
A Rare Fish Amphistome Revisited: The Phylogenetic Position of Kalitrema kalitrema (Trematoda: Cladorchiidae) Found in Hypostomus spp. (Siluriformes: Loricariidae) from Brazil

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Morphological Data
2.3. Molecular Data
3. Results
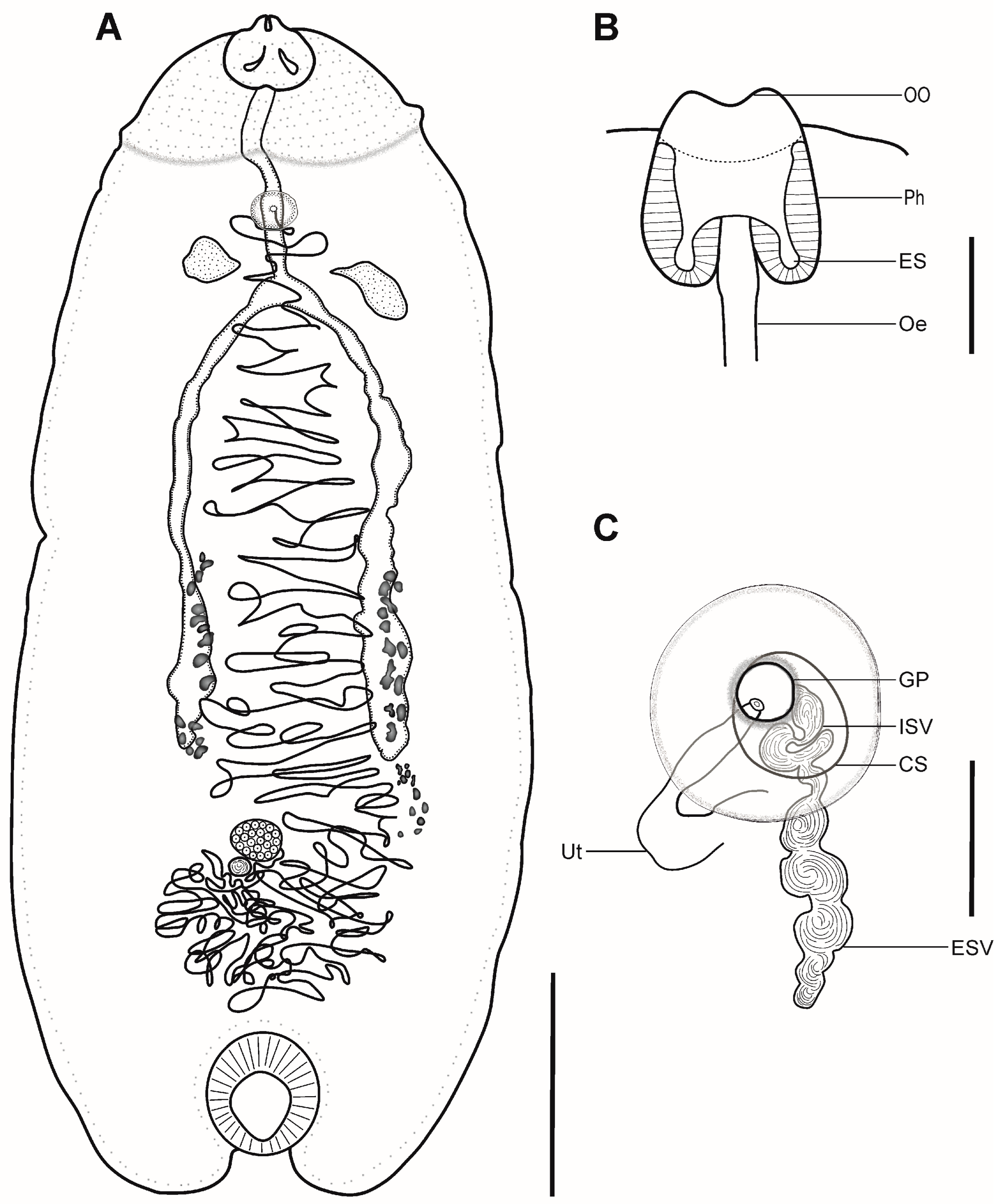
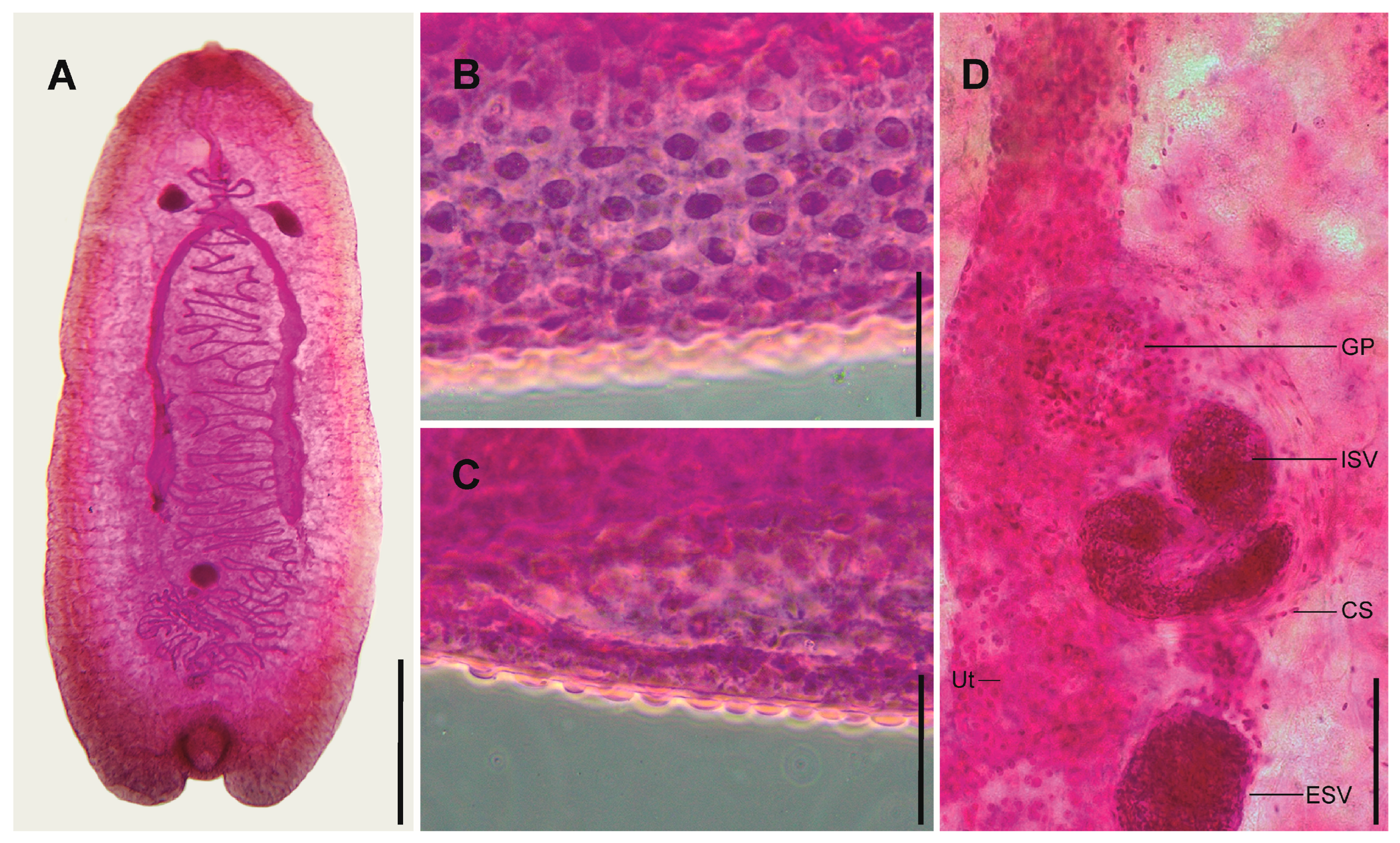
3.1. Kalitrema Kalitrema Travassos, 1933 (Figure 1 and Figure 2)

Remarks

3.2. Molecular Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reis, R.E.; Alberts, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish biodiversity and conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.R.; Hoberg, E.P. Triage for the biosphere: The need and rationale for taxonomic inventories and phylogenetic studies of parasites. Comp. Parasitol. 2000, 67, 1–25. [Google Scholar]
- Marcogliese, D.J. Parasites of the superorganism: Are they indicators of ecosystem health? Int. J. Parasitol. 2005, 35, 705–716. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Hopkins, S.; Bell, K.C.; Doña, J.; Godfrey, S.S.; Kwak, M.L.; Lafferty, K.D.; Moir, M.L.; Speer, K.A.; Strona, G.; et al. A global parasite conservation plan. Biol. Conserv. 2020, 250, 108596. [Google Scholar] [CrossRef]
- Sey, O. CRC Handbook of the Zoology of Amphistomes, 1st ed.; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Jones, A. Family Cladorchiidae Fischoeder, 1901. In Keys to the Trematoda; Jones, A., Bray, R.A., Gibson, D.I., Eds.; CAB International & The Natural History Museum: Wallingford, UK, 2005; Volume 2, pp. 257–317. [Google Scholar]
