
Taxonomic Delimitation of the Monostromatic Green Algal Genera Monostroma Thuret 1854 and Gayralia Vinogradova 1969 (Ulotrichales, Chlorophyta)

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
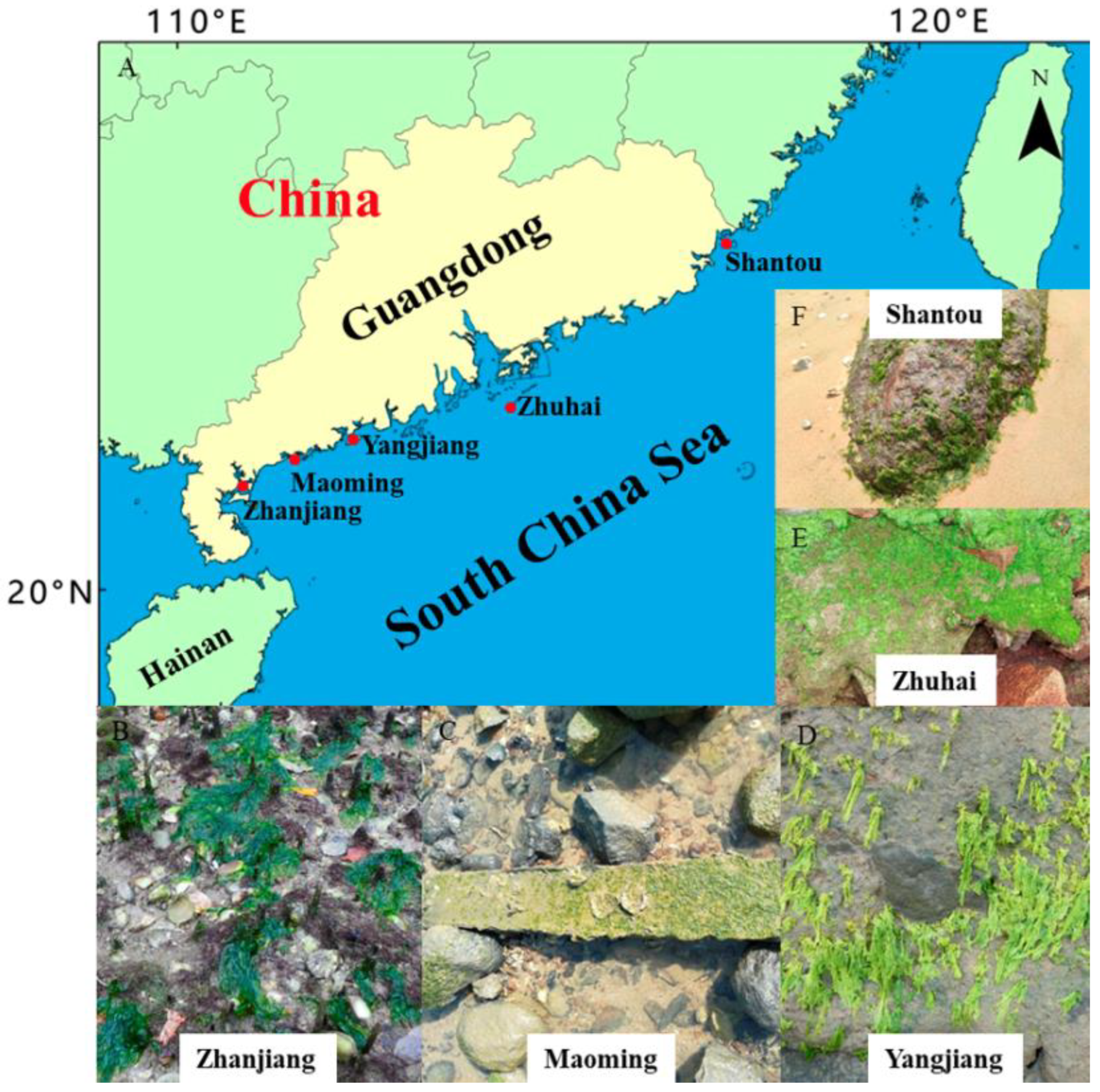
2.1. Monostromatic Green Algal Collection
2.2. Morphological Examination
2.3. DNA Extraction
2.4. PCR Amplification
