

Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean)



Abstract
1. Introduction
2. Materials and Methods
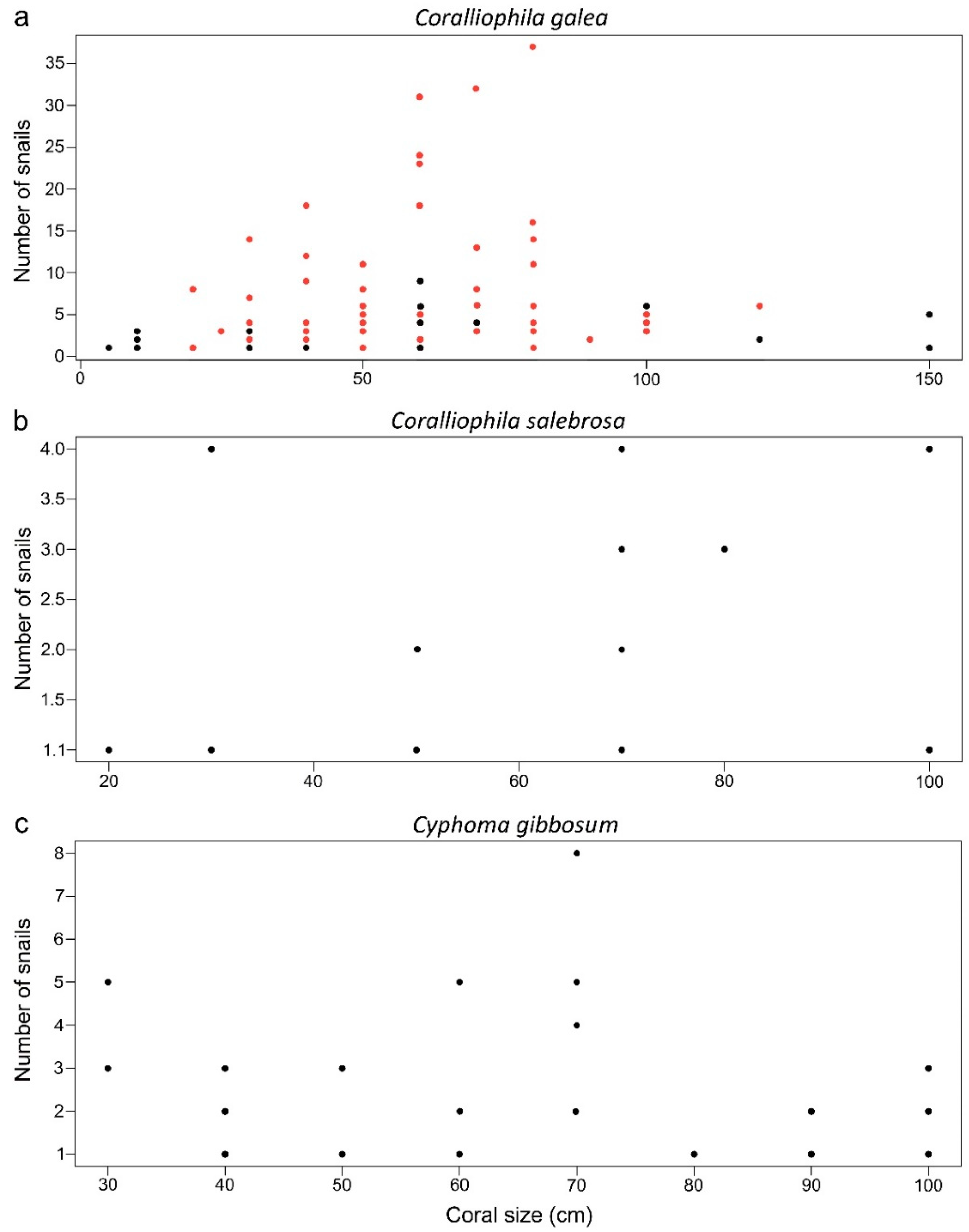
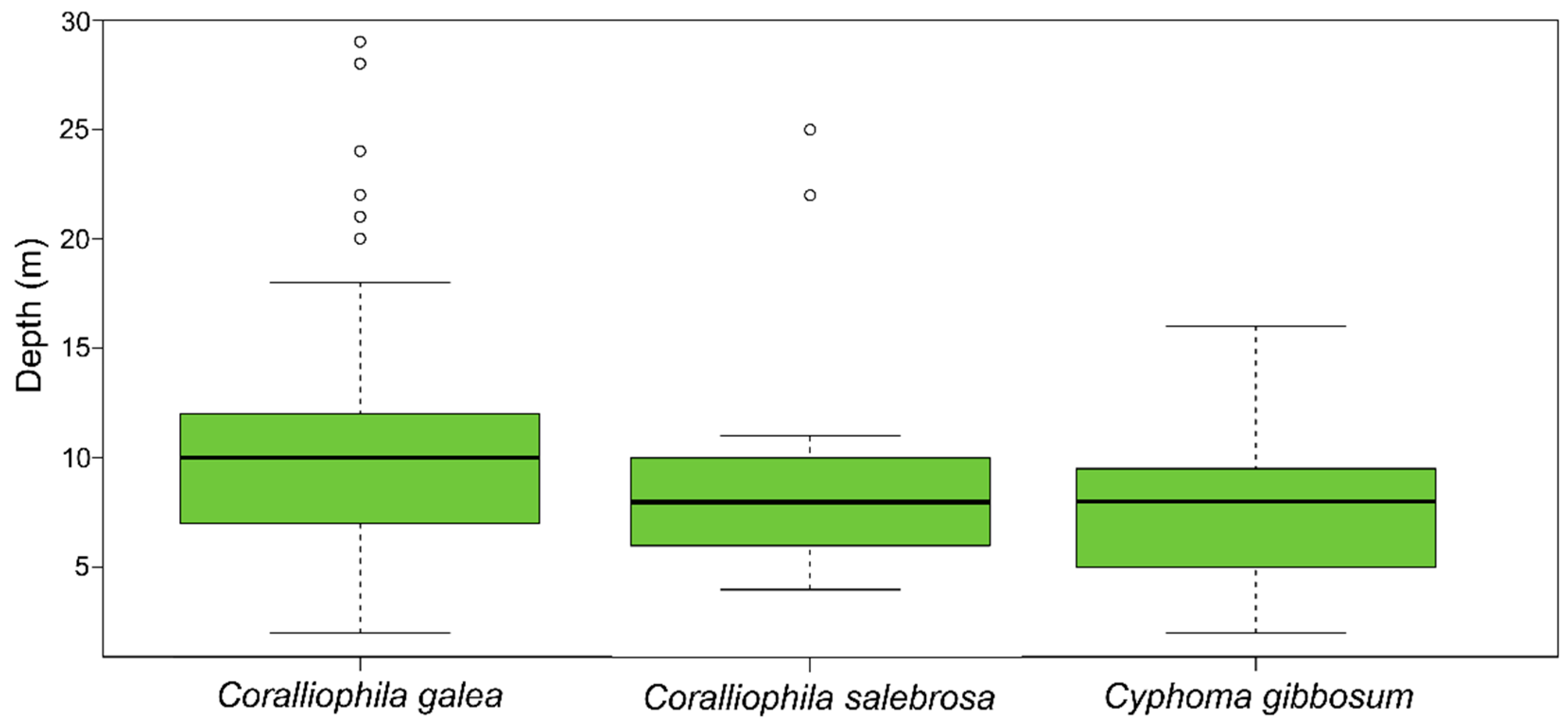
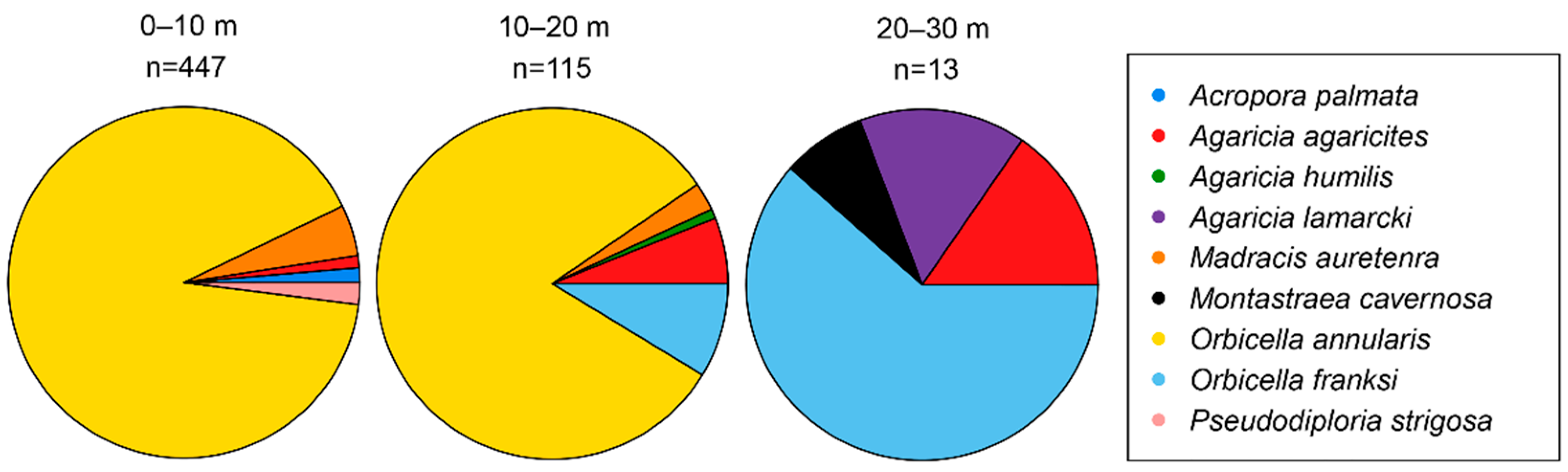
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reaka-Kudla, M.L. Biodiversity of Caribbean coral reefs. In Caribbean Marine Biodiversity: The Known and the Unknown; Miloslavich, P., Klein, E., Eds.; DEStech Publishers: Lancaster, PA, USA, 2005; pp. 259–276. [Google Scholar]
- Miloslavich, P.; Díaz, J.M.; Klein, E.; Alvarado, J.J.; Díaz, C.; Gobin, J.; Escobar-Briones, E.; Cruz-Motta, J.J.; Weil, E.; Cortés, J.; et al. Marine biodiversity in the Caribbean: Regional estimates and distribution patterns. PLoS ONE 2010, 5, e11916. [Google Scholar] [CrossRef] [PubMed]
- DeVantier, L.; Turak, E.; Szava-Kovats, R. Species richness and abundance of reef-building corals in the Indo-West Pacific: The local–regional relation revisited. Front. Mar. Sci. 2020, 7, 487. [Google Scholar] [CrossRef]
- Stella, J.S.; Pratchett, M.S.; Hutchings, P.A.; Jones, G.P. Coral-associated invertebrates: Diversity, ecological importance and vulnerability to disturbance. Oceanogr. Mar. Biol. Annu. Rev. 2011, 49, 43–104. [Google Scholar]
- Hoeksema, B.W.; van der Meij, S.E.T.; Fransen, C.H.J.M. The mushroom coral as a habitat. J. Mar. Biol. Assoc. U. K. 2012, 92, 647–663. [Google Scholar] [CrossRef]
- Nützel, A. Gastropods as parasites and carnivorous grazers: A major guild in marine ecosystems. In The Evolution and Fossil Record of Parasitism; De Baets, K., Huntley, J.W., Eds.; Springer: Cham, Switzerland, 2021; pp. 209–229. [Google Scholar] [CrossRef]
- Lucas, M.Q.; Rodríguez, L.R.; Sanabria, D.J.; Weil, E. Natural prey preferences and spatial variability of predation pressure by Cyphoma gibbosum (Mollusca: Gastropoda) on octocoral communities off La Parguera, Puerto Rico. Int. Sch. Res. Notices 2014, 2014, 742387. [Google Scholar] [CrossRef]
- Pinto, T.K.; Benevides, L.J.; Sampaio, C.L.S. Cyphoma macumba Petuch 1979 (Gastropoda: Ovulidae): A versatile predator of the Brazilian Octocorallia. Mar. Biodivers. 2017, 47, 165–166. [Google Scholar] [CrossRef]
- Shaver, E.C.; Renzi, J.J.; Bucher, M.G.; Silliman, B.R. Relationships between a common Caribbean corallivorous snail and protected area status, coral cover, and predator abundance. Sci. Rep. 2020, 10, 16463. [Google Scholar] [CrossRef]
- Gittenberger, A.; Hoeksema, B.W. Habitat preferences of coral-associated wentletrap snails (Gastropoda: Epitoniidae). Contrib. Zool. 2013, 82, 1–25. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Scott, C.; True, J.D. Dietary shift in corallivorous Drupella snails following a major bleaching event at Koh Tao, Gulf of Thailand. Coral Reefs 2013, 32, 423–428. [Google Scholar] [CrossRef]
- Potkamp, G.; Vermeij, M.J.A.; Hoeksema, B.W. Genetic and morphological variation in corallivorous snails (Coralliophila spp.) living on different host corals at Curaçao, southern Caribbean. Contrib. Zool. 2017, 86, 111–144. [Google Scholar] [CrossRef]
- Kaullysing, D.; Taleb-Hossenkhan, N.; Kulkarni, B.; Bhagooli, R. Variations in the density of two ectoparasitic gastropods (Coralliophila spp.) on scleractinian corals on a coast-reef scale. Symbiosis 2019, 78, 65–71. [Google Scholar] [CrossRef]