- Tonella, L.H.; Ruaro, R.; Daga, V.S.; Garcia, D.A.Z.; Vitorino, O.B.; Magalhães, T.L.; Reis, R.E.; Di Dario, F.; Petry, A.C.; Mincarone, M.M.; et al. Neotropical freshwater fishes: A dataset of occurrence and abundance of freshwater fishes in the Neotropics. Ecology 2023, 104, e3713. [Google Scholar] [CrossRef]
- Pantoja, C.; Scholz, T.; Luque, J.L.; Jones, A. New genera and species of paramphistomes (Digenea: Paramphistomoidea: Cladorchiidae) parasitic in fishes from the Amazon basin in Peru. Syst. Parasitol. 2018, 95, 611–624. [Google Scholar] [CrossRef]
- Pantoja, C.; Scholz, T.; Luque, J.L.; Jones, A. First molecular assessment of the interrelationships of cladorchiid digeneans (Digenea: Paramphistomoidea), parasites of Neotropical fishes, including descriptions of three new species and new host and geographical records. Folia Parasitol. 2019, 66, 011. [Google Scholar] [CrossRef]
- Choudhury, A.; Aguirre-Macedo, M.L.; Curran, S.S.; Ostrowski de Núñez, M.; Overstreet, R.M.; Pérez-Ponce de León, G.; Santos, C.P. Trematode diversity in freshwater fishes of the globe II: ‘New World’. Syst. Parasitol. 2016, 93, 271–282. [Google Scholar] [CrossRef]
- Travassos, L. Sur un nouveau trematode de poisons de la vallée du fleuve Parahyba. CR Soc. Biol. Paris 1933, 114, 839–840. [Google Scholar]
- Tkach, V.V.; Littlewood, D.T.J.; Olson, P.D.; Kinsella, J.M.; Swiderski, Z. Molecular phylogenetic analysis of the Microphalloidea Ward, 1901 (Trematoda: Digenea). Syst. Parasitol. 2003, 56, 1–15. [Google Scholar] [CrossRef]
