan, T.; Adamczuk, M.; Nawrot, D. Effect of water chemistry on the planktonic communities and relationships among food web components across a freshwater ecotone. Arch. Biol. Sci. 2013, 4, 1491–1504. [Google Scholar] [CrossRef]
- Ward, J.V.; Tockner, K.; Schiemer, F. Biodiversity of floodplain river ecosystems: Ecotones and connectivity. Regul. Rivers Res. Manag. 1999, 15, 125–139. [Google Scholar] [CrossRef]
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams; Oxford University Press: North York, ON, Canada, 1997; p. 400. [Google Scholar]
- Moss, B. Ecology of Freshwater; Wiley-Blackwell: Hoboken, NJ, USA, 1988; p. 352. [Google Scholar]
- Sharma, R.C. Habitat ecology and diversity of freshwater zooplankton of Uttarakhand Himalaya, India. Biodivers. Int. J. 2020, 5, 188–196. [Google Scholar]
- Andronikova, I.N. Structural and Functional Organization of Zooplankton in Lake Ecosystems of Different Trophic Types; Nauka: St. Petersburg, Russia, 1996; p. 189. [Google Scholar]
- Krylov, A.V. Zooplankton of Lowland Small Rivers; Nauka: Moscow, Russia, 2005; p. 263. [Google Scholar]
- Shurganova, G.V. Dynamics of Species Structure of Zooplanktocenoses during Their Formation and Development (by the Example of Reservoirs in the Middle Volga: Gorki and Cheboksary); Extended Abstract of Doctoral (Biol.) Dissertation; Cyberleninka: Nizhny Novgorod, Russia, 2007; p. 48. [Google Scholar]
- Grabowska, M.; Ejsmont-Karabin, J.; Karpowicz, M. Reservoir-river relationships in lowland, shallow, eutrophic systems: An impact of zooplankton from hypertrophic reservoir on river zooplankton. Pol. J. Ecol. 2013, 4, 759–768. [Google Scholar]
- Ejsmont-Karabin, J. Is sandy beach of the lake an ecotone? Psammon rotifera in a mesotrophic Lake Kuc (Masurian Lakeland, Northern Poland). Pol. J. Ecol. 2003, 2, 219–224. [Google Scholar]
- Ilyin, M.Y. Composition and Structure of Zooplankton Communities of Water Bodies of Specially Protected Natural Areas (by Example of the Nizhny Novgorod Region); Extended Abstract of Doctoral (Biol.) Dissertation; Cyberleninka: Nizhny Novgorod, Russia, 2016; p. 36. [Google Scholar]
- Gavrilko, D.E.; Zhikharev, V.S.; Ruchkin, D.S.; Zolotareva, T.V.; Shurganova, G.V. Cladocerans in the higher aquatic plant thickets in European Russia, the inflows of the Gorkovsky and Cheboksarky reservoirs taken as examples. Zoo. J. 2020, 99, 146–156. [Google Scholar]
- Podshivalina, V.N.; Sheveleva, N.G. First record of Sinodiaptomus sarsi (Copepoda: Calanoida) from the East European Plain. Zoosyst. Ross. 2020, 29, 60–69. [Google Scholar] [CrossRef]
- Zhikharev, V.S.; Gavrilko, D.E.; Shurganova, G.V. A record of the tropical species Thermocyclops taihokuensis Harada, 1931 (Copepoda: Cyclopoida) in European Russia. Biol. Bull. 2020, 47, 1347–1350. [Google Scholar] [CrossRef]
- Zhikharev, V.S.; Neretina, A.N.; Zolotareva, T.V.; Gavrilko, D.E.; Shurganova, G.V. Ilyocryptus spinifer Herrick 1882 (Crustacea, Branchiopoda, Cladocera): The first record of the species in the european fauna. Biol. Bull. 2020, 47, 930–936. [Google Scholar] [CrossRef]
