
Impact of Stump Removal on Communities of Ectomycorrhizal and Other Soil Fungi in Norway Spruce Stands of Latvia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Work
2.2. Sample Preparation and Sequencing
2.3. Data Analyses
3. Results
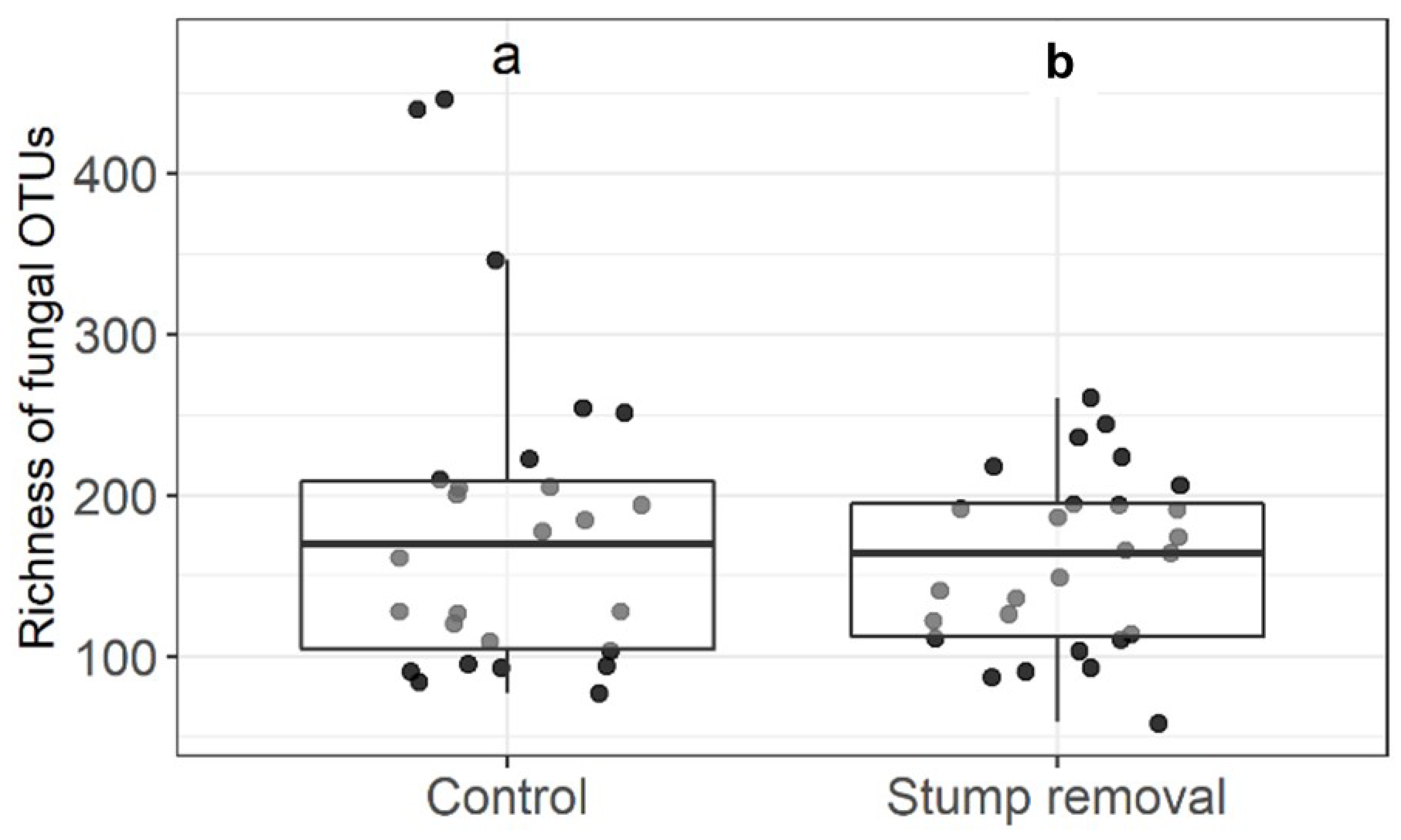
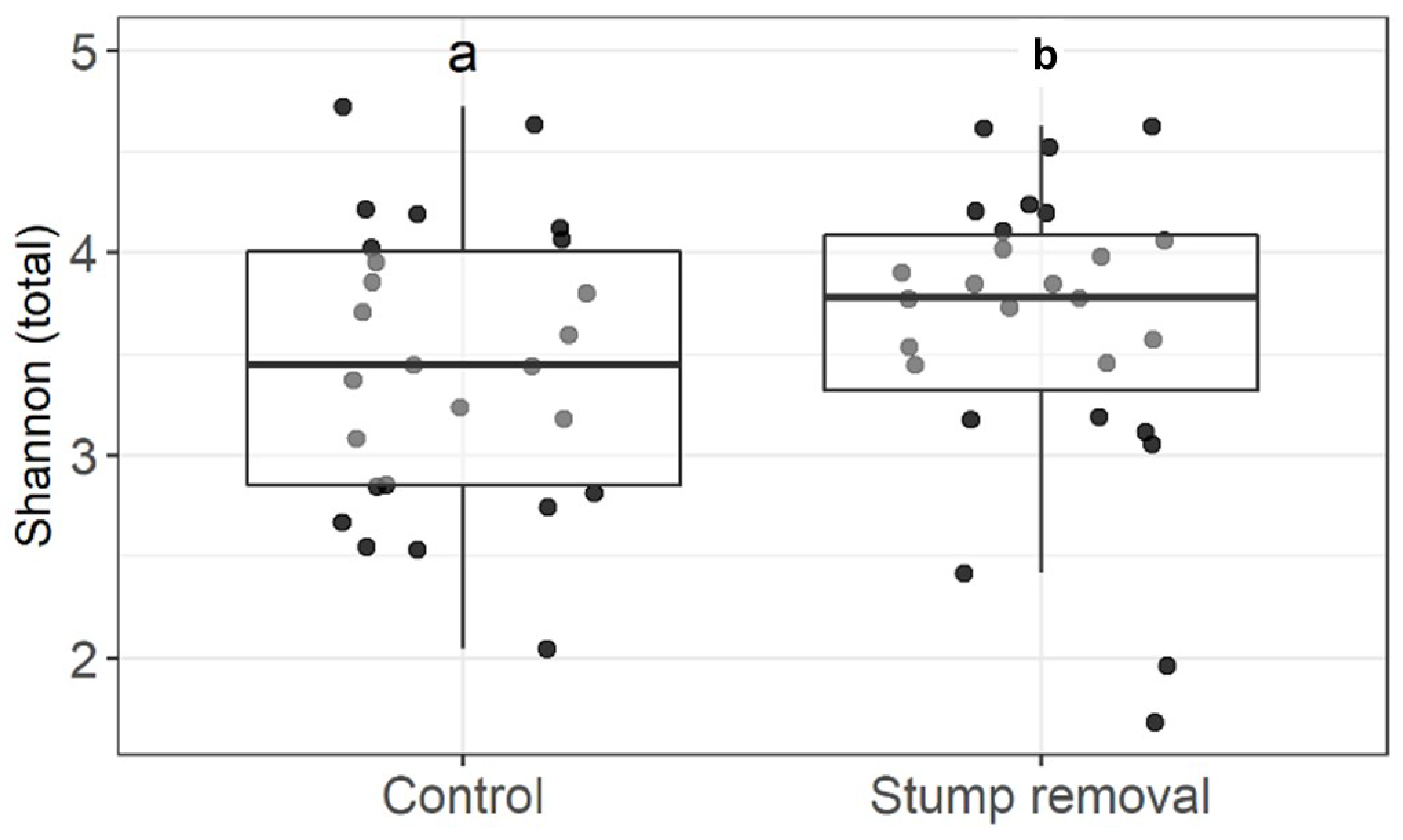
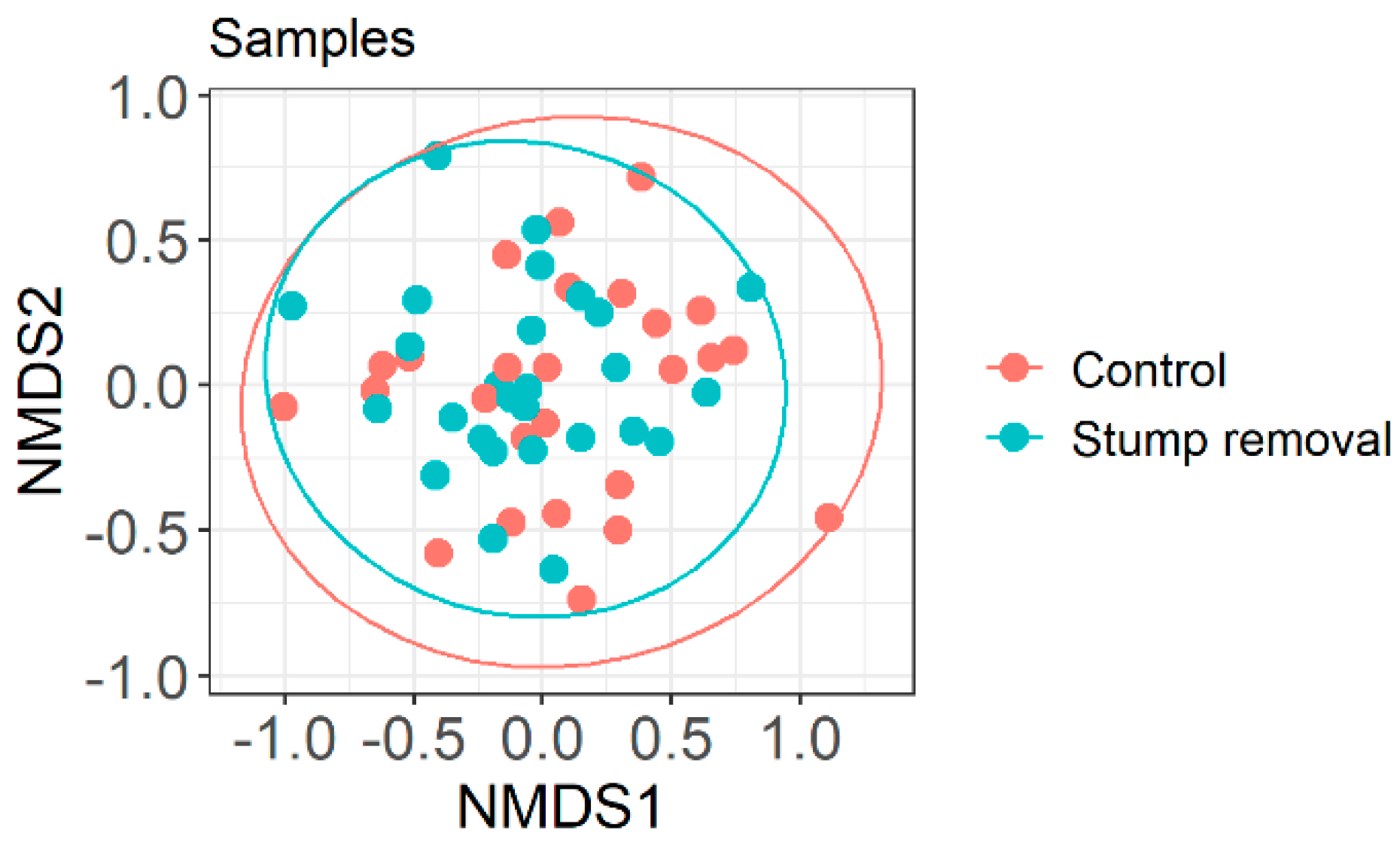
3.1. Impact of Stump Removal on Diversity of Soil and Mycorrhizal Fungi
3.2. Impact of Forest Site Type on Diversity of Soil and Mycorrhizal Fungi
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woodward, S.; Stenlid, J.; Karjalainen, R.A. Heterobasidion Annosum: Biology, Ecology, Impact and Control; CAB International: Wallingford, UK, 1998; 616p. [Google Scholar]
- Gonthier, P.; Thor, M. Annosus root and butt rots. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CAB International: Wallingford, UK, 2013; pp. 128–158. [Google Scholar]
- Stenlid, J.; Redfern, D.B. Spread within the tree and stand. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 125–141. [Google Scholar]
- Piri, T. The spreading of the S type of Heterobasidion annosum from Norway spruce stumps to subsequent tree stands. Eur. J. For. Pathol. 1998, 26, 193–204. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Bruna, L.; Zaluma, A.; Burnevica, N.; Klavina, D.; Legzdina, L.; Jansons, J.; Piri, T. Development of Heterobasidion spp. fruit bodies on decayed Picea abies. For. Ecol. Manag. 2021, 482, 118835. [Google Scholar] [CrossRef]
