
The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Definition of Variables Used in This Study
2.3. Data Collection
2.4. Data Analysis
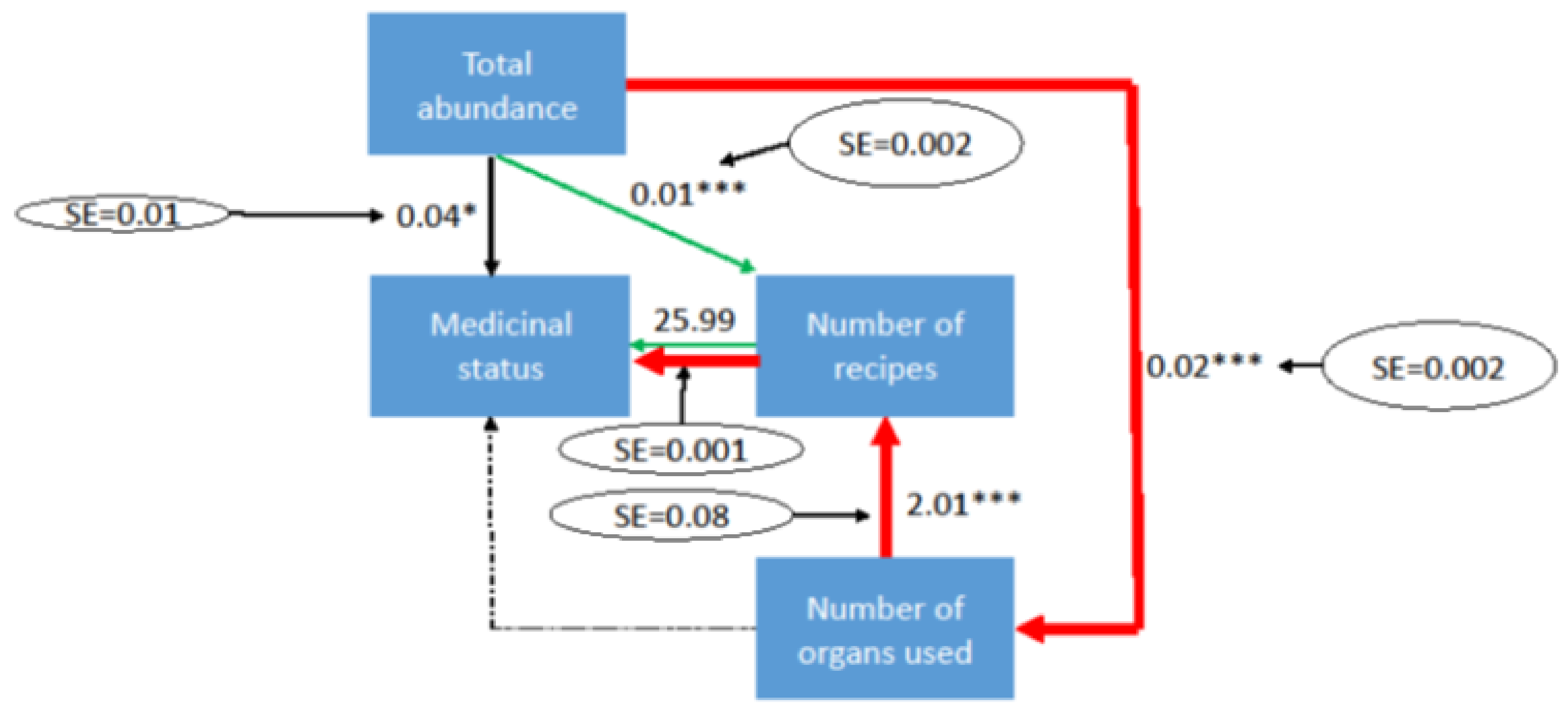
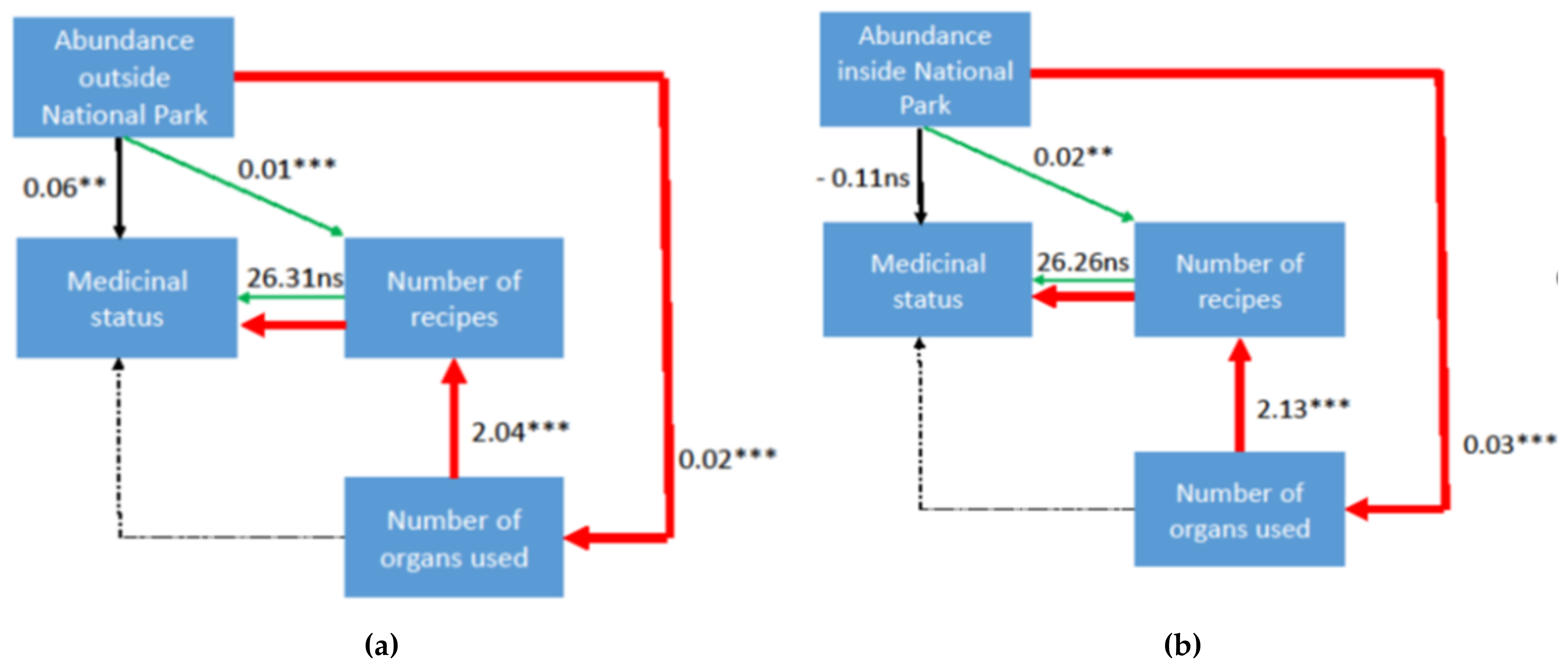
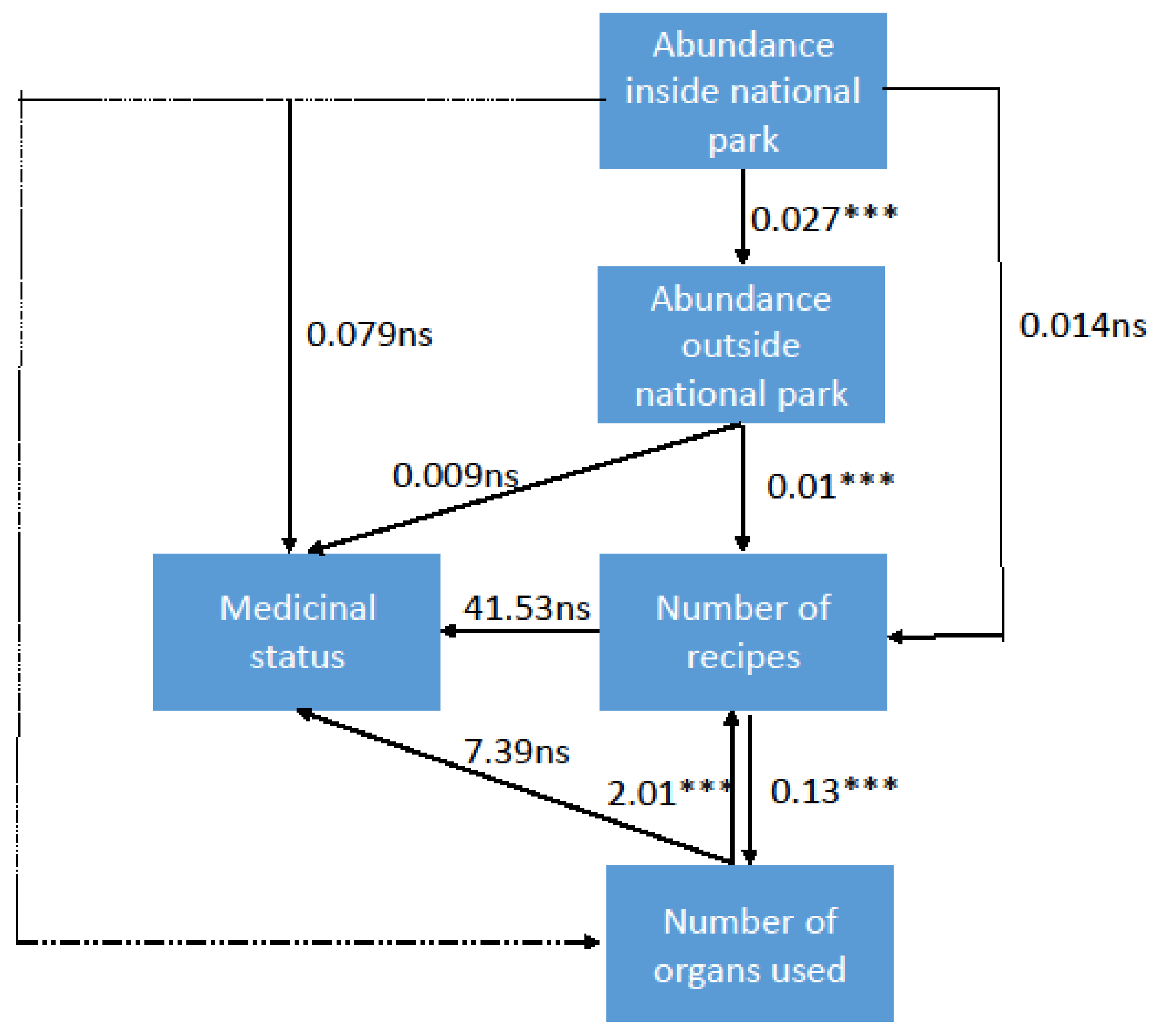
3. Results and Discussion
4. Way Forward: Ex-Situ Conservation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs Over the 30 Years from 1981 to 2012. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Schippmann, U.; Leaman, J.D.; Cunningham, A.B. Impact of Cultivation and Gathering of Medicinal Plants on Biodiversity: Global Trends and Issues; Inter-Departmental Working Group on Biological Diversity for Food and Agriculture: Rome, Italy, 2002. [Google Scholar]
- Voeks, R.A. Disturbance pharmacopoeias: Medicine and myth from the humid Tropics. Ann. Assoc. Am. Geogr. 2004, 94, 868–888. [Google Scholar]
- Albuquerque, U.P. Re-examining hypotheses concerning the use and knowledge of medicinal plants: A study in the Caatinga vegetation of NE Brazil. J. Ethnobiol. Ethnomed. 