- Moerland, M.S.; Scott, C.M.; Hoeksema, B.W. Prey selection of corallivorous muricids at Koh Tao (Gulf of Thailand) four years after a major coral bleaching event. Contrib. Zool. 2016, 85, 291–309. [Google Scholar] [CrossRef]
- Saponari, L.; Dehnert, I.; Galli, P.; Montano, S. Assessing population collapse of Drupella spp. (Mollusca: Gastropoda) 2 years after a coral bleaching event in the Republic of Maldives. Hydrobiologia 2021, 848, 2653–2666. [Google Scholar] [CrossRef]
- Sánchez, J.A. Remarkable specialization in Eastern Pacific sea fan ectoparasites (Neosimnia). Coral Reefs 2013, 32, 891. [Google Scholar] [CrossRef][Green Version]
- Sánchez, J.A.; Fuentes-Pardo, A.P.; Almhain, Í.N.; Ardila-Espitia, N.E.; Cantera-Kintz, J.; Forero-Shelton, M. The masquerade game: Marine mimicry adaptation between egg-cowries and octocorals. PeerJ 2016, 4, e2051. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger, A.; Gittenberger, E. A hitherto unnoticed adaptive radiation: Epitoniid species (Gastropoda: Epitoniidae) associated with corals (Scleractinia). Contrib. Zool. 2005, 74, 125–203. [Google Scholar] [CrossRef]
- Oliverio, M. Coralliophilinae (Neogastropoda: Muricidae) from the southwest Pacific. Mem. Mus. Natl. Hist. Nat. 2008, 196, 481–585. [Google Scholar]
- Taviani, M.; Angeletti, L.; Dimech, M.; Mifsud, C.; Freiwald, A.; Harasewych, M.G.; Oliverio, M. Coralliophilinae (Gastropoda: Muricidae) associated with deep-water coral banks in the Mediterranean. Nautilus 2009, 123, 106–112. [Google Scholar]
- Claremont, M.; Houart, R.; Williams, S.T.; Reid, D.G. A molecular phylogenetic framework for the Ergalataxinae (Neogastropoda: Muricidae). J. Mollusc. Stud. 2013, 79, 19–29. [Google Scholar] [CrossRef][Green Version]
- Goud, J.; Hoeksema, B.W. Pedicularia vanderlandi spec. nov., a symbiotic snail (Caenogastropoda: Ovulidae) on the hydrocoral Distichopora vervoorti Cairns and Hoeksema, 1998 (Hydrozoa: Stylasteridae), from Bali, Indonesia. Zool. Verh. 2001, 334, 77–97. [Google Scholar]
- Lorenz, F.; Fehse, D. The Living Ovulidae. A Manual of the Families of Allied Cowries: Ovulidae, Pediculariidae and Eocypraeidae; Conchbooks: Hackenheim, Germany, 2009. [Google Scholar]
- Braga-Henriques, A.; Carreiro-Silva, M.; Porteiro, F.M.; de Matos, V.; Sampaio, Í.; Ocaña, O.; Avila, S.P. The association between a deep-sea gastropod Pedicularia sicula (Caenogastropoda: Pediculariidae) and its coral host Errina dabneyi (Hydrozoa: Stylasteridae) in the Azores. ICES J. Mar. Sci. 2011, 68, 399–407. [Google Scholar] [CrossRef]
- Schiaparelli, S.; Barucca, M.; Olmo, E.; Boyer, M.; Canapa, A. Phylogenetic relationships within Ovulidae (Gastropoda: Cypraeoidea) based on molecular data from the 16S rRNA gene. Mar. Biol. 2005, 147, 411–420. [Google Scholar] [CrossRef]
- Reijnen, B.T.; Hoeksema, B.W.; Gittenberger, E. Host specificity and phylogenetic relationships among Atlantic Ovulidae (Mollusca: Gastropoda). Contrib. Zool. 2010, 79, 69–78. [Google Scholar] [CrossRef]
- Fritts-Penniman, A.L.; Gosliner, T.M.; Mahardika, G.N.; Barber, P.H. Cryptic ecological and geographic diversification in coral-associated nudibranchs. Mol. Phylogenet. Evol. 2020, 144, 106698. [Google Scholar] [CrossRef]
- Mehrotra, R.; Arnold, S.; Wang, A.; Chavanich, S.; Hoeksema, B.W.; Caballer, M. A new species of coral-feeding nudibranch (Mollusca: Gastropoda) from the Gulf of Thailand. Mar. Biodivers. 2020, 50, 36. [Google Scholar] [CrossRef]