- Zhikharev, V.S.; Sinev, A.Y.; Shurganova, G.V. New record of non-indigenous cladoceran Pleuroxus denticulatus Birge, 1879 (Cladocera: Chydoridae) in the European Russia. Invertebr. Biol. 2022, 19, 317–323. [Google Scholar] [CrossRef]
- Alekseev, V.R. Confusing Invader: Acanthocyclops americanus (Copepoda: Cyclopoida) and Its Biological, Anthropogenic and Climate-Dependent Mechanisms of Rapid Distribution in Eurasia. Water 2021, 13, 1423. [Google Scholar] [CrossRef]
- Fefilova, E.B. Records of the Siberian–Beringian Genus Nordodiaptomus (Copepoda, Calanoida) from the European Part of Russia. Biol. Bull. 2020, 48, 1231–1235. [Google Scholar] [CrossRef]
- Kotov, A.A.; Karabanov, D.P.; Van Damme, K. Non-indigenous Cladocera (Crustacea: Branchiopoda): From a few notorious cas58es to a potential global faunal mixing in aquatic ecosystems. Water 2022, 14, 2806. [Google Scholar] [CrossRef]
- Lazareva, V.I.; Zhdanova, S.M.; Sabitova, R.Z. The spread of east asian copepod Thermocyclops taihokuensis (Crustacea, Copepoda) in the Volga River basin. Inland Water Biol. 2022, 15, 139–148. [Google Scholar] [CrossRef]
- Mikhailov, V.N.; Mikhailova, M.V.; Magritsky, D.V. Fundamentals of the Hydrology of River Mouths: A Textbook; Triumf: Moscow, Russia, 2018; p. 314. [Google Scholar]
- Hudec, I. Anomopoda, Ctenopoda, Haplopoda, Onychopoda (Crustacea: Branchiopoda). Fauna Slovenska III; VEDA: Bratislava, Slovakia, 2010; p. 496. [Google Scholar]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe: Cladocera & Copepoda (Calanoida, Cyclopoida) Key to Species Identification, with Notes on Ecology, Distribution, Methods and Introduction to Data Analysis; Springer: Cham, Switzerland, 2016; p. 918. [Google Scholar]
- Rogers, D.C.; Thorp, J.H. Keys to Palaearctic Fauna. Thorp and Covich’s Freshwater. Invertebrates; Academic Press: Oxford, UK, 2019; Volume IV, p. 920. [Google Scholar]
- Korovchinsky, N.M.; Kotov, A.A.; Sinev, A.Y.; Neretina, A.N.; Garibyan, P.G. Cladocera (Crustacea: Cladocera) of Northern Eurasia; KMK Scientific Press Ltd.: Moscow, Russia, 2021; Volume II, p. 544. [Google Scholar]
- Sapozhnikov, V.V.; Agatova, A.I.; Arzhanova, N.V.; Mordasova, N.V.; Lapina, N.M.; Zubarevich, V.L. Guidelines for Chemical Analysis of Marine and Fresh Waters in Environmental Monitoring of Fishery Waters and Fishery Prospective Areas of the World Ocean; VNIRO: Moscow, Russia, 2003; p. 202. [Google Scholar]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Oxford, UK, 2012; p. 990. [Google Scholar]
- Gavrilko, D.E.; Shurganova, G.V.; Kudrin, I.A.; Yakimov, B.N. Identification of Freshwater Zooplankton Functional Groups Based on the Functional Traits of Species. Biol. Bull. 2021, 48, 1849–1856. [Google Scholar] [CrossRef]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comp. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; p. 306. [Google Scholar]
- Yakimov, B.N.; Shurganova, G.V.; Cherepennikov, V.V.; Kudrin, I.A.; Il’in, M.I. Methods for comparative assessment of the results of cluster analysis of hydrobiocenoses structure (by the example of zooplankton communities of the Linda River, Nizhny Novgorod region). Inland Water Biol. 2016, 2, 200–208. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: http://www.R–project.org/ (accessed on 30 November 2022).