2.5. Phylogenetic Analysis
3. Results
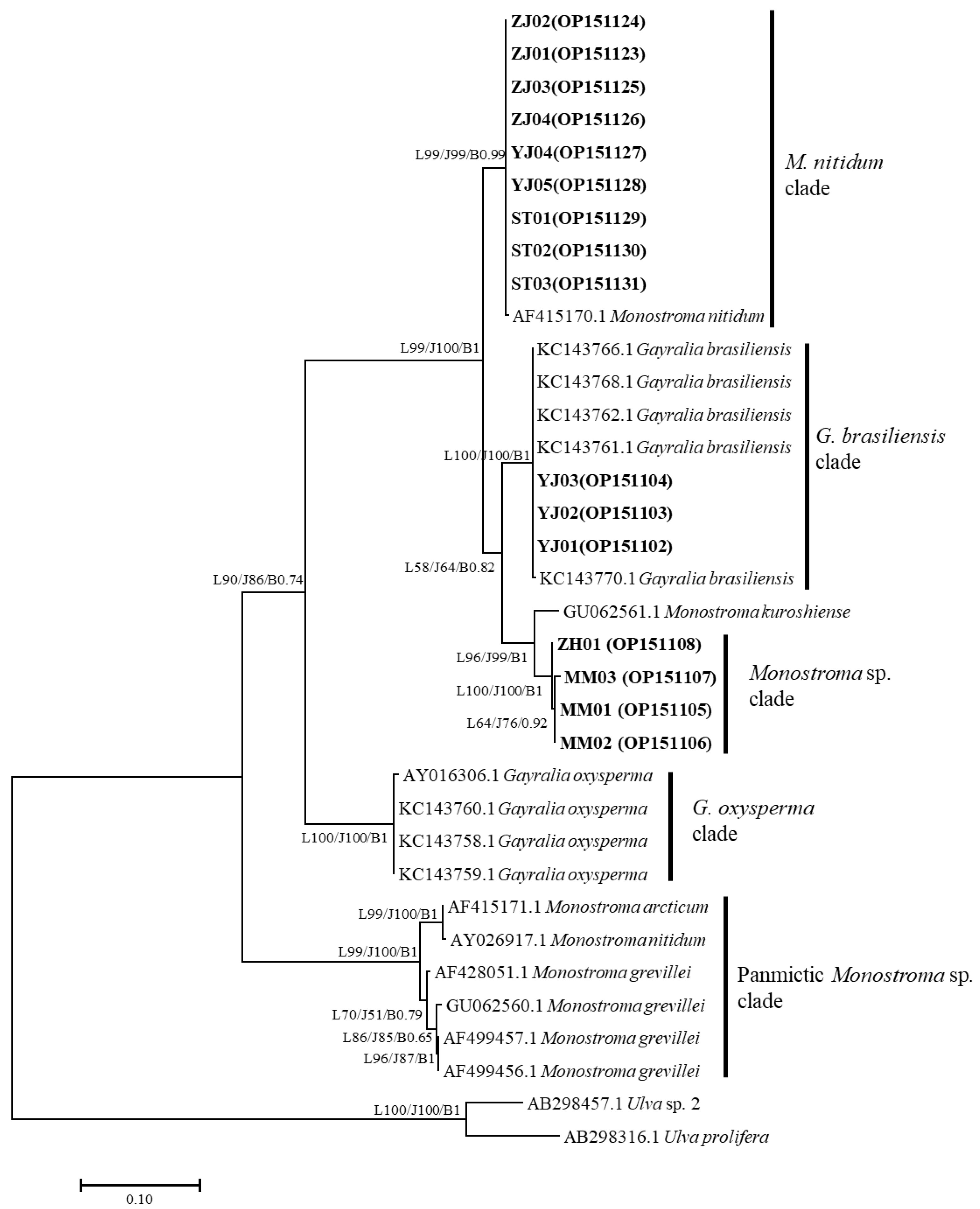
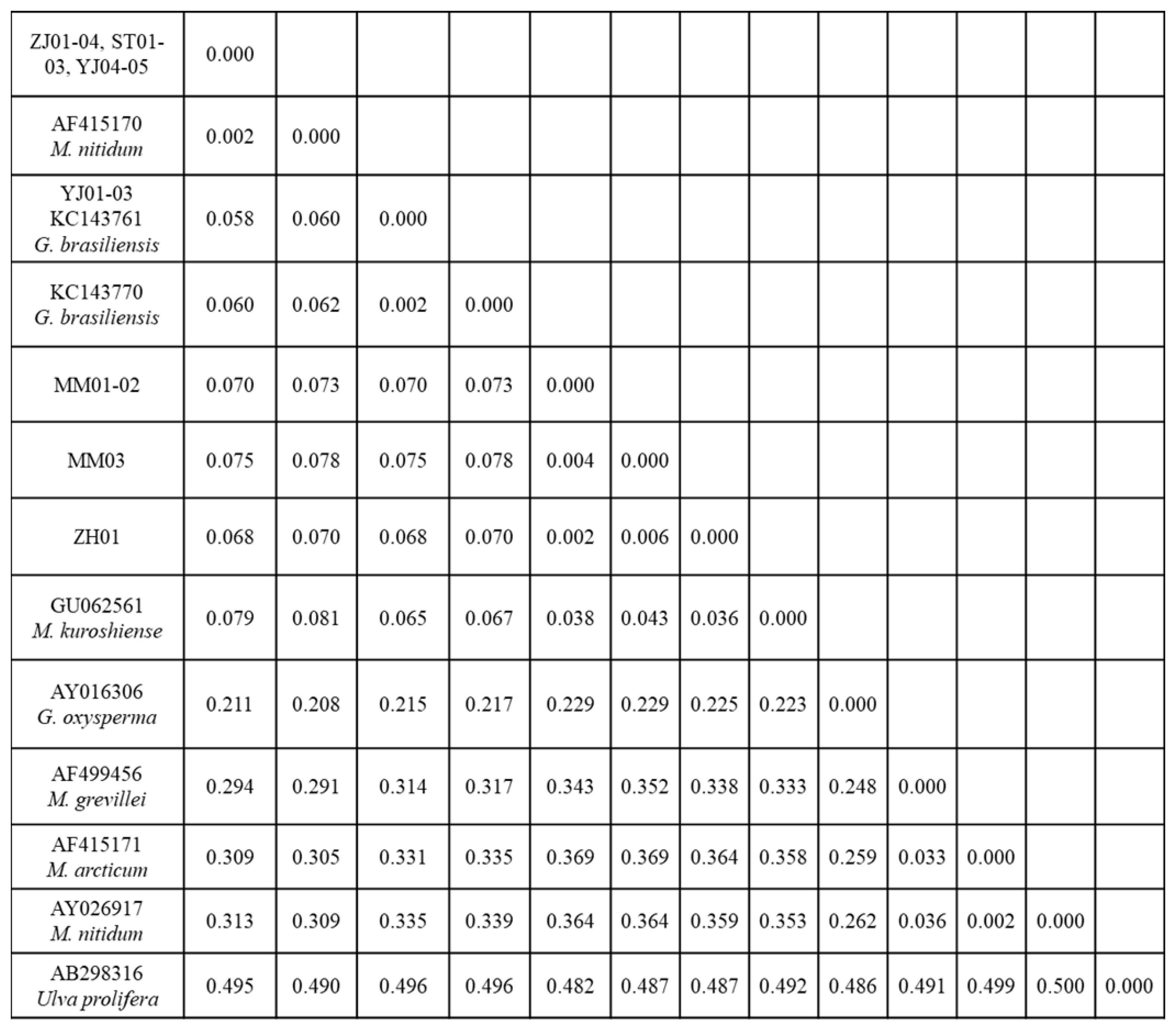
3.1. Phylogenetic Analyses
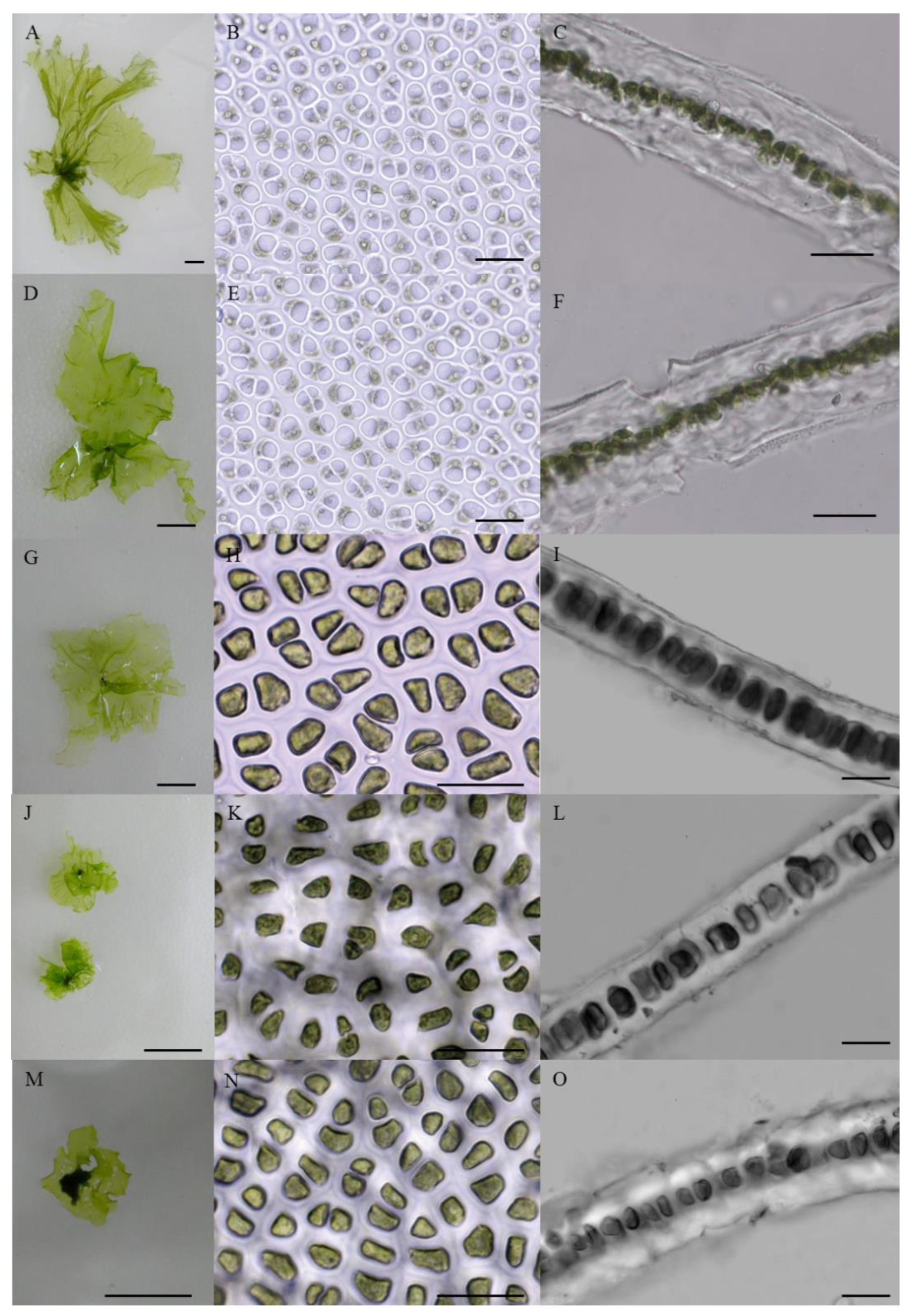
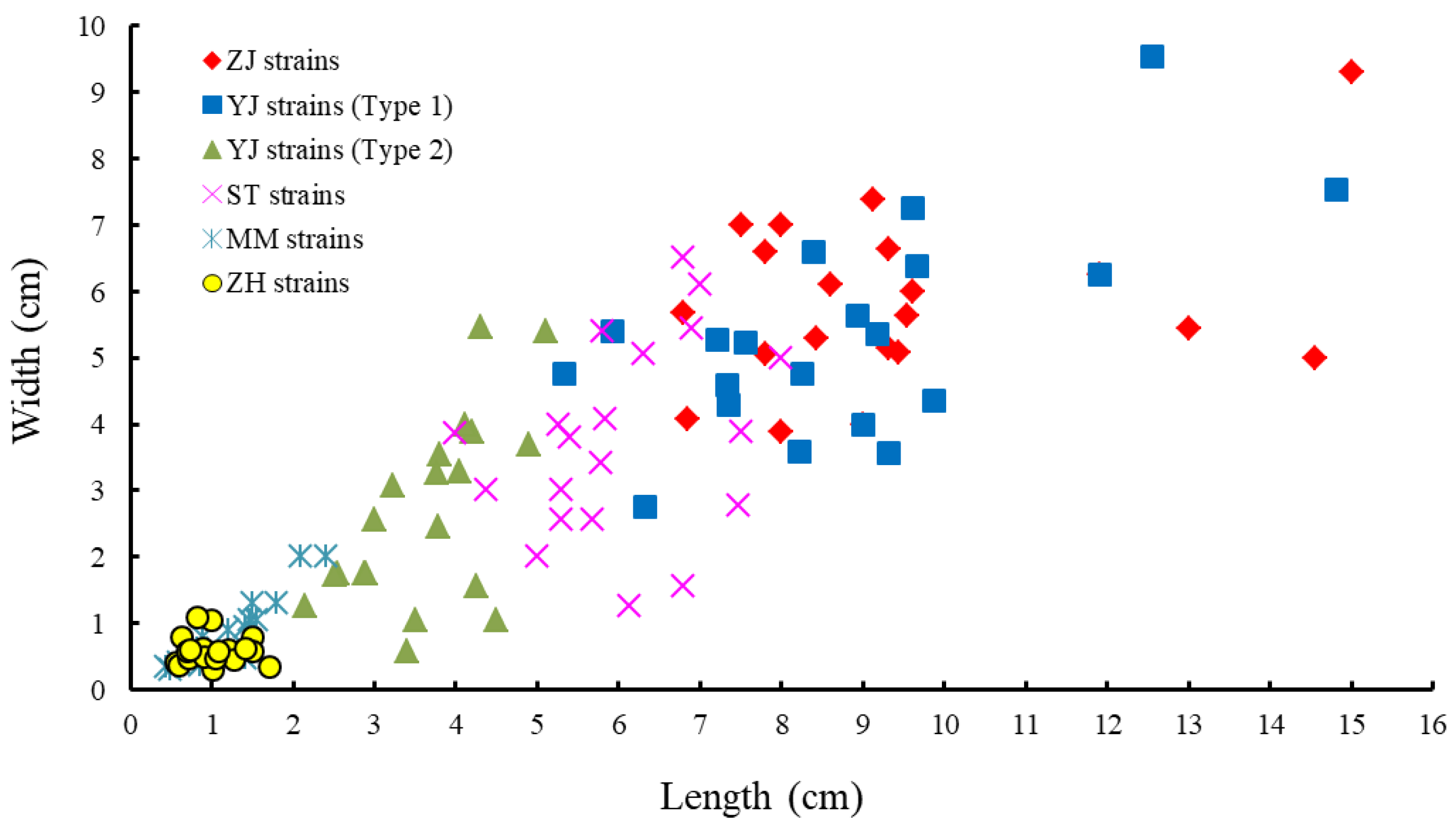
3.2. Morphology of Monostromatic Green Algae
4. Discussion
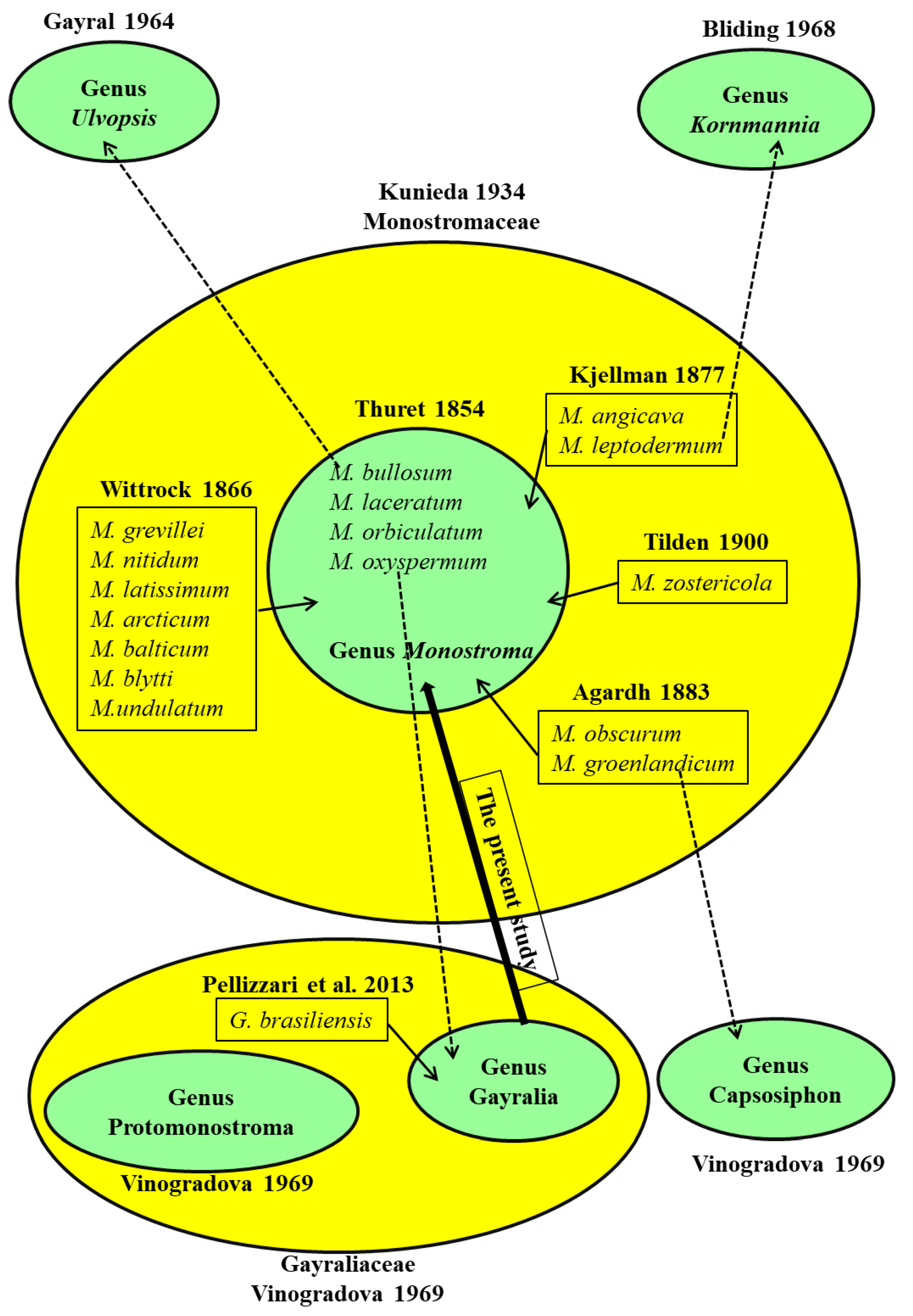
4.1. Taxonomic History of Monostromatic Green Algae
4.2. Taxonomic Assessment of Monostromatic Green Algae
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; De Clerck, O. Taylor & francis online: Phylogeny and molecular evolution of the green algae. Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar]
- Kunieda, H. On the life history of Monostroma. Proc. Imp. Acad. 1934, 10, 103–106. [Google Scholar] [CrossRef]
- Vinogradova, K.L. Sistematike poryadka Ulvales (Chlorophyta). Botanicheskii Zhurnal. 1969, 54, 1347–1355. [Google Scholar]
- Bast, F. Monostroma: The Jeweled Seaweed for Future; Lap Lambert Academic Publishing: Chisinau, Moldova, 2011; pp. 1–192. [Google Scholar]
- Pellizzari, F.; Oliveira, M.C.; Amanda, D.S.M.; Yokoya, N.S.; Oliveira, E.C. Morphology, ontogeny, and phylogenetic position of Gayralia brasiliensis sp. nov. (ulotrichales, chlorophyta) from the southern coast of brazil. Bot. Mar. 2013, 56, 197–205. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M.; AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 22 September 2021).