- Laivins, M. Geography of Norway spruce (Picea abies) stands in Latvia. Proc. Latv. Univ. Agric. 2005, 14, 1–9, (In Latvian with English Abstract). [Google Scholar]
- Arhipova, N.; Gaitnieks, T.; Donis, J.; Stenlid, J.; Vasaitis, R. Butt rot incidence, causal fungi, and related yield loss in Picea abies stands of Latvia. Can. J. For. Res. 2011, 41, 2337–2345. [Google Scholar] [CrossRef]
- Korhonen, K.; Stenlid, J. Biology of Heterobasidion annosum. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 43–70. [Google Scholar]
- Vasaitis, R.; Stenlid, J.; Thomsen, I.M.; Berklund, P.; Dahlberg, A. Stump removal to control root rot in forest stands. A literature study. Silva Fenn. 2008, 42, 457–483. [Google Scholar] [CrossRef]
- Cleary, M.R.; Arhipova, N.; Morrison, D.J.; Thomsen, I.M.; Sturrock, R.N.; Vasaitis, R.; Gaitnieks, T.; Stenlid, J. Stump removal to control root disease in Canada and Scandinavia: A synthesis of results from long-term trials. For. Ecol. Manag. 2013, 290, 5–14. [Google Scholar] [CrossRef]
- Aosaar, J.; Drenkhan, T.; Adamson, K.; Aun, K.; Becker, H.; Buht, M.; Drenkhan, R.; Fjodorov, M.; Jürimaa, K.; Morozov, G.; et al. The effect of stump harvesting on tree growth and the infection of root rot in young Norway spruce stands in hemiboreal Estonia. For. Ecol. Manag. 2020, 475, 118425. [Google Scholar] [CrossRef]
- Hope, G.D. Changes in soil properties, tree growth, and nutrition over a period of a 10 years after stump removal and scarification on moderately coarse soils in interior British Columbia. For. Ecol. Manag. 2007, 242, 625–635. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal biodiversity and their role in soil health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef]
