
Normalized Difference Vegetation Index as a Proxy of Urban Bird Species Presence and Distribution at Different Spatial Scales

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Bird Surveys
2.3. NDVI Estimation
2.4. Data Analysis
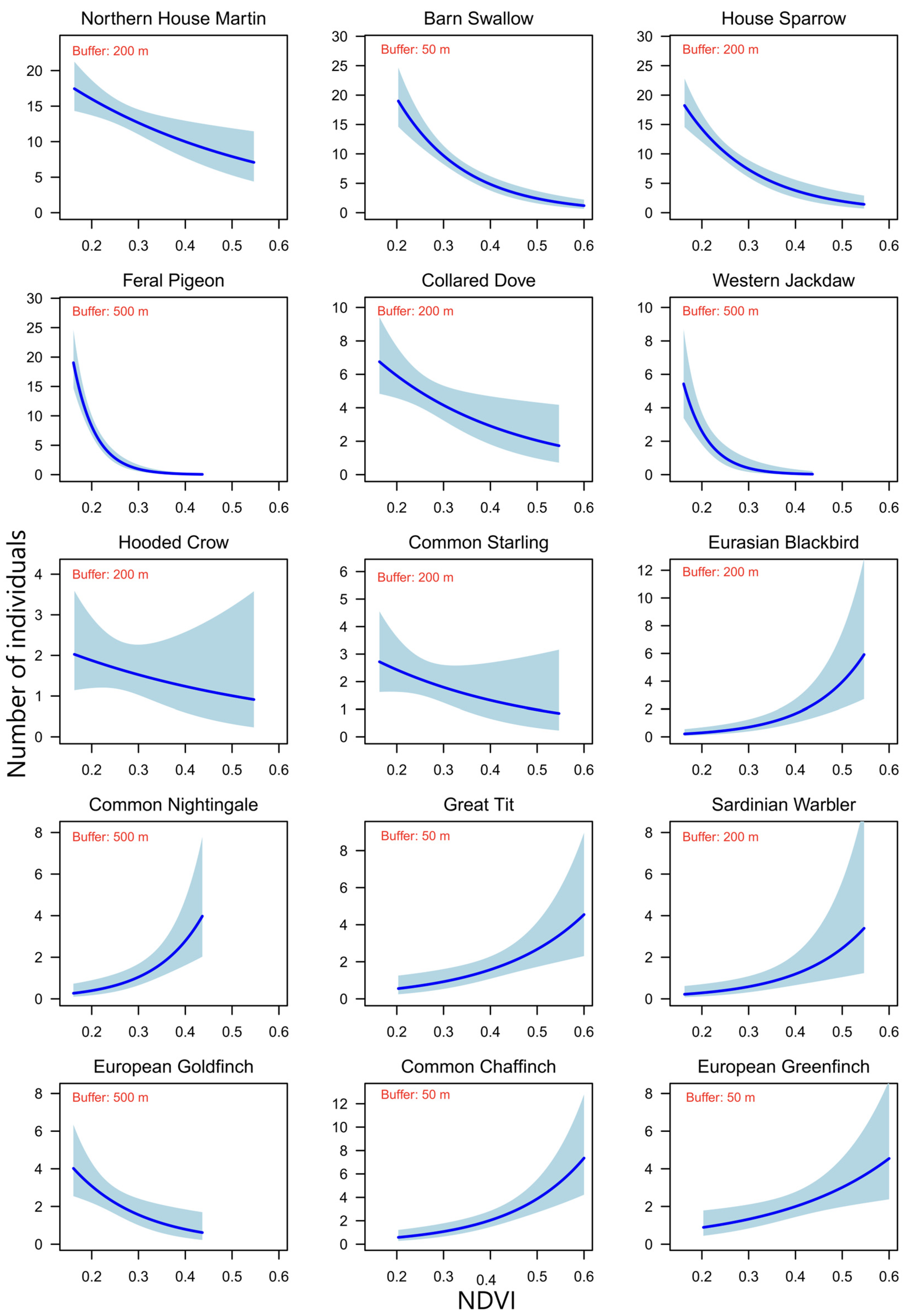
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Urbanization Prospects: The 2018 Revision (ST/ESA/SER.A/420). United Nations. Available online: https://population.un.org/wup/Publications/Files/WUP2018-Report.pdf (accessed on 20 October 2023).