- Ohno, M. Cultivation of Monostroma nitidum (Chlorophyta) in a river estuary, southern Japan. J. Appl. Phycol. 1995, 7, 207–213. [Google Scholar] [CrossRef]
- Pellizzari, F.M.; Absher, T.; Yokoya, N.; Oliveira, E.C. Cultivation of the edible green seaweed Gayralia (Chlorophyta) in Southern Brazil. J. Appl. Phycol. 2007, 19, 63–69. [Google Scholar] [CrossRef]
- Pise, N.M.; Verlecar, X.N.; Gaikwad, D.K.; Jagtap, T.G. Nutraceutical properties of the marine macroalga Gayralia oxysperma. Bot. Mar. 2012, 55, 581–589. [Google Scholar]
- Cui, J.J.; Wang, H.; Chen, C.L.; Chen, X.Y.; Chen, Y.H.; Weng, M.X.; Huang, B.W.; Huang, Y.J.; Zhu, D.W.J.; Zhang, N.N.; et al. Formation and early development of the Monostroma nitidum Wittrock. Aquacult. Rep. 2021, 20, 100759. [Google Scholar] [CrossRef]
- Cassolato, J.E.; Noseda, M.D.; Pujol, C.A.; Pellizzari, F.M.; Damonte, E.B.; Duarte, M.E. Chemical structure and antiviral activity of the sulfated heterorhamnan isolated from the green seaweed Gayralia oxysperma. Carb. Res. 2008, 343, 3085–3095. [Google Scholar]
- Zhang, H.J.; Mao, W.J.; Fang, F.; Li, H.Y.; Sun, H.H.; Yin, C.; Qi, X.H. Chemical characteristics and anticoagulant activities of a sulfated polysaccharide and its fragments from Monostroma latissimum. Carbohydr. Polym. 2008, 71, 428–434. [Google Scholar] [CrossRef]
- Chen, Y.C. A simple method for isolating filaments as “algal seed stock” from monostroma latissimum (chlorophyta) germlings, and application for mass cultivation. J. Phycol. 2012, 48, 246–247. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, Y.; Togashi, T. Identification of genomic differences between the sexes and sex-specific molecular markers in Monostroma angicava (Ulvophyceae). J. Phycol. 2021, 57, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Thuret, G. Note sur la synonymie des Ulva lactuca et latissima L. suivie de quelques remarques sur la tribu des Ulvacees. Mem. Soc. Sci. Nat. Cherbourg. 1854, 2, 17–32. (In French) [Google Scholar]
- Papenfuss, G.F. On the genera of Ulvales and the status of the order. Bot. J. Linn. Soc. 1960, 56, 303–318. [Google Scholar] [CrossRef]
- Wittrock, V.B. Försök till en Monographi öfver AlgsläGtet Monostroma. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 1866; p. 66. (In Swedish). [Google Scholar]
- Kornmann, P. Über Monostroma bullosum (Roth) Thuret und M. oxyspermum (Kützing) Doty. Helgol. Wiss. Meeresuntersuch. 1964, 11, 13–21. (In German) [Google Scholar] [CrossRef]
- Bliding, C.A. Critical survey of European taxa in Ulvales, Part II: Ulva, Ulvaria, Monostroma, Kornmannia. Bot. Not. 1968, 121, 535–629. [Google Scholar]
- South, G.R.; Skelton, P.A. Catalogue of the marine benthic macroalgae of the Fiji, Islands, South Pacific. Aust. Syst. Bot. 2003, 16, 699–758. [Google Scholar] [CrossRef]
- Cui, J.J.; Monotilla, A.P.; Zhu, W.R.; Takano, Y.; Shimada, S.; Ichihara, K.; Matsui, T.; He, P.M.; Hiraoka, M. Taxonomic reassessment of Ulva prolifera (Ulvophyceae, Chlorophyta) based on specimens from the type locality and Yellow Sea green tides. Phycologia 2018, 57, 692–704. [Google Scholar] [CrossRef]
- Calderon, M.S.; Bustamante, D.E.; Gabrielson, P.W.; Martone, P.T.; Hind, K.R.; Huber, S.; Mansilla, A. Type specimen sequencing, multilocus analyses, and species delimitation methods recognize the cosmopolitan Corallina berteroi and establish the northern Japanese C. yendoi sp. nov. (Corallinaceae, Rhodophyta). J. Phycol. 2021, 57, 1659–1672. [Google Scholar] [CrossRef]
- Woolcott, G.W.; King, R.J. Ulvaria (Ulvales, Chlorophyta) in eastern Australia: Morphology, anatomy and ontogeny compared with molecular data. Bot. Mar. 1998, 41, 63–76. [Google Scholar] [CrossRef]
- Hiraoka, M.; Ichihara, K.; Zhu, W.R.; Shimada, S.; Oka, N.; Cui, J.J.; Tsubaki, S.; He, P.M. Examination of species delimitation of ambiguous DNA-based Ulva (Ulvophyceae, Chlorophyta) clades by culturing and hybridisation. Phycologia 2017, 56, 517–532. [Google Scholar] [CrossRef]
- Wang, S.Y.; Huo, Y.Z.; Zhang, J.H.; Cui, J.J.; Wang, Y.; Yang, L.L.; Zhou, Q.Y.; Lu, Y.W.; Yu, K.F.; He, P.M. Variations of dominant free-floating Ulva species in the source area for the world’s largest macroalgal blooms, China: Differences of ecological tolerance. Harmful Algae 2018, 74, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Gaysina, L.A.; Johansen, J.R.; Saraf, A.; Allaguvatova, R.Z.; Pal, S.; Singh, P. Roholtiella volcanica sp. nov., a New Species of Cyanobacteria from Kamchatkan Volcanic Soils. Diversity 2022, 14, 620. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, X.; Pang, W.; Wang, Q. Phylogeny of Trachelomonas and Strombomonas (Euglenaceae) Based on Morphological and Molecular Data. Diversity 2022, 14, 623. [Google Scholar] [CrossRef]
- Lorenzo-Carballa, M.O.; Sanmartín-Villar, I.; Cordero-Rivera, A. Molecular and Morphological Analyses Support Different Taxonomic Units for Asian and Australo-Pacific Forms of Ischnura aurora (Odonata, Coenagrionidae). Diversity 2022, 14, 606. [Google Scholar] [CrossRef]
- Yan, Q.; Dai, Q.; Liu, B.; Liu, G.; Zhu, H. A New Deep-Water Epilithic Green Alga, Ulvella lacustris, from an Alpine Brackish Lake in Qinghai–Tibet Plateau. Diversity 2022, 14, 594. [Google Scholar] [CrossRef]
- Li, F.; Dong, M.; Zhang, N.; Zhang, Y.; Li, Q.; Qian, Z.; Lian, Q.; Luo, J.; Huang, X.; Li, C. Euchlorocystis marina sp. nov. (Oocystaceae, Trebouxiophyceae), a New Species of Green Algae from a Seawater Shrimp Culture Pond. Diversity 2022, 14, 119. [Google Scholar] [CrossRef]
- Su, Q. Molecular Systematic Studies of Several Seaweeds in China. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2002; p. 81. [Google Scholar]
- Blomster, J.; Back, S.; Fewer, D.P.; Kiirikki, M.; Lehvo, A.; Maggs, C.A.; Stanhope, M.J. Novel morphology of Enteromorpha forming green tides. Am. J. Bot 2002, 89, 1756–1763. [Google Scholar] [CrossRef]
- Shimada, S.; Yokoyama, N.; Arai, S.; Hiraoka, M. Phylogeography of the genus Ulva (Ulvophyceae, Chlorophyta), with special reference to the Japanese freshwater and brackish taxa. J. Appl. Phycol 2008, 20, 979–989. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huellsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Suchard, M.; Drummond, A.J. Tracer, Version 1.6. 2013. Available online: https://tree.bio.ed.ac.uk/software/tracer (accessed on 28 July 2016).