- Powell, J.R.; Rillig, M.C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 2018, 220, 1059–1075. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomus, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Makiola, A.; Dickie, I.A.; Holdaway, R.J.; Wood, J.R.; Orwin, K.H.; Glare, T.R. Land use is a determinant of plant pathogen alpha-but not beta-diversity. Mol. Ecol. 2019, 28, 3786–3798. [Google Scholar] [CrossRef] [PubMed]
- Sterkenburg, E.; Clemmensen, K.E.; Lindahl, B.D.; Dahlberg, A. The significance of retention trees for survival of ectomycorrhizal fungi in clear-cut Scots pine forests. J. Appl. Ecol. 2019, 56, 1367–1378. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Harvey, A.E.; Jurgensen, M.F.; Amaranthus, M.P. Impacts of soil compaction and tree stump removal on soil properties and outplanted seedlings in northern Idaho, USA. Can. J. Soil Sci. 1998, 78, 29–34. [Google Scholar] [CrossRef]
- Menkis, A.; Uotila, A.; Arhipova, N.; Vasaitis, R. Effects of stump and slash removal on growth and mycorrhization of Picea abies seedlings outplanted on a forest clear-cut. Mycorrhiza 2010, 20, 505–509. [Google Scholar] [CrossRef]
- Kataja-aho, S.; Pennanen, T.; Lensu, A.; Haimi, J. Does stump removal affect early growth and mycorrhizal infection of spruce (Picea abies) seedlings in clear-cuts? Scand. J. For. Res. 2012, 27, 746–753. [Google Scholar] [CrossRef]