- Liordos, V.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Valsamidis, E.; Kontsiotis, V.J. Niche analysis and conservation of bird species using urban core areas. Sustainability 2021, 13, 6327. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.G.; Bai, X.M.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Kontsiotis, V.J.; Valsamidis, E.; Liordos, V. Organization and differentiation of breeding bird communities across a forested to urban landscape. Urban For. Urban Green. 2019, 38, 242–250. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.-L. Urban core areas are important for species conservation: A European-level analysis of breeding bird species. Landsc. Urban Plan. 2018, 178, 73–81. [Google Scholar] [CrossRef]
- Tzortzakaki, O.; Kati, V.; Kassara, C.; Tietze, D.T.; Giokas, S. Seasonal patterns of urban bird diversity in a Mediterranean coastal city: The positive role of open green spaces. Urban Ecosyst. 2018, 21, 27–39. [Google Scholar] [CrossRef]
- Liordos, V.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Valsamidis, E.; Kontsiotis, V.J. Patch, matrix and disturbance variables negatively influence bird community structure in small-sized managed green spaces located in urban core areas. Sci. Total Environ. 2021, 801, 149617. [Google Scholar] [CrossRef] [PubMed]
- Cohn, J.P. Urban wildlife. BioScience 2005, 55, 201–205. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Warren, P.S. Urban Bird Ecology and Conservation; University of California Press: Berkeley, CA, USA, 2012. [Google Scholar]
- Xu, X.; Xie, Y.; Qi, K.; Luo, Z.; Wang, X. Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci. Total Environ. 2018, 624, 1561–1576. [Google Scholar] [CrossRef]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 7, 46–69. [Google Scholar] [CrossRef]
- Pellissier, V.; Cohen, M.; Boulay, A.; Clergeau, P. Birds are also sensitive to landscape composition and configuration within the city centre. Landsc. Urban Plan. 2012, 104, 181–188. [Google Scholar] [CrossRef]
- Fischer, J.D.; Schneider, C.; Ahlers, A.A.; Miller, J.R. Categorizing wildlife responses to urbanization and conservation implications of terminology. Conserv. Biol. 2015, 29, 1246–1248. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.T.C.; Shanahan, D.F.; Hudson, H.L.; Fuller, R.A.; Gaston, K.J. The impact of urbanization on nature dose and the implications for human health. Landsc. Urban Plan. 2018, 179, 72–80. [Google Scholar] [CrossRef]
- Soga, M.; Gaston, K.J. Extinction of experience: The loss of human–nature interactions. Front. Ecol. Environ. 2016, 14, 94–101. [Google Scholar] [CrossRef]
- Fuller, R.A.; Irvine, K.N.; Devine-Wright, P.; Warren, P.H.; Gaston, K.J. Psychological benefits of green space increase with biodiversity. Biol. Lett. 2007, 3, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Lerman, S.B.; Warren, P.S. The conservation value of residential yards: Linking birds and people. Ecol. Appl. 2011, 21, 1327–1339. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.W.F.; Brindley, P.; Mears, M.; McEwan, K.; Ferguson, F.; Sheffield, D.; Jorgensen, A.; Riley, J.; Goodrick, J.; Ballard, L.; et al. Where the wild things are! Do urban green spaces with greater avian biodiversity promote more positive emotions in humans? Urban Ecosyst. 2020, 23, 301–317. [Google Scholar] [CrossRef]
- Shwartz, A.; Turbé, A.; Simon, L.; Julliard, R. Enhancing urban biodiversity and its influence on city-dwellers: An experiment. Biol. Conserv. 2014, 171, 82–90. [Google Scholar] [CrossRef]