- Wang, A.; Conti-Jerpe, I.E.; Richards, J.L.; Baker, D.M. Phestilla subodiosus sp. nov. (Nudibranchia, Trinchesiidae), a corallivorous pest species in the aquarium trade. ZooKeys 2020, 909, 35278. [Google Scholar] [CrossRef]
- Yiu, S.K.F.; Chung, S.S.W.; Qiu, J.W. New observations on the corallivorous nudibranch Phestilla melanobrachia: Morphology, dietary spectrum and early development. J. Mollusc. Stud. 2021, 87, eyab034. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Gittenberger, A. Records of some marine parasitic molluscs from Nha Trang, Vietnam. Basteria 2008, 72, 129–133. [Google Scholar]
- Gittenberger, A.; Gittenberger, E. Cryptic, adaptive radiation of endoparasitic snails: Sibling species of Leptoconchus (Gastropoda: Coralliophilidae) in corals. Org. Divers. Evol. 2011, 11, 21–41. [Google Scholar] [CrossRef]
- Bieler, R.; Petit, R.E. Catalogue of Recent and fossil “worm-snail” taxa of the families Vermetidae, Siliquariidae, and Turritellidae (Mollusca: Caenogastropoda). Zootaxa 2011, 2948, 1–103. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Harper, C.E.; Langdon-Down, S.J.; van der Schoot, R.J.; Smith-Moorhouse, A.; Spaargaren, R.; Timmerman, R.F. Host range of the coral-associated worm snail Petaloconchus sp. (Gastropoda: Vermetidae), a newly discovered cryptogenic pest species in the southern Caribbean. Diversity 2022, 14, 196. [Google Scholar] [CrossRef]
- Knowlton, N.; Lang, J.C.; Keller, B.D. Case study of natural population collapse: Post-hurricane predation on Jamaican staghorn corals. Smithson. Contrib. Mar. Sci. 1990, 31, 1–25. [Google Scholar] [CrossRef]
- Bruckner, A.W.; Bruckner, R.J. Mechanical lesions and corallivory. In Diseases of Coral; Woodley, C.M., Downs, C.A., Bruckner, A.W., Porter, J.W., Galloway, S.B., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 242–265. [Google Scholar] [CrossRef]
- Scott, C.M.; Mehrotra, R.; Hein, M.Y.; Moerland, M.S.; Hoeksema, B.W. Population dynamics of corallivores (Drupella and Acanthaster) on coral reefs of Koh Tao, a diving destination in the Gulf of Thailand. Raffles Bull. Zool. 2017, 65, 68–79. [Google Scholar]
- Boucher, L.M. Coral predation by muricid gastropods of the genus Drupella at Enewetak, Marshall Islands. Bull. Mar. Sci. 1986, 38, 9–11. [Google Scholar]
- Cumming, R.; McCorry, D. Corallivorous gastropods in Hong Kong. Coral Reefs 1998, 17, 178. [Google Scholar] [CrossRef]
- Cumming, R.L. Predation on reef-building corals: Multiscale variation in the density of three corallivorous gastropods, Drupella spp. Coral Reefs 1999, 18, 147–157. [Google Scholar] [CrossRef]
- Schärer, M.T.; Nemeth, M.I. Mass mortality of gorgonians due to a Cyphoma gibbosum (Linnaeus) population outbreak at Mona Island, Puerto Rico. Coral Reefs 2010, 29, 533. [Google Scholar] [CrossRef]
- Bessey, C.; Babcock, R.C.; Thomson, D.P.; Haywood, M.D.E. Outbreak densities of the coral predator Drupella in relation to in situ Acropora growth rates on Ningaloo Reef, Western Australia. Coral Reefs 2018, 37, 985–993. [Google Scholar] [CrossRef]
- Bruckner, A.W.; Coward, G.; Bimson, K.; Rattanawongwan, T. Predation by feeding aggregations of Drupella spp. inhibits the recovery of reefs damaged by a mass bleaching event. Coral Reefs 2017, 36, 1181–1187. [Google Scholar] [CrossRef]