- Holland, M.; Risser, P.; Naiman, R. Ecotones; Springer: New York, NY, USA, 1991; p. 142. [Google Scholar]
- Janauer, G.A. Macrophytes, hydrology, and aquatic ecotones: A GIS-supported ecological survey. Aquat. Bot. 1997, 58, 379–391. [Google Scholar] [CrossRef]
- Kent, M.; Gill, W.J.; Weaver, R.E.; Armitage, R.P. Landscape and plant community boundaries in biogeography. Prog. Phys. Geogr. 1997, 21, 315–353. [Google Scholar] [CrossRef]
- Kark, S. Chapter 9. Ecotones and Ecological Gradients/Ecological Systems: Selected Entries from the Encyclopedia of Sustainability Science and Technology; Springer: New York, NY, USA, 2013; pp. 147–160. [Google Scholar]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Bastow-Wilson, J. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Mouillot, D.; Villéger, S.; Scherer-Lorenzen, M.; Mason, N.W.H. Functional Structure of Biological Communities Predicts Ecosystem Multifunctionality. PLoS ONE 2011, 6, e17476. [Google Scholar] [CrossRef]
- Mikhailov, V.N.; Gorin, S.L. New definitions, regionalization, and typification of river mouth areas and estuaries as their parts. Water Resour. 2012, 39, 243–257. [Google Scholar] [CrossRef]
- Shkolny, D.I.; Aibulatov, D.N. Types of River Mouth Sites of the Southern Island (The Novaya Zemlya Archipelago). Vestn. Mosk. Universiteta. Seriya 5 Geogr. 2016, 6, 50–58. [Google Scholar]
- Bhattacharya, J. Deltas and Estuaries. Sedimentology; Springer: Berlin, Germany, 1978; pp. 145–152. [Google Scholar]
- Bianchi, T.S.; Allison, M.A. Large-river delta-front estuaries as natural “recorders” of global environmental change. Proc. Natl. Acad. Sci. USA 2009, 106, 8085–8092. [Google Scholar] [CrossRef] [PubMed]
- Kennish, M.J. Ecology of Estuaries; CRC Press: Boca Raton, FL, USA, 1986; p. 272. [Google Scholar]
- Petchey, O.L. Integrating methods that investigate how complementarity influences ecosystem functioning. Oikos 2003, 101, 323–330. [Google Scholar] [CrossRef]
- Tilman, D. Biodiversity: Population versus ecosystem stability. Ecology 1996, 77, 350–363. [Google Scholar] [CrossRef]
- Ward, J.V.; Wiens, J.A. Ecotones of riverine ecosystems: Role and typology, spatiotemporal dynamics, and river regulation. Ecohyd. Hydrob. 2001, 1, 25–36. [Google Scholar]
- Patrick, C.J. Macroinvertebrate communities of ecotones between the boundaries of streams, wetlands, and lakes. Fundam. Appl. Limnol. 2014, 185, 223–233. [Google Scholar] [CrossRef]
- Huang, X.; Hu, B.; Wang, P.; Chen, X.; Xu, B. Microbial diversity in lake–river ecotone of Poyang Lake, China. Environ. Earth Sci. 2016, 75, 965. [Google Scholar] [CrossRef]
- Toruan, R.L.; Coggins, L.X.; Ghadouani, A. Response of Zooplankton Size Structure to Multiple Stressors in Urban Lakes. Water 2021, 13, 2305. [Google Scholar] [CrossRef]