- Wolf, L.J.; Zu Ermgassen, S.; Balmford, A.; White, M.; Weinstein, N. Is variety the spice of life? An experimental investigation into the effects of species richness on self-reported mental well-being. PLoS ONE 2017, 12, e0170225. [Google Scholar] [CrossRef]
- Ferenc, M.; Sedláček, O.; Fuchs, R. How to improve urban greenspace for woodland birds: Site and local-scale determinants of bird species richness. Urban Ecosyst. 2014, 17, 625–640. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Morelli, F.; Mikula, P.; Krištín, A.; Indykiewicz, P.; Grzywaczewski, G.; Kronenberg, J.; Jerzak, L. Bird diversity in urban green space: A large-scale analysis of differences between parks and cemeteries in Central Europe. Urban For. Urban Green. 2017, 27, 264–271. [Google Scholar] [CrossRef]
- Leveau, L.M.; Federico, I.; Isla, F.I. Predicting bird species presence in urban areas with NDVI: An assessment within and between cities. Urban For. Urban Green. 2021, 63, 127199. [Google Scholar] [CrossRef]
- Rocchini, D.; Balkenhol, N.; Carter, G.A.; Foody, G.M.; Gillespie, T.W.; He, K.S.; Kark, S.; Levin, N.; Lucas, K.; Luoto, M.; et al. Remotely sensed spectral heterogeneity as a proxy of species diversity: Recent advances and open challenges. Ecol. Inform. 2010, 5, 318–329. [Google Scholar] [CrossRef]
- Rocchini, D.; Boyd, D.S.; Féret, J.B.; Foody, G.M.; He, K.S.; Lausch, A.; Nagendra, H.; Wegmann, M.; Pettorelli, N. Satellite remote sensing to monitor species diversity: Potential and pitfalls. Remote Sens. Ecol. Conserv. 2016, 2, 25–36. [Google Scholar] [CrossRef]
- Wang, H.; Zeng, Z.-C. Satellite observations of urban greenery phenology in downtown Beijing at meter to kilometer scales. In Proceedings of the IGARSS 2022–2022, Kuala Lumpur, Malaysia, IEEE International Geoscience and Remote Sensing Symposium, Kuala Lumpur, Malaysia, 17–22 July 2022; pp. 6268–6271. [Google Scholar] [CrossRef]
- Xofis, P.; Spiliotis, J.A.; Chatzigiovanakis, S.; Chrysomalidou, A.S. Long-term monitoring of vegetation Dynamics in the Rhodopi Mountain Range National Park-Greece. Forests 2022, 13, 377. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L.J.; Hupy, P.; Wang, Y.; Shao, G. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2021, 3, 1–6. [Google Scholar] [CrossRef]
- Zhengxing, W.; Chuang, L.; Alfredo, H. From AVHRR-NDVI to MODIS-EVI: Advances in vegetation index research. Acta Ecol. Sin. 2003, 23, 979–987. [Google Scholar]
- Anguluri, R.; Narayanan, P. Role of green space in urban planning: Outlook towards smart cities. Urban For. Urban Green. 2017, 25, 58–65. [Google Scholar] [CrossRef]
- Li, X.; Gong, P.; Liang, L. A 30-year (1984–2013) record of annual urban dynamics of Beijing City derived from Landsat data. Remote Sens. Environ. 2015, 166, 78–90. [Google Scholar] [CrossRef]
- Jasmani, Z.; Ravn, H.P.; van den Bosch, C.C.K. The influence of small urban parks characteristics on bird diversity: A case study of Petaling Jaya, Malaysia. Urban Ecosyst. 2017, 20, 227–243. [Google Scholar] [CrossRef]
- Strohbach, M.W.; Lerman, S.B.; Warren, P.S. Are small greening areas enhancing bird diversity? Insights from community-driven greening projects in Boston. Landsc. Urban Plan. 2013, 114, 69–79. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Schondube, J.E. Gray vs. green urbanization: Relative importance of urban features for urban bird communities. Basic Appl. Ecol. 2011, 12, 372–381. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A metaanalysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Bino, G.; Levin, N.; Darawshi, S.; Van Der Hal, N.; Reich-Solomon, A.; Kark, S. Accurate prediction of bird species richness patterns in an urban environment using Landsat-derived NDVI and spectral unmixing. Int. J. Remote Sens. 2008, 29, 3675–3700. [Google Scholar] [CrossRef]