- Hamman, E.A. Aggregation patterns of two corallivorous snails and consequences for coral dynamics. Coral Reefs 2018, 37, 851–860. [Google Scholar] [CrossRef]
- Dalton, S.J.; Godwin, S. Progressive coral tissue mortality following predation by a corallivorous nudibranch (Phestilla sp.). Coral Reefs 2006, 25, 529. [Google Scholar] [CrossRef]
- Gignoux-Wolfsohn, S.; Marks, C.; Vollmer, S. White Band Disease transmission in the threatened coral, Acropora cervicornis. Sci. Rep. 2012, 2, 804. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, L.J.; Work, T.M.; Miller, R.L.; Lozada-Misa, P.L. Effects of Coralliophila violacea on tissue loss in the scleractinian corals Porites spp. depend on host response. Dis. Aquat. Organ. 2016, 119, 75–83. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nicolet, K.J.; Chong-Seng, K.M.; Pratchett, M.S.; Willis, B.L.; Hoogenboom, M.O. Predation scars may influence host susceptibility to pathogens: Evaluating the role of corallivores as vectors of coral disease. Sci. Rep. 2018, 8, 5258. [Google Scholar] [CrossRef] [PubMed]
- Montano, S.; Aeby, G.; Galli, P.; Hoeksema, B.W. Feeding behavior of Coralliophila sp. on corals affected by Caribbean ciliate infection (CCI): A new possible vector? Diversity 2022, 14, 363. [Google Scholar] [CrossRef]
- Thur, S.M. User fees as sustainable financing mechanisms for marine protected areas: An application to the Bonaire National Marine Park. Mar. Policy 2010, 34, 63–69. [Google Scholar] [CrossRef]
- Trembanis, A.C.; Forrest, A.L.; Keller, B.M.; Patterson, M.R. Mesophotic coral ecosystems: A geoacoustically derived proxy for habitat and relative diversity for the leeward shelf of Bonaire, Dutch Caribbean. Front. Mar. Sci. 2017, 4, 51. [Google Scholar] [CrossRef]
- Bak, R.P.M.; Nieuwland, G.; Meesters, E.H. Coral reef crisis in deep and shallow reefs: 30 years of constancy and change in reefs of Curaçao and Bonaire. Coral Reefs 2005, 24, 475–479. [Google Scholar] [CrossRef]
- Stokes, M.D.; Leichter, J.J.; Genovese, S.J. Long-term declines in coral cover at Bonaire, Netherlands Antilles. Atoll Res. Bull. 2010, 582, 1–21. [Google Scholar] [CrossRef]
- de Bakker, D.M.; Meesters, E.H.; Bak, R.P.M.; Nieuwland, G.; van Duyl, F.C. Long-term shifts in coral communities on shallow to deep reef slopes of Curaçao and Bonaire: Are there any winners? Front. Mar. Sci. 2016, 3, 247. [Google Scholar] [CrossRef]
- de Bakker, D.M.; van Duyl, F.C.; Bak, R.P.M.; Nugues, M.M.; Nieuwland, G.; Meesters, E.H. 40 years of benthic community change on the Caribbean reefs of Curaçao and Bonaire: The rise of slimy cyanobacterial mats. Coral Reefs 2017, 36, 355–367. [Google Scholar] [CrossRef]
- Hoeksema, B.W. (Ed.) Marine Biodiversity of Bonaire: A Baseline Survey, 2nd ed.; Naturalis Biodiversity Center: Leiden, The Netherlands; ANEMOON Foundation: Bennebroek, The Netherlands, 2022. [Google Scholar]