- Huusko, K.; Tarvainen, O.; Saravesi, K.; Pennanen, T.; Fritze, H.; Kubin, E.; Markkola, A. Short-term impacts of energy woods harvesting on ectomycorrhizal fungal communities of Norway spruce saplings. ISME J. 2015, 9, 581–591. [Google Scholar] [CrossRef]
- Kļaviņa, D.; Menkis, A.; Gaitnieks, T.; Pennanen, T.; Lazdiņš, A.; Velmala, S.; Vasaitis, R. Low impact of stump removal on mycorrhization and growth of replanted Picea abies: Data from three types of Hemiboreal forest. Balt. For. 2016, 22, 16. [Google Scholar]
- Modi, D.; Simard, S.; Bérubé, J.; Lavkulich, L.; Hamelin, R.; Grayston, S.J. Long-term effect of stump removal and tree species composition on the diversity and structure of soil fungal communities. FEMS Microbiol. Ecol. 2020, 96, fiaa061. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Drenkhan, R.; Pritsch, K.; Buegger, F.; Padari, A.; Hagh-Doust, N.; Mikryukov, V.; Gohar, D.; et al. Regional-Scale In-Depth Analysis of Soil Fungal Diversity Reveals Strong pH and Plant Species Effects in Northern Europe. Front. Microbiol. 2020, 11, 1953. [Google Scholar] [CrossRef]
- Burņeviča, N.; Zaļuma, A.; Kļaviņa, D.; Brūna, L.; Legzdiņa, L.; Gaitnieks, T. Initial and long-term fungal diversity and occurrence of Heterobasidion spp. in Norway spruce root fragments remaining in soil after stump extraction. Scand. J. For. Res. 2021, 36, 117–125. [Google Scholar] [CrossRef]
- Bušs, K. Forest ecosystem classification in Latvia. Proc. Latv. Acad. Sci. 1997, 51, 204–218. [Google Scholar]