- Cooper, W.J.; McShea, W.J.; Forrester, T.; Luther, D.A. The value of local habitat heterogeneity and productivity when estimating avian species richness and species of concern. Ecosphere 2020, 11, e03107. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Major, R.E.; Lyons, M.B.; Martin, J.M.; Kingsford, R.T. The effects of local and landscape habitat attributes on bird diversity in urban greenspaces. Ecosphere 2018, 9, e02347. [Google Scholar] [CrossRef]
- Nieto, S.; Flombaum, P.; Garbulsky, M.F. Can temporal and spatial NDVI predict regional bird-species richness? Glob. Ecol. Conserv. 2015, 3, 729–735. [Google Scholar] [CrossRef]
- Katoh, K.; Matsuba, M. Effectiveness of nature reserves for bird conservation in urban parks in Tokyo. J. For. Res. 2021, 32, 2011–2022. [Google Scholar] [CrossRef]
- Leveau, L.M. Primary productivity and habitat diversity predict bird species richness and composition along urban-rural gradients of central Argentina. Urban For. Urban Green. 2019, 43, 126349. [Google Scholar] [CrossRef]
- Leveau, L.M.; Isla, F.I.; Bellocq, M.I. Predicting the seasonal dynamics of bird communities along an urban-rural gradient using NDVI. Landsc. Urban Plan. 2018, 177, 103–113. [Google Scholar] [CrossRef]
- Leveau, L.M.; Isla, F.I.; Bellocq, M.I. From Town to Town: Predicting the Taxonomic, Functional and Phylogenetic Diversity of Birds Using NDVI. Ecol. Indic. 2020, 119, 106703. [Google Scholar] [CrossRef]
- Liordos, V.; Foutsa, E.; Kontsiotis, V.J. Differences in encounters, likeability and desirability of wildlife species among residents of a Greek city. Sci. Total Environ. 2020, 739, 139892. [Google Scholar] [CrossRef] [PubMed]
- Greek Statistical Authority. Population Census 2011. 2011. Available online: http://www.statistics.gr/portal/page/portal/ESYE/PAGE-census2011 (accessed on 20 October 2023).
- Bibby, C.; Burgess, N.; Hill, D.; Mustoe, S. Bird Census Techniques, 3rd ed.; Academic: London, UK, 2000. [Google Scholar]
- Ferry, C.; Frochot, B. L’avifaune nidificatrice d’une foret de chenes pedoncules en Bourgogne: Etude de deux successions ecologiques. Terre Vie 1970, 2, 153–250. Available online: https://www.persee.fr/doc/revec_0040-3865_1970_num_24_2_4591 (accessed on 4 November 2023). [CrossRef]
- USGS. NDVI, the Foundation for Remote Sensing Phenology. 2018. Available online: https://www.usgs.gov/special-topics/remote-sensing-phenology/science/ndvi-foundation-remote-sensing-phenology (accessed on 20 October 2023).
- Christidis, N.; Jones, G.S.; Stott, P.A. Dramatically increasing chance of extremely hot summers since the 2003 European heatwave. Nat. Clim. Chang. 2015, 5, 46–50. [Google Scholar] [CrossRef]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P.; et al. Sentinel-2: ESA’s optical high-resolution mission for GMES operational services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Snow, D.W.; Perrins, C.M. The Birds of the Western Palearctic.; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 October 2023).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference based on (Q)AIC(c): R Package Version 2.0-4. 2016. Available online: http://CRAN.R-project.org/package=AICcmodavg (accessed on 20 October 2023).