2006, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Saslis-Lagoudakis, C.H.; Hawkins, J.A.; Greenhill, S.J.; Pendry, C.A.; Watson, M.F.; Tuladhar-Douglas, W.; Baral, S.R.; Savolainen, V. The evolution of traditional knowledge: Environment shapes medicinal plant use in Nepal. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132768. [Google Scholar] [CrossRef] [PubMed]
- Yessoufou, K.; Ambani, A.E.; Elansary, H.O.; El-Sabrout, A.M.; Shokralla, S. Time, Mediated through Plant Versatility, Is a Better Predictor of Medicinal Status of Alien Plants. Diversity 2022, 14, 286. [Google Scholar] [CrossRef]
- de Oliveira Trindade, M.R.; Jardim, J.G.; Casas, A.; Guerra, N.M.; de Lucena, R.F.P. Availability and use of woody plant resources in two areas of Caatinga in Northeastern Brazil. Ethnobot. Res. Appl. 2015, 14, 313–957. [Google Scholar] [CrossRef]
- Gonçalves, P.H.S.; Albuquerque, U.P.; Medeiros, P.M. The most commonly available woody plant species are the most useful for human populations: A meta-analysis. Ecol. Appl. 2016, 26, 2238–2253. [Google Scholar] [CrossRef] [PubMed]