- García-Hernández, J.E.; de Gier, W.; van Moorsel, G.W.N.M.; Hoeksema, B.W. The scleractinian Agaricia undata as a new host for the Caribbean coral gall crab Opecarcinus hypostegus at Bonaire, southern Caribbean. Symbiosis 2020, 81, 303–311. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; García-Hernández, J.E. Host-related morphological variation of dwellings inhabited by the crab Domecia acanthophora in the corals Acropora palmata and Millepora complanata (Southern Caribbean). Diversity 2020, 12, 143. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Olthof, G.; ten Hove, H.A. Extension of the recorded host range of Caribbean Christmas tree worms (Spirobranchus spp.) with two scleractinians, a zoantharian, and an ascidian. Diversity 2020, 12, 115. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Timmerman, R.F.; Spaargaren, R.; Smith-Moorhouse, A.; van der Schoot, R.J.; Langdon-Down, S.J.; Harper, C.E. Morphological modifications and injuries of corals caused by symbiotic feather duster worms (Sabellidae) in the Caribbean. Diversity 2022, 14, 332. [Google Scholar] [CrossRef]
- Korzhavina, O.A.; Hoeksema, B.W.; Ivanenko, V.N. A review of Caribbean Copepoda associated with reef-dwelling cnidarians, echinoderms and sponges. Contrib. Zool. 2019, 88, 297–349. [Google Scholar] [CrossRef]
- Montano, S.; Reimer, J.D.; Ivanenko, V.N.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Galli, P.; Hoeksema, B.W. Widespread occurrence of a rarely known association between the hydrocorals Stylaster roseus and Millepora alcicornis at Bonaire, southern Caribbean. Diversity 2020, 12, 218. [Google Scholar] [CrossRef]
- Montenegro, J.; Hoeksema, B.W.; Santos, M.E.A.; Kise, H.; Reimer, J.D. Zoantharia (Cnidaria: Hexacorallia) of the Dutch Caribbean with historical distribution records from the Atlantic and one new species of Parazoanthus. Diversity 2020, 12, 190. [Google Scholar] [CrossRef]
- Reimer, J.D.; Wee, H.B.; García-Hernández, J.E.; Hoeksema, B.W. Same but different? Zoantharian assemblages (Anthozoa: Hexacorallia: Zoantharia) in Bonaire and Curaçao, southern Caribbean. Coral Reefs 2022, 41, 383–396. [Google Scholar] [CrossRef]
- Rotjan, R.D.; Lewis, S.M. Impact of coral predators on tropical reefs. Mar. Ecol. Prog. Ser. 2008, 367, 73–91. [Google Scholar] [CrossRef]
- Miller, M. Corallivorous snail removal: Evaluation of impact on Acropora palmata. Coral Reefs 2001, 19, 293–295. [Google Scholar] [CrossRef]
- Precht, W.; Bruckner, A.; Aronson, R.; Bruckner, R. Endangered acroporid corals of the Caribbean. Coral Reefs 2002, 21, 41–42. [Google Scholar] [CrossRef]
- Garrigues, B.; Lamy, D.; Zuccon, D. The Coralliophilinae from the Antilles and French Guiana with the description of six new species. Xenophora Taxon. 2022, 37, 4–53. [Google Scholar]
- Johnston, L.; Miller, M.W. Variation in life-history traits of the corallivorous gastropod Coralliophila abbreviata on three coral hosts. Mar. Biol. 2007, 150, 1215–1225. [Google Scholar] [CrossRef]
- Johnston, L.; Miller, M.W.; Baums, I.B. Assessment of host-associated genetic differentiation among phenotypically divergent populations of a coral-eating gastropod across the Caribbean. PLoS ONE 2012, 7, e47630. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, S.E.; Chou, V.; Cheng, S.H.; Rachmawati, R.; Calumpong, H.P.; Ngurah Mahardika, G.; Barber, P.H. Evidence of host-associated divergence from coral-eating snails (genus Coralliophila) in the Coral Triangle. Coral Reefs 2018, 37, 355–371. [Google Scholar] [CrossRef]
- Miller, A.C. Observations on the associations and feeding of six species of prosobranch gastropods on anthozoans. Atoll Res. Bull. 1972, 152, 4–5. [Google Scholar]
- Martin, D.; Gil, J.; Abgarian, C.; Evans, E.; Turner, E.M.; Nygren, A. Coralliophila from Grand Cayman: Specialized coral predator or parasite? Coral Reefs 2014, 33, 1017. [Google Scholar] [CrossRef][Green Version]