- Covas, R.; Blondel, J. Biogeography and history of the Mediterranean bird fauna. Ibis 1998, 140, 395–407. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; La Sorte, F.A.; Aronson, M.F.; Goddard, M.A.; MacGregor-Fors, I.; Nilon, C.H.; Warren, P.S. Global patterns and drivers of urban bird diversity. In Ecology and Conservation of Birds in Urban Environments; Murgui, H., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 13–33. [Google Scholar]
- La Sorte, F.A.; Tingley, M.W.; Hurlbert, A.H. The role of urban and agricultural areas during avian migration: An assessment of within-year temporal turnover. Glob. Ecol. Biogeogr. 2014, 23, 1225–1234. [Google Scholar] [CrossRef]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L.; et al. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.L. Urbanization and species occupancy frequency distribution patterns in core zone areas of European towns. Eur. J. Ecol. 2016, 2, 23–43. [Google Scholar] [CrossRef]
- Ortega-Álvarez, R.; MacGregor-Fors, I. Living in the big city: Effects of land-use on bird community structure, diversity, and composition. Landsc. Urban Plan. 2009, 90, 189–195. [Google Scholar] [CrossRef]
- Pereira, H.M.; Daily, G.C.; Roughgarden, J. A framework for assessing the relative vulnerability of species to land-use change. Ecol. Appl. 2004, 14, 730–742. [Google Scholar] [CrossRef]
- Savard, J.P.L.; Clergeau, P.; Mennechez, G. Biodiversity concepts and urban ecosystems. Landsc. Urban Plan. 2000, 48, 131–142. [Google Scholar] [CrossRef]
- Donnelly, R.; Marzluff, J.M. Importance of reserve size and landscape context to urban bird conservation. Conserv. Biol. 2004, 18, 733–745. [Google Scholar] [CrossRef]
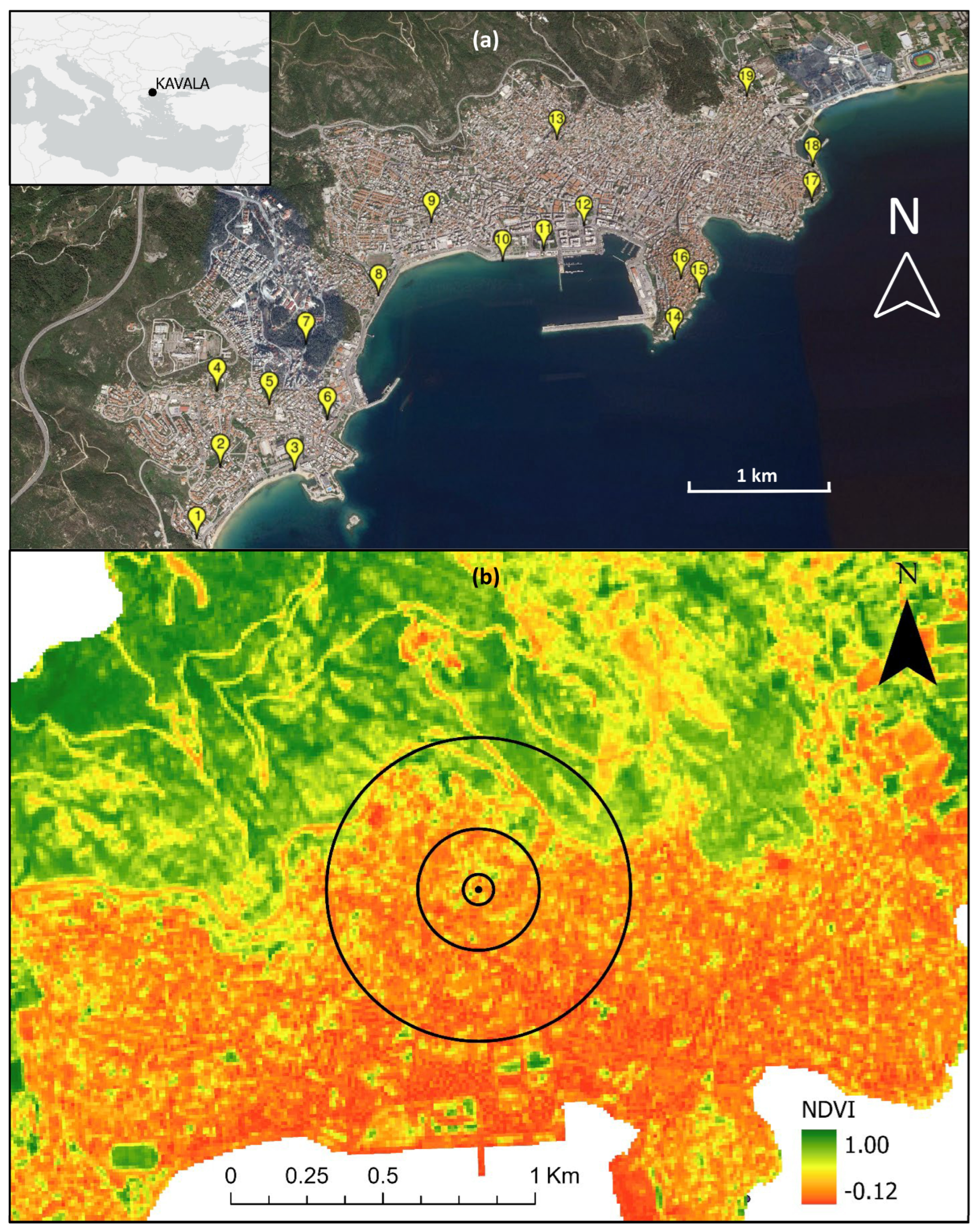

{kind=link}
{kind=link}
| Mean NDVI Values | |||||
|---|---|---|---|---|---|
| Green Space Number | Green Space Type | Number of Species | 50 m Buffer | 200 m Buffer | 500 m Buffer |
| 1 | Square garden | 12 | 0.367 | 0.372 | 0.428 |
| 2 | Woodland | 11 | 0.399 | 0.266 | 0.294 |
| 3 | Playground | 10 | 0.355 | 0.228 | 0.242 |
| 4 | Street hedgerow | 12 | 0.376 | 0.207 | 0.285 |