- Zimelis, A.; Lazdiņš, A.; Sarmulis, Z. Comparison of productivity of CBI and MCR—500 stump lifting buckets in Latvia. In Proceedings of the 19th International Scientific Conference “Research for Rural Development 2013”, Jelgava, Latvia, 15–17 May 2013; pp. 59–66. [Google Scholar]
- Klavina, D.; Tadersoo, L.; Agan, A.; Adamson, K.; Bitinieks, K.; Gaitnieks, T.; Drenkhan, R. Soil fungal communities in young Norway-spruce dominant stands: Footprints of former agricultural land use and selective thinning. Eur. J. For. Res. 2022, 141, 503–516. [Google Scholar] [CrossRef]
- Tedersoo, L.; Lindahl, B. Fungal identification biases in microbiome projects. Environ. Microbiol. Rep. 2016, 8, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Anslan, S. Towards PacBio-based pan-eukaryote metabarcoding using full-length ITS sequences. Environ. Microbiol. Rep. 2019, 11, 659–668. [Google Scholar] [CrossRef]
- Anslan, S.; Bahram, M.; Hiiesalu, I.; Tedersoo, L. PipeCraft: Flexible open-source toolkit for bioinformatics analysis of custom high-throughput amplicon sequencing data. Mol. Ecol. Resour. 2017, 17, 234–240. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, J.B.; Clemente, C.J.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; Wit, P.D.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Engelbrecht Clemmensen, K.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 14 August 2022).
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, 11. [Google Scholar] [CrossRef]
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 14 August 2022).