- Ott, B.; Lewis, J.B. The importance of the gastropod Coralliophila abbreviata (Lamarck) and the polychaete Hermodice carunculata (Pallas) as coral reef predators. Can. J. Zool. 1972, 50, 1651–1656. [Google Scholar] [CrossRef]
- Reijnen, B.T.; van der Meij, S.E.T. Coat of many colours-DNA reveals polymorphism of mantle patterns and colouration in Caribbean Cyphoma Röding, 1798 (Gastropoda, Ovulidae). PeerJ 2017, 5, e3018. [Google Scholar] [CrossRef]
- Lorenz, F. The “Black Morph Cyphoma” from the Netherlands Antilles (Gastropoda: Ovulidae). Acta Conchyl. 2020, 19, 69–75. [Google Scholar]
- Nowlis, J.P. Mate-and oviposition-influenced host preferences in the coral-feeding snail Cyphoma gibbosum. Ecology 1993, 74, 1959–1969. [Google Scholar] [CrossRef]
- Chiappone, M.; Dienes, H.; Swanson, D.W.; Miller, S.L. Density and Gorgonian host-occupation patterns by Flamingo Tongue snails (Cyphoma gibbosum) in the Florida keys. Caribb. J. Sci. 2003, 39, 116–127. [Google Scholar]
- Lasker, H.R.; Coffroth, M.A.; Fitzgerald, L.M. Foraging patterns of Cyphoma gibbosum on octocorals: The roles of host choice and feeding preference. Biol. Bull. 1988, 174, 254–266. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Koh, E.G.L. Depauperation of the mushroom coral fauna (Fungiidae) of Singapore (1860s–2006) in changing reef conditions. Raffles Bull. Zool. Suppl. 2009, 22, 91–101. [Google Scholar]
- Ruesink, J.L.; Harvell, C.D. Specialist predation on the Caribbean gorgonian Plexaurella spp. by Cyphoma signatum (Gastropoda). Mar. Ecol. Prog. Ser. 1990, 65, 265–272. [Google Scholar] [CrossRef]
- Humann, P.; Deloach, N. Reef Coral Identification: Florida, the Caribbean and the Bahamas, 3rd ed.; New World Publications Inc.: Jacksonville, FL, USA, 2013. [Google Scholar]
- Hoeksema, B.W.; van der Loos, L.M.; van Moorsel, G.W.N.M. Coral diversity matches marine park zonation but not economic value of coral reef sites at St. Eustatius, eastern Caribbean. J. Environ. Manag. 2022, 320, 115829. [Google Scholar] [CrossRef]
- Bayer, F.M. The shallow-water Octocorallia of the West Indian region. Stud. Fauna Curaçao Caribb. Isl. 1961, 12, 506065. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 11 September 2022).
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite Package: Analysing Ecological Networks. R News 2008, 8, 8–11. [Google Scholar]
- Dormann, C.F.; Fruend, J.; Gruber, B.; Beckett, S.; Devoto, M.; Felix, G.M.F.; Iriondo, J.M.; Opsahl, T.; Pinheiro, R.B.P.; Strauss, R.; et al. Visualising Bipartite Networks and Calculating Some (Ecological) Indices. 2022. Available online: https://github.com/biometry/bipartite (accessed on 11 September 2022).
- Pavlopoulos, G.A.; Kontou, P.I.; Pavlopoulou, A.; Bouyioukos, C.; Markou, E.; Bagos, P.G. Bipartite graphs in systems biology and medicine: A survey of methods and applications. GigaScience 2018, 7, giy014. [Google Scholar] [CrossRef]
- Coker, D.J.; DiBattista, J.D.; Stat, M.; Arrigoni, R.; Reimer, J.; Terraneo, T.I.; Villabos, R.; Nowicki, J.P.; Bunce, M.; Berumen, M.L. DNA metabarcoding confirms primary targets and breadth of diet for coral reef butterflyfishes. Coral Reefs 2022, in press. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef]