| 5 | Woodland | 9 | 0.522 | 0.274 | 0.308 |
| 6 | Playground | 10 | 0.303 | 0.321 | 0.302 |
| 7 | Woodland | 9 | 0.600 | 0.547 | 0.349 |
| 8 | Woodland | 11 | 0.437 | 0.347 | 0.323 |
| 9 | Square garden | 5 | 0.301 | 0.202 | 0.223 |
| 10 | Playground | 10 | 0.236 | 0.180 | 0.168 |
| 11 | Square garden | 8 | 0.302 | 0.166 | 0.164 |
| 12 | Square garden | 9 | 0.454 | 0.163 | 0.161 |
| 13 | Street hedgerow | 6 | 0.260 | 0.276 | 0.229 |
| 14 | Street hedgerow | 9 | 0.423 | 0.282 | 0.209 |
| 15 | Playground | 1 | 0.255 | 0.272 | 0.199 |
| 16 | Square garden | 7 | 0.284 | 0.230 | 0.283 |
| 17 | Woodland | 11 | 0.495 | 0.289 | 0.218 |
| 18 | Street hedgerow | 6 | 0.203 | 0.266 | 0.254 |
| 19 | Street hedgerow | 11 | 0.345 | 0.323 | 0.436 |
| Common Name | Mean | SE | Incidence (%) | Urbanization Category |
|---|---|---|---|---|
| Northern House Martin (Delichon urbicum) | 13.684 | 1.355 | 94.7 | Dweller |
| Βarn Swallow (Hirundo rustica) | 7.737 | 3.159 | 47.4 | Dweller |
| House Sparrow (Passer domesticus) | 9.895 | 1.832 | 89.5 | Dweller |
| Feral Pigeon (Columba livia forma urbana) | 5.053 | 2.374 | 52.6 | Dweller |
| Collared Dove (Streptopelia decaocto) | 4.737 | 0.611 | 89.5 | Dweller/Utilizer |
| Western Jackdaw (Corvus monedula) | 1.579 | 0.735 | 31.6 | Dweller |
| Hooded Crow (Corvus cornix) | 1.632 | 0.317 | 68.4 | Utilizer |
| Common Starling (Sturnus vulgaris) | 2.000 | 0.688 | 42.1 | Dweller/Utilizer |
| Eurasian Blackbird (Turdus merula) | 0.842 | 0.369 | 31.6 | Utilizer |
| Common Nightingale (Luscinia megarhynchos) | 1.053 | 0.346 | 36.8 | Utilizer |
| Great Tit (Parus major) | 1.526 | 0.353 | 68.4 | Utilizer |
| Sardinian Warbler (Curruca melanocephala) | 0.632 | 0.256 | 26.3 | Avoider |
| European Goldfinch (Carduelis carduelis) | 2.211 | 0.436 | 68.4 | Utilizer |
| Common Chaffinch (Fringilla coelebs) | 2.053 | 0.575 | 57.9 | Utilizer |
| European Greenfinch (Chloris chloris) | 1.895 | 0.404 | 73.7 | Utilizer |
| Common Name | Estimate | SE | z | AICc | ΔAICc | wi |
|---|---|---|---|---|---|---|
| Northern House Martin | ||||||
| NDVI50 | 0.055 | 0.605 | 0.091 | 143.72 | 8.85 | 0.01 |