- DeFries, R.; Hansen, A.; Turner, B.L.; Reid, R.; Liu, J.G. Land use change around protected areas: Management to balance human needs and ecological function. Ecol. Appl. 2007, 17, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Dudley, N.; Stolton, S.; Belokurov, A.; Krueger, L.; Lopoukhine, N.; MacKinnon, K.; Sandwith, T.; Sekhran, N. Natural Solutions: Protected Areas Helping People Cope with Climate Change; IUCNWCPA: Gland, Switzerland; TNC: Washington, DC, USA; UNDP: New York, NY, USA; WCS: New York, NY, USA; The World Bank: Washington, DC, USA; WWF: Gland, Switzerland, 2010. [Google Scholar]
- Heywood, V.H. Conserving plants within and beyond protected areas–still problematic and future uncertain. Plant Divers. 2018, 41, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- Gülez, S. A method of evaluating areas for national park status. Environ. Manag. 1992, 16, 811–818. [Google Scholar] [CrossRef]
- Schmidt, E.; Lotter, M.; McCleland, W. Trees and Shrubs of Mpumalanga and Kruger National Park; Jacana Press: Johannesburg, South Africa, 2007; 702p. [Google Scholar]
- Lahaye, R.; van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [PubMed]
- Yessoufou, K.; Davies, T.J.; Maurin, O.; Kuzmina, M.; Schaefer, H.; van der Bank, M.; Savolainen, V. Large herbivores favour species diversity but have mixed impacts on phylogenetic community structure in an African savanna ecosystem. J. Ecol. 2013, 101, 614–625. [Google Scholar] [CrossRef]
- Perera, S.J.; Ratnayake-Perera, D.; Procheş, Ş. Vertebrate distributions indicate a Greater Maputaland-Pondoland-Albany region of endemism. S. Afr. J. Sci. 2011, 107, 49–63. [Google Scholar] [CrossRef][Green Version]
- Coates-Palgrave, M. Keith Coates-Palgrave. In Trees of Southern Africa, 3rd ed.; Struik Nature: Cape Town, South Africa, 2002. [Google Scholar]
- van Wyk, B.; van Wyk, P. Field Guide to Trees of Southern Africa, 2nd ed.; Struik Publisher: Cape Town, South Africa, 2013. [Google Scholar]
- Glen, H.; Van Wyk, B. Guide to Trees Introduced into Southern Africa; Penguin Random House: Cape Town, South Africa, 2016. [Google Scholar]
- Royal Museum for Central Africa. Prelude Database for Medicinal Plants in Africa. 2018. Available online: http://www.africamuseum.be/en/research/collections_libraries/biology/prelude (accessed on 25 January 2018).