- Kjellman, F.R. Ueber die Algenvegetationen des Murmanschen Meeres av der Westküste von Nowaja Semlja und Wajgatsch. Nova Acta Regiae Soc. Sci. Ups. 1877, 3, 1–86. (In German) [Google Scholar]
- Agardh, J.G. Till algernes systematik. Nya bidrag. (Tredje afdelningen). Lunds Univ. Års-Skr. Afdelningen Math. Och Nat. 1883, 19, 1–177. (In Swedish) [Google Scholar]
- Tilden, J.E. American Algae; University of Michigan: Minneapolis, MN, USA, 1900; Volume 4, pp. 301–400. [Google Scholar]
- Gayral, P. Sur le démembrement de l’actual genre Monostroma Thuret (Chlorophycées, Ulotrichales s.l.). C. R. Acad. Sci. Paris 1964, 258, 2149–2152. [Google Scholar]
- Tatewaki, M. Culture Studies on the Life History of Some Species of the Genus Monostroma; Scientific Papers of the Institute of Algological Research; Hokkaido University: Sapporo, Japan, 1969; Volume 6, pp. 1–56. [Google Scholar]
- Bast, F.; Shimada, S.; Hiraoka, M.; Okuda, K. Asexual life history by biflagellate zoids in Monostroma latissimum (Ulotrichales). Aqua. Bot. 2009, 91, 213–218. [Google Scholar] [CrossRef]
- Mayden, R.L. A hierarchy of species concepts: The denouement in the saga of the species problem. In Species: The Units of Biodiversity; Claridge, M.F., Dawah, H.A., Wilson, M.R., Eds.; Chapman and Hall: London, UK, 1997; pp. 381–424. [Google Scholar]
- Baldwin, B.G. Phylogenetic utility of the Internal Transcribed Spacers of nuclear ribosomal DNA in plants: An example from the Compositae. Mol. Phyl. Evol. 1992, 1, 3–16. [Google Scholar] [CrossRef]
- Karnjanapratum, S.; You, S.G. Molecular characteristics of sulfated polysaccharides from Monostroma nitidum and their in vitro anticancer and immunomodulatory activities. Int. J. Biol. Macromol. 2011, 48, 311–318. [Google Scholar] [CrossRef]
- Kawashima, Y.; Akasaki, T.; Matsumoto, Y.; Yamazaki, Y.; Shimada, S. Species identification of imported and Japanese commercial green algal products based on phylogenetic analyses using the nrITS2 and 5s rDNA spacer regions. Fisheries. Sci. 2013, 79, 521–529. [Google Scholar] [CrossRef]
- Wang, D.; Ma, J.; Du, J.; Xie, E.; Shi, J. Morphological and molecular study of Monostroma nitidum. Acta Agric. Zhejiangensis. 2015, 27, 1593–1600, (In Chinese with English Abstract). [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Strain Code | Collection Locality | Collection Date | Accession No. of ITS | Reference |
|---|---|---|---|---|---|
| Monostromatic species | ZJ01 | Dalang, Naozhou Island, Zhanjiang, China | 05-May-2020 | OP151123 | This study |
| ZJ02 | Dalang, Naozhou Island, Zhanjiang, China | 05-May-2020 | OP151124 | This study | |
| ZJ03 | Techeng Island, Zhanjiang, China | 15-May-2020 | OP151125 | This study | |
| ZJ04 | Techeng Island, Zhanjiang, China | 15-May-2020 | OP151126 | This study | |
| MM01 | Xiaofangji Island, Maoming, China | 19-Feb-2020 | OP151105 | This study | |
| MM02 | Xiaofangji Island, Maoming, China | 19-Feb-2020 | OP151106 | This study | |
| MM03 | Xiaofangji Island, Maoming, China | 19-Feb-2020 | OP151107 | This study | |
| YJ01 | Yinglu, Hailing Island, Yangjiang, China | 10-Feb-2020 | OP151102 | This study | |
| YJ02 | Yinglu, Hailing Island, Yangjiang, China | 10-Feb-2020 | OP151103 | This study | |
| YJ03 | Yinglu, Hailing Island, Yangjiang, China | 10-Feb-2020 | OP151104 | This study | |
| YJ04 | Mali, Hailing Island, Yangjiang, China | 01-Nov-2019 | OP151127 | This study | |
| YJ05 | Mali, Hailing Island, Yangjiang, China | 01-Nov-2019 | OP151128 | This study | |