- Brawley, S.H.; Adey, W.H. Coralliophila abbreviata: A significant corallivore! Bull. Mar. Sci. 1982, 32, 595–599. [Google Scholar]
- Frade, P.R.; Bongaerts, P.; Baldwin, C.C.; Trembanis, A.C.; Bak, R.P.M.; Vermeij, M.J.A. Bonaire and Curaçao. In Mesophotic Coral Ecosystems. Coral Reefs of the World; Loya, Y., Puglise, K., Bridge, T., Eds.; Springer: Cham, Switzerland, 2019; Volume 12, pp. 149–162. [Google Scholar] [CrossRef]
- Bongaerts, P.; Ridgway, T.; Sampayo, E.M.; Hoegh-Guldberg, O. Assessing the ‘deep reef refugia’ hypothesis: Focus on Caribbean reefs. Coral Reefs 2010, 29, 309–327. [Google Scholar] [CrossRef]
- Potkamp, G.; Vermeij, M.J.A.; Hoeksema, B.W. Host-dependent variation in density of corallivorous snails (Coralliophila spp.) at Curaçao, southern Caribbean. Mar. Biodivers. 2017, 47, 91–99. [Google Scholar] [CrossRef]
- Hayes, J.A. Distribution, movement and impact of the corallivorous gastropod Coralliophila abbreviata (Lamarck) on a Panamánian patch reef. J. Exp. Mar. Biol. Ecol. 1990, 142, 25–42. [Google Scholar] [CrossRef]
- Baums, I.B.; Miller, M.W.; Szmant, A.M. Ecology of a corallivorous gastropod, Coralliophila abbreviata, on two seleractinian hosts. II. Feeding, respiration and growth. Mar. Biol. 2003, 142, 1093–1101. [Google Scholar] [CrossRef]
- Van der Schoot, R.J.; Hoeksema, B.W. Abundance of coral-associated fauna in relation to depth and eutrophication along the leeward side of Curaçao, southern Caribbean. Mar. Environ. Res. 2022, 181, 105738. [Google Scholar] [CrossRef]
- Bruckner, A.; Bruckner, R. Condition of restored Acropora palmata fragments off Mona Island, Puerto Rico, 2 years after the Fortuna Reefer ship grounding. Coral Reefs 2001, 20, 235–243. [Google Scholar] [CrossRef]
- Williams, D.E.; Miller, M.W. Attributing mortality among drivers of population decline in Acropora palmata in the Florida Keys (USA). Coral Reefs 2012, 31, 369–382. [Google Scholar] [CrossRef]
- Bright, A.J.; Rogers, C.S.; Brandt, M.E.; Muller, E.; Smith, T.B. Disease prevalence and snail predation associated with swell-generated damage on the threatened coral, Acropora palmata (Lamarck). Front. Mar. Sci. 2016, 3, 77. [Google Scholar] [CrossRef]
- Queiroz, V.; Sales, L.; Neves, E.; Johnsson, R. Siderastrea stellata Verrill, 1868 as a new host for the corallivore snail Coralliophila caribaea Abbott, 1958 and some biological aspects of their association. Spixiana 2021, 44, 17–18. [Google Scholar]
- Del Mónaco, C.; Villamiza, E.; Narciso, S. Selectividad de presas de Coralliophila abbreviata y C. caribaea en arrecifes coralinos del Parque Nacional Morrocoy, Venezuela: Una aproximación experimental. Lat. Am. J. Aquat. Res. 2010, 38, 57–70. [Google Scholar] [CrossRef]
- Miller, A.C. Cnidarian prey of the snails Coralliophila abbreviata and C. caribaea (Gastropoda: Muricidae) in Discovery Bay, Jamaica. Bull. Mar. Sci. 1981, 31, 932–934. [Google Scholar]
- Oren, U.; Brickner, I.; Loya, Y. Prudent sessile feeding by the corallivore snail, Coralliophila violacea on coral energy sinks. Proc. R. Soc. B 1998, 265, 2043–2050. [Google Scholar] [CrossRef]
- Clements, C.S.; Hay, M.E. Overlooked coral predators suppress foundation species as reefs degrade. Ecol. Appl. 2018, 28, 1673–1682. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; García-Hernández, J.E. Diversity and distribution of stony corals. In Marine Biodiversity of Bonaire: A Baseline Study, 2nd ed.; Hoeksema, B.W., Ed.; Naturalis Biodiversity Center: Leiden, The Netherlands, 2022; pp. 17–19. [Google Scholar]