- Florescu, L.I.; Moldoveanu, M.M.; Catană, R.D.; Păceșilă, I.; Dumitrache, A.; Gavrilidis, A.A.; Iojă, C.I. Assessing the Effects of Phytoplankton Structure on Zooplankton Communities in Different Types of Urban Lakes. Diversity 2022, 14, 231. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, W.; Zhou, Z.; Chen, W. Spatial Differences in Zooplankton Community Structure between Two Fluvial Lakes in the Middle and Lower Reaches of the Yangtze River: Effects of Land Use Patterns and Physicochemical Factors. Diversity 2022, 14, 908. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Czerniawski, R. Trophic state (TSISD) and mixing type significantly influence pelagic zooplankton biodiversity in temperate lakes (NW Poland). PeerJ 2018, 6, e5731. [Google Scholar] [CrossRef]
- Mamun, M.; Kim, J.Y.; An, K. Trophic Responses of the Asian Reservoir to Long-Term Seasonal and Interannual Dynamic Monsoon. Water 2020, 12, 2066. [Google Scholar] [CrossRef]
- Lu, X.; Liu, Y.; Fan, Y. Diatom Taxonomic Composition as a Biological Indicator of the Ecological Health and Status of a River Basin under Agricultural Influence. Water 2020, 12, 2067. [Google Scholar] [CrossRef]
- Xia, M.; Xiong, F.; Li, X.; Li, D.; Wang, Z.; Zhai, D.; Liu, H.; Chen, Y.; Yu, J.; Wang, Y. Different Assembly Patterns of Planktonic and Sedimentary Bacterial Community in a Few Connected Eutrophic Lakes. Water 2022, 14, 723. [Google Scholar] [CrossRef]
- Rajwa, A.; Bialik, R.J.; Karpiński, M.; Luks, B. Dissolved Oxygen in Rivers: Concepts and Measuring Techniques; Springer: Cham, Switzerland, 2014; pp. 337–350. [Google Scholar]
- Kruzhylina, S.V.; Buzevych, I.Y.; Rudyk-Leuska, N.Y.; Khyzhniak, M.I.; Didenko, A.V. Changes in the structure and dominance of the zooplankton communityof the Kremenchuk Reservoir under the effect of climate changes. Biosystems Diversity 2021, 29, 217–224. [Google Scholar] [CrossRef]
- Banerjee, A.; Rakshit, N.; Chakrabarty, M.; Sinha, S.; Ghosh, S.; Ray, S. Zooplankton community of Bakreswar reservoir: Assessment and visualization of distribution pattern using self-organizing maps. Ecol. Inform. 2022, 72, 101837. [Google Scholar] [CrossRef]
- Xiong, M.; Li, R.; Zhang, T.; Liao, C.; Yu, G.; Yuan, J.; Liu, J.; Ye, S. Zooplankton Compositions in the Danjiangkou Reservoir, a Water Source for the South-to-North Water Diversion Project of China. Water 2022, 14, 3253. [Google Scholar] [CrossRef]
- Phan, N.; Duong, Q.H.; Tran-Nguyen, Q.A.; Trinh-Dang, M. The Species Diversity of Tropical Freshwater Rotifers (Rotifera: Monogononta) in Relation to Environmental Factors. Water 2021, 13, 1156. [Google Scholar] [CrossRef]
- Ko, E.; Kim, D.; Jung, E.; Heo, Y.; Joo, G.; Kim, H. Comparison of Zooplankton Community Patterns in Relation to Sediment Disturbances by Dredging in the Guemho River, Korea. Water 2020, 12, 3434. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Bodies | Length, km | Basin Area, km2 | Mouth Area/Reservoir Type |
|---|---|---|---|
| Uzola River | 147 | 1920 | simple (Sm) |
| Sura River | 841 | 67,500 | simple (Sm) |
| Bol’shoj Civil’ River | 172 | 4690 | simple (Sm) |
| Mera River | 152 | 2380 | estuarine (Es) |
| Troca River | 22 | 217 | estuarine (Es) |
| Vetluga River | 889 | 39,400 | estuarine (Es) |
| Shirmaksha River | 28 | 205 | estuary–deltaic (ED) |