| ZH01 | Miaowan Island, Zhuhai, China | 18-Mar-2021 | OP151108 | This study | |
| ST01 | Nanao Island, Shaotou, China | 23-Mar-2021 | OP151129 | This study | |
| ST02 | Nanao Island, Shaotou, China | 23-Mar-2021 | OP151130 | This study | |
| ST03 | Nanao Island, Shaotou, China | 23-Mar-2021 | OP151131 | This study | |
| Monostroma nitidum | / | Bailongwei, Fangcheng, Guangxi, China | / | AF415170 | [31] |
| / | / | / | AY026917 | [5] | |
| Monostroma kuroshiense | / | Sakurajima, Kagoshima Pref, Japan | Mar/Apr-2009 | GU062561 | [4] |
| Monostroma arcticum | / | Shimiao, Dalian, China | / | AF415171 | [31] |
| Monostroma grevillei | / | Donegal, Ireland | Mar/Apr-2009 | GU062560 | [4] |
| / | Xiaoping Island, Dalian, China | / | AF428051 | [31] | |
| / | Atlantic | / | AF499456 | [32] | |
| / | Baltic | / | AF499457 | [32] | |
| Gayralia brasiliensis | / | Florianópolis, Santa Catarina, Brazil | 01-Jul-2006 | KC143761 | [5] |
| / | Guaratuba Bay, Paraná, Brazil | 11-Aug-2006 | KC143762 | [5] | |
| / | Piúna Beach, Piúna, Espírito Santo, Brazil | 12-Sep-2007 | KC143766 | [5] | |
| / | Caravelas, Bahia, Brazil | 26-Jan-2008 | KC143768 | [5] | |
| / | Itapissuma, Pernambuco, Brazil | 09-May-2007 | KC143770 | [5] | |
| Gayralia oxysperma | / | / | / | AY016306 | [5] |
| / | Antonina Bay, Paraná, Brazil | 09-Jul-2006 | KC143758 | [5] | |
| / | Lagoinha Beach, São Paulo, Brazil | 07-Aug-2005 | KC143759 | [5] | |
| / | Laguna, Santa Catarina, Brazil | 23-Nov-2010 | KC143760 | [5] | |
| Ulva prolifera Ulva sp.2 | / | Sekiguchi R., Yamada, Iwate, Japan | 02-May-2005 | AB298316 | [33] |
| / | Yoshino River, Tokushima, Japan | 18-Jul-2000 | AB298457 | [33] |
| Sample Name | Thallus Morphology | |||
|---|---|---|---|---|
| Percentage of Cells with 1 to 2 Pyrenoids a | Cell Size (Length × Width μm) b | Thallus Thickness (μm) b | ||
| 1 | 2 | |||
| YJ02 | 96% | 4% | 9.06 ± 1.36 × 6.06 ± 0.68 | 25.2 ± 1.9 |
| YJ05 | 98% | 2% | 11.47 ± 2.13 × 8.42 ± 1.72 | 33.8 ± 2.7 |
| ST01 | 98% | 2% | 11.49 ± 2.06 × 8.44 ± 1.52 | 34.2 ± 2.3 |
| MM01 | 96% | 4% | 7.18 ± 1.41 × 5.47 ± 1.04 | 24.9 ± 1.6 |
| ZH01 | 96% | 4% | 7.15 ± 1.28 × 5.43 ± 1.02 | 26.3 ± 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, J.; Chen, C.; Tan, H.; Huang, Y.; Chen, X.; Xin, R.; Liu, J.; Huang, B.; Xie, E. Taxonomic Delimitation of the Monostromatic Green Algal Genera Monostroma Thuret 1854 and Gayralia Vinogradova 1969 (Ulotrichales, Chlorophyta). Diversity 2022, 14, 773. https://doi.org/10.3390/d14090773
Cui J, Chen C, Tan H, Huang Y, Chen X, Xin R, Liu J, Huang B, Xie E. Taxonomic Delimitation of the Monostromatic Green Algal Genera Monostroma Thuret 1854 and Gayralia Vinogradova 1969 (Ulotrichales, Chlorophyta). Diversity. 2022; 14(9):773. https://doi.org/10.3390/d14090773
Chicago/Turabian StyleCui, Jianjun, Chunli Chen, Huaqiang Tan, Yongjian Huang, Xinyi Chen, Rong Xin, Jinlin Liu, Bowen Huang, and Enyi Xie. 2022. "Taxonomic Delimitation of the Monostromatic Green Algal Genera Monostroma Thuret 1854 and Gayralia Vinogradova 1969 (Ulotrichales, Chlorophyta)" Diversity 14, no. 9: 773. https://doi.org/10.3390/d14090773
APA StyleCui, J., Chen, C., Tan, H., Huang, Y., Chen, X., Xin, R., Liu, J., Huang, B., & Xie, E. (2022). Taxonomic Delimitation of the Monostromatic Green Algal Genera Monostroma Thuret 1854 and Gayralia Vinogradova 1969 (Ulotrichales, Chlorophyta). Diversity, 14(9), 773. https://doi.org/10.3390/d14090773