- Germishuizen, G.; Meyer, N.L. Plants of Southern Africa: An Annotated Checklist; National Botanical Institute: Pretoria, South Africa, 2003. [Google Scholar]
- Boon, D. Pooley’s Trees of Eastern South Africa–A Complete Guide; Flora and Fauna Publication Trust: Durban, South Africa, 2010. [Google Scholar]
- Van Wyk, B.; van der Berg, E.; Coates-Palgrave, M.; Jordaan, M. Dictionary of Names for Southern African Trees, 1st ed.; Briza: Pretoria, South Africa, 2011. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Lefcheck, J.S. PIECEWISESEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Venter, F.J. A Classification of Land for Management Planning in the Kruger National Park. PhD Thesis, University of South Africa, Pretoria, South Africa, 2012. [Google Scholar]
- Ford, R.I. The Nature and Status of Ethnobotany; University of Michigan: Ann Arbor, MI, USA, 1978. [Google Scholar]
- Phillips, O.; Gentry, A.H. The useful plants of Tambopata, Peru: I. Statistical hypotheses tests with a new quantitative technique. Econ. Bot. 1993, 47, 15–32. [Google Scholar] [CrossRef]
- Bennett, B.C. Ethnobotany education, opportunities, and needs in the US. Ethnobot. Res. Appl. 2005, 3, 113–122. [Google Scholar] [CrossRef][Green Version]
- Martin, G.J. Ethnobotany: A Methods Manual; Earthscan: London, UK, 2007. [Google Scholar]
- Albuquerque, U.P.; Hanazaki, N. Commentary: Five Problems in Current Ethnobotanical Research—And Some Suggestions for Strengthening Them. Human Ecol. 2009, 37, 653–661. [Google Scholar] [CrossRef]
- Gaoue, O.G.; Coe, M.A.; Bond, M.; Hart, G.; Seyler, B.C.; McMillen, H. Theories and Major Hypotheses in Ethnobotany. Econ. Bot. 2017, 71, 269–287. [Google Scholar] [CrossRef]
- Robles Ariasa, D.M.; Cevallos, D.; Gaoue, O.; Fadimana, M.G.; Hindlea, T. Non-random medicinal plants selection in the Kichwa community of the Ecuadorian Amazon. J. Ethnopharmacol. 2020, 246, 112220. [Google Scholar] [CrossRef] [PubMed]
- CBD. Global Biodiversity Outlook 2; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 2006; p. 81. [Google Scholar]
- IUCN. IUCN Technical Guidelines on the Management of Ex Situ Populations for Conservation. In Proceedings of the 14th Meeting of the Programme Committee of Council, Gland, Switzerland, 10 December 2002. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yessoufou, K.; Muleba, I.; Rampedi, I.T. The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species. Diversity 2022, 14, 693. https://doi.org/10.3390/d14080693
Yessoufou K, Muleba I, Rampedi IT. The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species. Diversity. 2022; 14(8):693. https://doi.org/10.3390/d14080693
Chicago/Turabian StyleYessoufou, Kowiyou, Isidore Muleba, and Isaac T. Rampedi. 2022. "The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species" Diversity 14, no. 8: 693. https://doi.org/10.3390/d14080693
APA StyleYessoufou, K., Muleba, I., & Rampedi, I. T. (2022). The Test of the Availability Hypothesis Reveals the Needs for Ex-Situ Conservation for Some Protected Area-Restricted Species. Diversity, 14(8), 693. https://doi.org/10.3390/d14080693