- De Jong, K.M.; Coomans, H.E. Marine gastropods from Curaçao, Aruba and Bonaire. Stud. Fauna Curaçao Caribb. Isl. 1988, 69, 1–261. Available online: https://repository.naturalis.nl/pub/506113 (accessed on 11 September 2022).
- Whalen, K.E.; Starczak, V.R.; Nelson, D.R.; Goldstone, J.V.; Hahn, M.E. Cytochrome P450 diversity and induction by gorgonian allelochemicals in the marine gastropod Cyphoma gibbosum. BMC Ecol. 2010, 10, 24. [Google Scholar] [CrossRef]
- Williams, E.H.; Bunkley-Williams, L. Marine major ecological disturbances of the Caribbean. Infect. Dis. Rev. 2000, 2, 110–127. [Google Scholar]
- Hector, A.; Hooper, R. Ecology: Darwin and the first ecological experiment. Science 2002, 295, 639–640. [Google Scholar] [CrossRef]
- Shaver, E.C.; Burkepile, D.E.; Silliman, B.R. Local management actions can increase coral resilience to thermally-induced bleaching. Nat. Ecol. Evol. 2018, 2, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- De Bakker, D.M.; van Duyl, F.C.; Perry, C.T.; Meesters, E.H. Extreme spatial heterogeneity in carbonate accretion potential on a Caribbean fringing reef linked to local human disturbance gradients. Glob. Change Biol. 2019, 25, 4092–4104. [Google Scholar] [CrossRef] [PubMed]








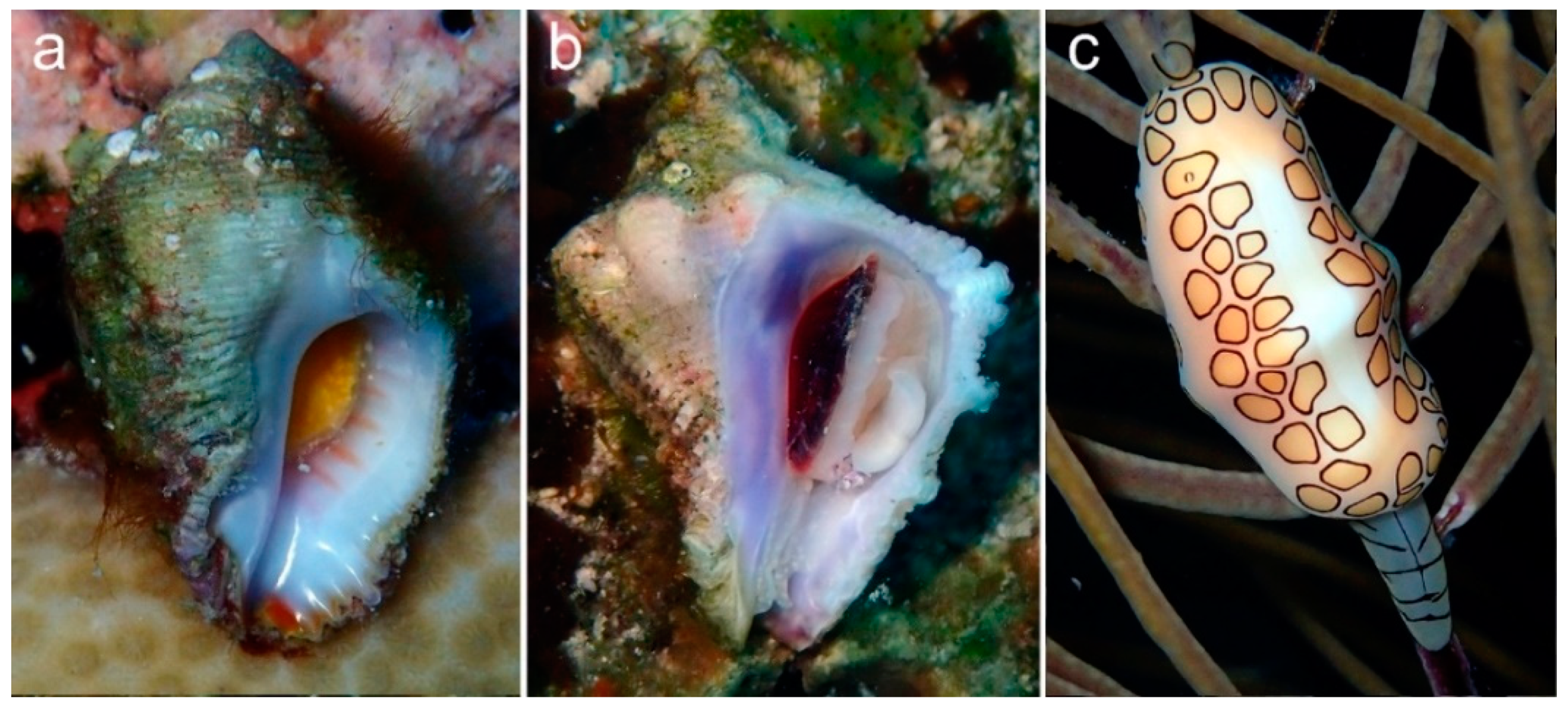{kind=link}
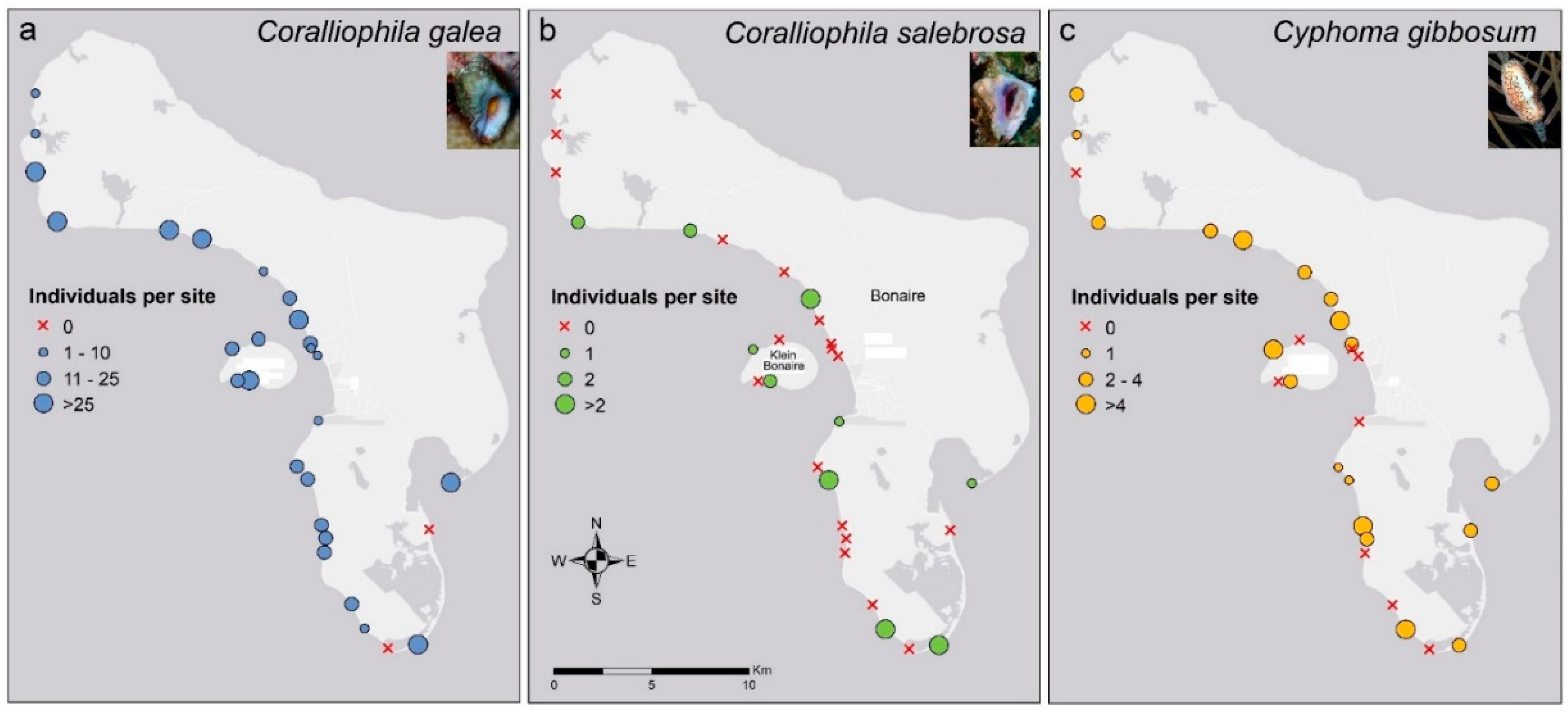{kind=link}
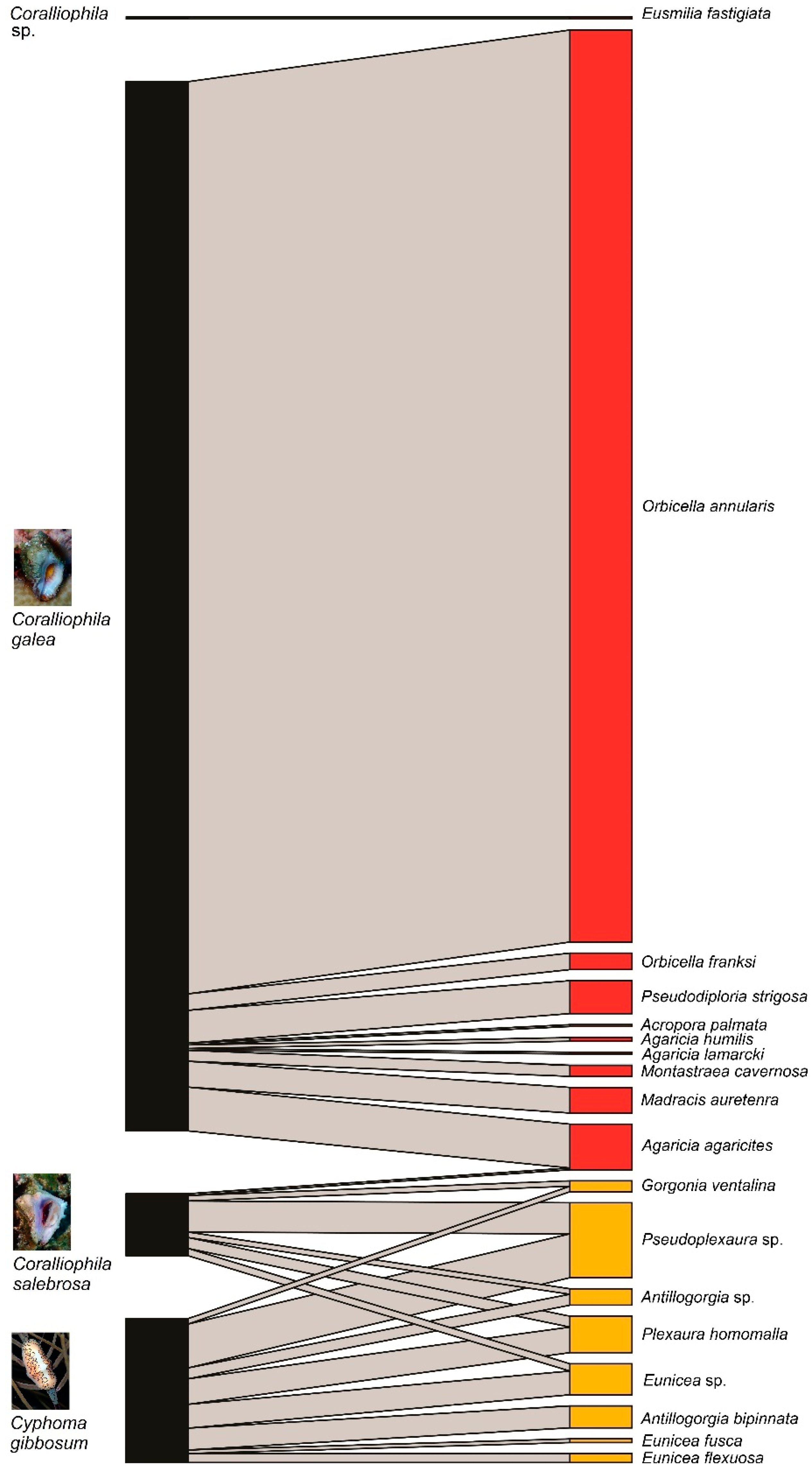{kind=link}
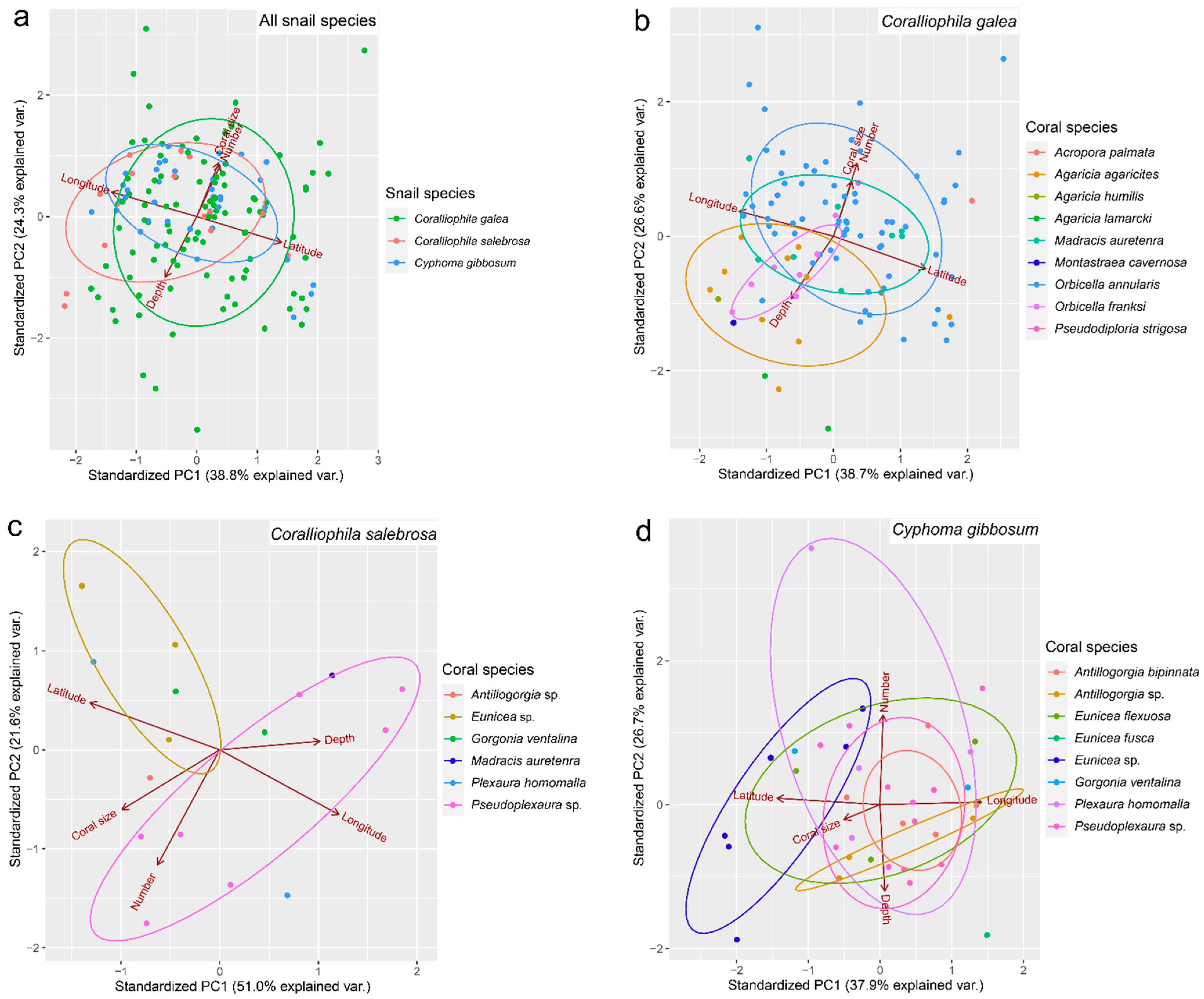{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Snail Species | ||||
|---|---|---|---|---|
| Host Coral Taxa | Coralliophila galea | Coralliophila salebrosa | Coralliophila sp. | Cyphoma gibbossum |
| Scleractinia | ||||
| Acroporidae | ||||
| Acropora palmata | 6 (1) | |||
| Agariciidae | ||||
| Agaricia agaricites | 14 (10) | |||
| Agaricia humilis | 1 (1) | |||
| Agaricia lamarcki | 2 (2) | |||
| Meandrinidae | ||||
| Eusmilia fastigiata | 1 (1) | |||
| Merulinidae | ||||
| Orbicella annularis | 500 (70) | |||
| Orbicella franksi | 18 (7) | |||
| Montastraeidae | ||||
| Montastraea cavernosa | 1 (1) | |||
| Mussidae | ||||
| Pseudodiploria strigosa | 9 (1) | |||
| Pocilloporidae | ||||
| Madracis auretenra | 24 (8) | 1 (1) | ||
| Octocorallia | ||||
| Gorgoniidae | ||||
| Antillogorgia bipinnata | 12 (7) | |||
| Antillogorgia sp. | 3 (1) | 6 (4) | ||
| Gorgonia ventalina | 3 (2) | 3 (2) | ||
| Plexauridae | ||||
| Eunicea flexuosa | 5 (3) | |||
| Eunicea fusca | 2 (1) | |||
| Eunicea sp. | 4 (3) | 13 (6) | ||
| Plexaura homomalla | 6 (2) | 14 (4) | ||
| Pseudoplexaura sp. | 17 (7) | 24 (13) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verboom, L.; Hoeksema, B.W. Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean). Diversity 2023, 15, 34. https://doi.org/10.3390/d15010034
Verboom L, Hoeksema BW. Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean). Diversity. 2023; 15(1):34. https://doi.org/10.3390/d15010034
Chicago/Turabian StyleVerboom, Lukas, and Bert W. Hoeksema. 2023. "Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean)" Diversity 15, no. 1: 34. https://doi.org/10.3390/d15010034
APA StyleVerboom, L., & Hoeksema, B. W. (2023). Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean). Diversity, 15(1), 34. https://doi.org/10.3390/d15010034