- Luton, K.; Walker, D.; Blair, D. Comparisons of ribosomal internal transcribed spacers from two congeneric species of flukes (Platyhelminthes: Trematoda: Digenea). Mol. Biochem. Parasitol. 1992, 56, 323–327. [Google Scholar] [CrossRef]
- Miura, O.; Kuris, A.M.; Torchin, M.E.; Hechinger, R.F.; Dunham, E.J.; Chiba, S. Molecular-genetic analyses reveal cryptic species of trematodes in the intertidal gastropod, Batillaria cumingi (Crosse). Int. J. Parasitol. 2005, 35, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.V.; Assis, J.C.A.; López-Hernández, D.; Pulido-Murillo, E.A.; Melo, A.L.; Locke, S.A.; Pinto, H.A. A phylogenetic study of the cecal amphistome Zygocotyle lunata (Trematoda: Zygocotylidae), with notes on the molecular systematics of Paramphistomoidea. Parasitol. Res. 2020, 119, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Benovics, M.; Mikulíček, P.; Žákovicová, Z.; Papežík, P.; Pantoja, C. Hidden in plain sight: Novel molecular data reveal unexpected genetic diversity among paramphistome parasites (Digenea: Paramphistomoidea) of European water frogs. Parasitology 2022, 149, 1425–1438. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFTT multiple sequence alignment software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res. 2015, 1, W7–W14. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- ICZN International Code of Zoological Nomenclature, 4th ed.; 1999; Available online: http://www.iczn.org (accessed on 15 March 2023).