| NDVI200 | −2.350 | 0.825 | –2.849 *** | 134.86 | 0.00 | 0.97 |
| NDVI500 | –0.681 | 0.812 | –0.838 | 143.01 | 8.15 | 0.02 |
| Barn Swallow | ||||||
| NDVI50 | –6.957 | 1.028 | –6.765 *** | 304.96 | 0.00 | 1 |
| NDVI200 | −0.87 | 1.010 | −0.861 | 357.57 | 52.61 | 0.00 |
| NDVI500 | 1.604 | 1.022 | 1.570 | 359.21 | 54.24 | 0.00 |
| House Sparrow | ||||||
| NDVI50 | −4.189 | 0.817 | −5.126 *** | 159.73 | 10.29 | 0.01 |
| NDVI200 | −6.646 | 1.164 | −5.709 *** | 149.44 | 0.00 | 0.99 |
| NDVI500 | −4.792 | 1.076 | −4.453 *** | 166.94 | 17.49 | 0.00 |
| Feral Pigeon | ||||||
| NDVI50 | 4.148 | 0.925 | 4.484 *** | 246.52 | 99.05 | 0.00 |
| NDVI200 | −19.721 | 2.889 | −8.617 *** | 157.71 | 10.25 | 0.01 |
| NDVI500 | −21.501 | 2.52 | −8.531 *** | 147.46 | 0.00 | 0.99 |
| Collared Dove | ||||||
| NDVI50 | −0.487 | 1.0431 | −0.467 | 98.80 | 6.22 | 0.04 |
| NDVI200 | −3.550 | 1.488 | −2.385 * | 92.57 | 0.00 | 0.92 |
| NDVI500 | −0.567 | 1.376 | −0.412 | 98.84 | 6.27 | 0.04 |
| Western Jackdaw | ||||||
| NDVI50 | −4.470 | 2.067 | −2.163 * | 105.93 | 26.25 | 0.00 |
| NDVI200 | −13.475 | 3.502 | −3.848 *** | 91.74 | 12.06 | 0.00 |
| NDVI500 | −18.912 | 4.174 | −4.531 *** | 79.68 | 0.00 | 1 |
| Hooded Crow | ||||||
| NDVI50 | 0.586 | 1.727 | 0.339 | 68.30 | 0.73 | 0.28 |
| NDVI200 | −2.076 | 2.355 | −0.882 | 67.57 | 0.00 | 0.41 |
| NDVI500 | −1.25 | 2.388 | −0.524 | 68.13 | 0.56 | 0.31 |
| Common Starling | ||||||
| NDVI50 | −0.513 | 1.606 | −0.320 | 107.55 | 1.95 | 0.20 |
| NDVI200 | −3.033 | 2.234 | −1.358 | 105.59 | 0.00 | 0.53 |
| NDVI500 | 1.791 | 2.001 | 0.895 | 106.87 | 1.28 | 0.28 |
| Eurasian Blackbird | ||||||
| NDVI50 | 5.960 | 2.256 | 2.641 ** | 57.34 | 10.54 | 0.01 |
| NDVI200 | 8.651 | 1.888 | 4.583 *** | 46.81 | 0.00 | 0.97 |
| NDVI500 | 8.898 | 2.894 | 3.075 ** | 54.92 | 8.11 | 0.02 |
| Common Nightingale | ||||||
| NDVI50 | 3.651 | 2.036 | 1.793 | 63.63 | 10.91 | 0.00 |
| NDVI200 | 6.151 | 1.811 | 3.396 *** | 57.34 | 4.62 | 0.09 |
| NDVI500 | 9.782 | 2.606 | 3.754 *** | 52.72 | 0.00 | 0.91 |
| Great Tit | ||||||
| NDVI50 | 5.298 | 1.675 | 3.163 ** | 59.92 | 0.00 | 0.89 |