| Sviyaga River | 375 | 16,700 | estuary–deltaic (ED) |
| Gorky Reservoir | 440 | 1590 | lowland |
| Cheboksary Reservoir | 341 | 2190 | lowland |
| Kuybyshev Reservoir | 500 | 6450 | lowland |
| Alien Species | Community 1 | Community 2 | Community 3 | Community 4 |
|---|---|---|---|---|
| Kellicottia bostoniensis (Rousselet, 1908) | 10.0 ± 7.0 | 10.0 ± 1.0 | – | – |
| Keratella tropica (Apstein, 1907) | 50.0 | 10.0 ± 2.0 | – | – |
| Acanthocyclops americanus (Marsh, 1893) | 1040.0 ± 1000.0 | 2230 ± 460.0 | 1380.0 ± 70.0 | 230.0 ± 90.0 |
| Thermocyclops taihokuensis (Harada, 1931) | 2910.0 ± 820.0 | 8560.0 ± 2230.0 | 4780.0 ± 390.0 | 20.0 ± 4.0 |
| Thermocyclops vermifer (Lindberg, 1960) | 10.0 ± 2.0 | 1130.0 ± 300.0 | 10.0 | 20.0 |
| Eurytemora caspica (Sukhikh & Alekseev, 2013) | – | – | – | 30.0 ± 20.0 |
| Eurytemora velox (Lilljeborg, 1853) | – | 40.0 ± 10.0 | 0.05 | – |
| Heterocope caspia (Sars G.O., 1897) | – | 10.0 | – | 70.0 ± 30.0 |
| Factor | Sm | Es | ED |
|---|---|---|---|
| Chlorophyll-a, mg/L (Chl_a) | 11.48 ± 3.00 | 12.73 ± 0.71 | 10.37 ± 1.97 |
| Dissolved oxygen, mg/L (DO) | 7.30 ± 0.63 | 11.12 ± 1.00 | 9.25 ± 1.18 |
| pH | 8.80 ± 0.12 | 8.22 ± 0.17 | 8.56 ± 0.02 |
| Water temperature, °C (WT) | 24.75 ± 0.29 | 25.72 ± 0.35 | 24.65 ± 0.37 |
| Electrical conductivity, μS/cm (EC) | 480.40 ± 90.01 | 245.50 ± 16.32 | 508.00 ± 181.85 |
| Water transparency, m (SD) | 1.14 ± 0.13 | 1.08 ± 0.10 | 0.78 ± 0.09 |
| Total phosphorus, µg/L (TP) | 0.23 ± 0.05 | 0.08 ± 0.003 | 0.24 ± 0.05 |
| Trophic state index (TSI) | 55.58 ± 4.65 | 52.65 ± 0.53 | 59.40 ± 2.79 |
| Factor | R2 | Pseudo-F | P |
|---|---|---|---|
| Water transparency (SD) | 9.77% | 7.80 | 0.001 * |
| Dissolved oxygen (DO) | 10.33% | 7.99 | 0.003 * |
| Trophic state index (TSI) | 5.08% | 1.36 | 0.024 * |
| Electrical conductivity (EC) | 21.07% | 8.91 | 0.001 * |
| Water temperature (WT) | 4.10% | 5.24 | 0.048 * |
| pH | 6.93% | 7.60 | 0.008 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhikharev, V.; Gavrilko, D.; Kudrin, I.; Vodeneeva, E.; Erina, O.; Tereshina, M.; Shurganova, G. Structural Organization of Zooplankton Communities in Different Types of River Mouth Areas. Diversity 2023, 15, 199. https://doi.org/10.3390/d15020199
Zhikharev V, Gavrilko D, Kudrin I, Vodeneeva E, Erina O, Tereshina M, Shurganova G. Structural Organization of Zooplankton Communities in Different Types of River Mouth Areas. Diversity. 2023; 15(2):199. https://doi.org/10.3390/d15020199
Chicago/Turabian StyleZhikharev, Vyacheslav, Dmitry Gavrilko, Ivan Kudrin, Ekaterina Vodeneeva, Oksana Erina, Maria Tereshina, and Galina Shurganova. 2023. "Structural Organization of Zooplankton Communities in Different Types of River Mouth Areas" Diversity 15, no. 2: 199. https://doi.org/10.3390/d15020199
APA StyleZhikharev, V., Gavrilko, D., Kudrin, I., Vodeneeva, E., Erina, O., Tereshina, M., & Shurganova, G. (2023). Structural Organization of Zooplankton Communities in Different Types of River Mouth Areas. Diversity, 15(2), 199. https://doi.org/10.3390/d15020199