- Huson, K.M.; Oliver, N.A.; Robinson, M.W. Paramphistomosis of ruminants: An emerging parasitic disease in Europe. Trends Parasitol. 2017, 33, 836–844. [Google Scholar] [CrossRef]
- Tandon, V.; Roy, B.; Shylla, J.A.; Ghatani, S. Amphistomes. In Digenetic Trematodes; Toledo, R., Fried, B., Eds.; Springer: Cham, Switzerland, 2019; Volume 1154, pp. 255–277. [Google Scholar]
- Virgilio, L.R.; Martins, W.M.O.; Lima, F.S.; Takemoto, R.M.; Camargo, L.M.A.; Meneguetti, D.U.O. Endoparasite fauna of freshwater fish from the upper Juruá River in the Western Amazon, Brazil. J. Helminthol. 2022, 96, e55. [Google Scholar] [CrossRef]
- Cherem, L.F.S.; Varajão, C.A.C.; Braucher, R.; Bourlés, D.; Salgado, A.A.R.; Varajão, A.C. Long-term evolution of denudational escarpments in southeastern Brazil. Geomorphology 2012, 173, 118–127. [Google Scholar] [CrossRef]
- Anjos, M.S.; Queiroz, L.J.; Penido, I.S.; Bitencourt, J.A.; Barreto, S.B.; Sarmento-Soares, L.M.; Batalha-Filho, H.; Affonso, P.R.A.M. A taxonomically complex catfish group from an underrepresented geographic area: Systematics and species limits in Hypostomus Lacépède, 1803 (Siluriformes, Loricariidae) from Eastern South America. J. Zool. Syst. Evol. Res. 2021, 59, 1994–2009. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Present Study | Travassos (1933) | |
| Host | Hypostomus alatus | Hypostomus punctatus | |
| Hypostomus francisci | |||
| N | N = 6 | N = 2 | |
| Body | L | 7729 (6175–9900) | 7000, 8700 |
| W | 2979 (2100–4400) | 2600, 3000 | |
| Body length/body width | 1:2.59 (1:2.25–3.34) | – | |
| Pharynx | L | 421 (260–595) | – |
| W | 493 (355–691) | – | |
| Oesophagus | L | 1314 (1035–1759) | 780,1300 |
| DTRAE | 1117 (821–1386) | 800, 900 | |
| Acetabulum | L | 823 (601–1095) | 760 |
| W | 797 (631–969) | 780 | |
| DIBAE | 1810 (1453–2271) | – | |
| Right testis | L | 300 (190–417) | 570 |
| W | 315 (211–485) | 390, 440 | |
| Left testis | L | 331 (216–382) | 570 |
| W | 346 (140–797) | 390, 440 | |
| External seminal vesicle | L | 378 (243–478) | – |
| W | 61 (45–93) | – | |
| Cirrus-sac | L | 252 (110–348) | – |
| W | 191 (105–367) | – | |
| Internal seminal vesicle | L | 304 (206–382) | – |
| W | 73 (50–95) | – | |
| GRSGP | L | 363 (276–553) | 310, 390 |
| W | 369 (230–558) | 310, 390 | |
| Genital pore opening | L | 81 (62–140) | – |
| W | 100 (61–209) | – | |
| Ovary | L | 299 (221–373) | 350 |
| W | 354 (281–450) | 360 | |
| Seminal receptacle | L | 165 (122–208) | – |
| W | 175 (148–211) | – | |
| DBOT | 3400 (2537–4150) | – | |
| Post-ovarian region | % | 31 (23–34) | – |
| Egg | L | n/a | 160, 168 |
| W | n/a | 96, 112 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, H.A.; Pantoja, C.; Assis, J.C.A.d.; López-Hernández, D.; Vieira, F.; Luque, J.L.; Alves, P.V. A Rare Fish Amphistome Revisited: The Phylogenetic Position of Kalitrema kalitrema (Trematoda: Cladorchiidae) Found in Hypostomus spp. (Siluriformes: Loricariidae) from Brazil. Diversity 2023, 15, 1034. https://doi.org/10.3390/d15101034
Pinto HA, Pantoja C, Assis JCAd, López-Hernández D, Vieira F, Luque JL, Alves PV. A Rare Fish Amphistome Revisited: The Phylogenetic Position of Kalitrema kalitrema (Trematoda: Cladorchiidae) Found in Hypostomus spp. (Siluriformes: Loricariidae) from Brazil. Diversity. 2023; 15(10):1034. https://doi.org/10.3390/d15101034
Chicago/Turabian StylePinto, Hudson Alves, Camila Pantoja, Jordana Costa Alves de Assis, Danimar López-Hernández, Fabio Vieira, José Luis Luque, and Philippe Vieira Alves. 2023. "A Rare Fish Amphistome Revisited: The Phylogenetic Position of Kalitrema kalitrema (Trematoda: Cladorchiidae) Found in Hypostomus spp. (Siluriformes: Loricariidae) from Brazil" Diversity 15, no. 10: 1034. https://doi.org/10.3390/d15101034
APA StylePinto, H. A., Pantoja, C., Assis, J. C. A. d., López-Hernández, D., Vieira, F., Luque, J. L., & Alves, P. V. (2023). A Rare Fish Amphistome Revisited: The Phylogenetic Position of Kalitrema kalitrema (Trematoda: Cladorchiidae) Found in Hypostomus spp. (Siluriformes: Loricariidae) from Brazil. Diversity, 15(10), 1034. https://doi.org/10.3390/d15101034