- Ferlian, O.; Cesarz, S.; Craven, D.; Hines, J.; Barry, K.E.; Bruelheide, H.; Buscot, F.; Hayder, S.; Heklau, H.; Herrmann, S.; et al. Mycorrhiza in tree diversity–ecosystem function relationships: Conceptual framework and experimental implementation. Ecosphere 2018, 9, e02226. [Google Scholar] [CrossRef]
- Wagner, K.; Krause, K.; Gallegos-Monterrosa, R.; Sammer, D.; Kovacs, A.T.; Kothe, E. The Ectomycorrhizospheric Habitat of Norway Spruce and Tricholoma vaccinum: Promotion of Plant Growth and Fitness by a Rich Microorganismic Community. Front. Microbiol. 2019, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Marx, D.H.; Cordell, C.E.; Kenney, D.S.; Mexal, J.G.; Artman, J.D.; Riffle, J.W.; Molina, R. Commercial vegetative inoculum of Pisolithus tinctorius and inoculation techniques for development of ectomycorrhizae on bare root tree seedlings. For. Sci. Monogr. 1984, 25, 101. [Google Scholar]
- Perry, A.D.; Molina, R.; Amaranthus, P.M. Mycorrhizae, mycorrhizospheres, and reforestation: Current knowledge and research needs. Can. J. For. Res. 1987, 17, 929–940. [Google Scholar] [CrossRef]
- Ivory, M.; Munga, F. Growth and survival of container-grown Pinus caribaea infected with various ectomycorrhizal fungi. Plant Soil 1983, 71, 339–344. [Google Scholar] [CrossRef]
- Lee, K.J. A ten-year result of artificial inoculation of pines with ectomycorrhizal fungi, Pisolithus tinctorius and Thelephora terrestris. J. Korean For. Soc. 1992, 81, 156–163. [Google Scholar]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland 6: Russulacea (Lactarius and Russula); Verlag Mycologia: Lucern, Switzerland, 2005; 320p. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Taxon | Functional Group | Sequence Incidence among All Sequences, % (n = 105,208) | Incidence in All Samples, % (n = 60) | ||
|---|---|---|---|---|---|
| All | Stump Removal | Control | |||
| Thelephora terrestris | Ectomycorrhizal | 12.0 | 7.9 | 4.2 | 98.1 |
| Inocybe lacera | Ectomycorrhizal | 10.0 | 3.6 | 6.8 | 94.3 |
| Fungi sp.1 | Unknown | 3.0 | 1.8 | 1.3 | 66.0 |
| Fungi sp.2 | Unknown | 3.0 | 0.7 | 2.2 | 71.7 |
| Camposporium sp. | Saproxylic | 2.0 | 1.3 | 1.1 | 96.2 |
| Trichophaea sp. | Ectomycorrhizal | 2.0 | 1.3 | 0.9 | 75.5 |
| Trechisporales sp. | Unknown | 2.0 | 0.3 | 1.4 | 54.7 |
| Luellia recondite | Saproxylic | 2.0 | 0.5 | 1.0 | 32.1 |
| Hyaloscypha bicolor | Saprophytic | 2.0 | 0.6 | 1.0 | 92.5 |
| Lactarius sp. | Ectomycorrhizal | 2.0 | 1.0 | 0.5 | 26.4 |
| Hyaloscyphaceae sp. | Unknown | 2.0 | 0.4 | 1.1 | 75.5 |
| Amphinema sp. | Ectomycorrhizal | 1.0 | 0.4 | 0.9 | 56.6 |
| Mycena galopus | Saprophytic | 1.0 | 0.6 | 0.7 | 92.5 |
| Meliniomyces sp. | Endophytic | 1.0 | 0.4 | 0.7 | 77.4 |
| Alternaria rosae | Plant pathogen and saprophytic | 1.0 | 0.4 | 0.8 | 58.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burnevica, N.; Klavina, D.; Polmanis, K.; Jansons, J.; Gaitnieks, T. Impact of Stump Removal on Communities of Ectomycorrhizal and Other Soil Fungi in Norway Spruce Stands of Latvia. Diversity 2022, 14, 673. https://doi.org/10.3390/d14080673
Burnevica N, Klavina D, Polmanis K, Jansons J, Gaitnieks T. Impact of Stump Removal on Communities of Ectomycorrhizal and Other Soil Fungi in Norway Spruce Stands of Latvia. Diversity. 2022; 14(8):673. https://doi.org/10.3390/d14080673
Chicago/Turabian StyleBurnevica, Natalija, Darta Klavina, Kaspars Polmanis, Jurgis Jansons, and Talis Gaitnieks. 2022. "Impact of Stump Removal on Communities of Ectomycorrhizal and Other Soil Fungi in Norway Spruce Stands of Latvia" Diversity 14, no. 8: 673. https://doi.org/10.3390/d14080673
APA StyleBurnevica, N., Klavina, D., Polmanis, K., Jansons, J., & Gaitnieks, T. (2022). Impact of Stump Removal on Communities of Ectomycorrhizal and Other Soil Fungi in Norway Spruce Stands of Latvia. Diversity, 14(8), 673. https://doi.org/10.3390/d14080673