| NDVI200 | 4.203 | 1.661 | 2.532 * | 64.20 | 4.28 | 0.10 |
| NDVI500 | 2.029 | 2.279 | 0.890 | 68.87 | 8.95 | 0.01 |
| Sardinian Warbler | ||||||
| NDVI50 | 4.525 | 2.610 | 1.734 | 48.20 | 5.18 | 0.04 |
| NDVI200 | 7.122 | 2.55 | 3.158 ** | 43.02 | 0.00 | 0.59 |
| NDVI500 | 9.009 | 3.344 | 2.694 ** | 43.99 | 0.97 | 0.36 |
| European Goldfinch | ||||||
| NDVI50 | 1.803 | 1.444 | 1.248 | 80.90 | 7.60 | 0.02 |
| NDVI200 | −2.832 | 2.104 | −1.346 | 80.40 | 7.11 | 0.03 |
| NDVI500 | −6.813 | 2.425 | −2.809 ** | 73.29 | 0.00 | 0.95 |
| Common Chaffinch | ||||||
| NDVI50 | 6.397 | 1.448 | 4.419 *** | 74.84 | 0.00 | 0.98 |
| NDVI200 | 5.128 | 1.361 | 3.768 *** | 82.30 | 7.46 | 0.02 |
| NDVI500 | 3.523 | 1.912 | 1.842 | 90.81 | 15.98 | 0.00 |
| European Greenfinch | ||||||
| NDVI50 | 4.109 | 1.511 | 2.719 ** | 68.75 | 0.00 | 0.78 |
| NDVI200 | 3.451 | 1.558 | 2.216 * | 71.62 | 2.86 | 0.18 |
| NDVI500 | 2.400 | 2.031 | 1.182 | 74.54 | 5.78 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontsiotis, V.J.; Chatzigiovanakis, S.; Valsamidis, E.; Xofis, P.; Liordos, V. Normalized Difference Vegetation Index as a Proxy of Urban Bird Species Presence and Distribution at Different Spatial Scales. Diversity 2023, 15, 1139. https://doi.org/10.3390/d15111139
Kontsiotis VJ, Chatzigiovanakis S, Valsamidis E, Xofis P, Liordos V. Normalized Difference Vegetation Index as a Proxy of Urban Bird Species Presence and Distribution at Different Spatial Scales. Diversity. 2023; 15(11):1139. https://doi.org/10.3390/d15111139
Chicago/Turabian StyleKontsiotis, Vasileios J., Stavros Chatzigiovanakis, Evangelos Valsamidis, Panteleimon Xofis, and Vasilios Liordos. 2023. "Normalized Difference Vegetation Index as a Proxy of Urban Bird Species Presence and Distribution at Different Spatial Scales" Diversity 15, no. 11: 1139. https://doi.org/10.3390/d15111139
APA StyleKontsiotis, V. J., Chatzigiovanakis, S., Valsamidis, E., Xofis, P., & Liordos, V. (2023). Normalized Difference Vegetation Index as a Proxy of Urban Bird Species Presence and Distribution at Different Spatial Scales. Diversity, 15(11), 1139. https://doi.org/10.3390/d15111139